Dietary Antioxidant Trans-Cinnamaldehyde Reduced Visfatin-Induced Breast Cancer Progression: In Vivo and In Vitro Study



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Systemic Review
2.3. Meta-Analysis
2.4. Survival Rate
2.5. Cell Culture
2.6. 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) Cell Viability Assay
2.7. Cell Counting
2.8. Colony Formation
2.9. Western Blot Analysis
2.10. Nicotinamide phosphoribosyl transferase (NAMPT) Activity Kit
2.11. Xenograft Animal Model
2.12. Immunohistochemistry Analysis
2.13. Statistical Analysis
3. Results
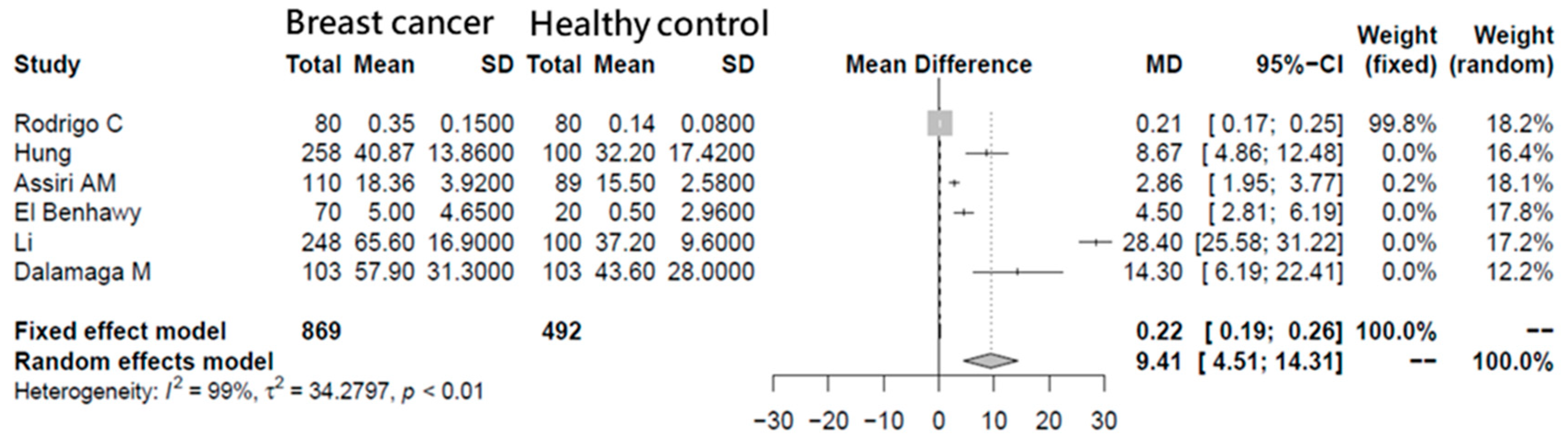
3.1. Meta-Analysis of Breast Cancer Patient Visfatin Concentrations
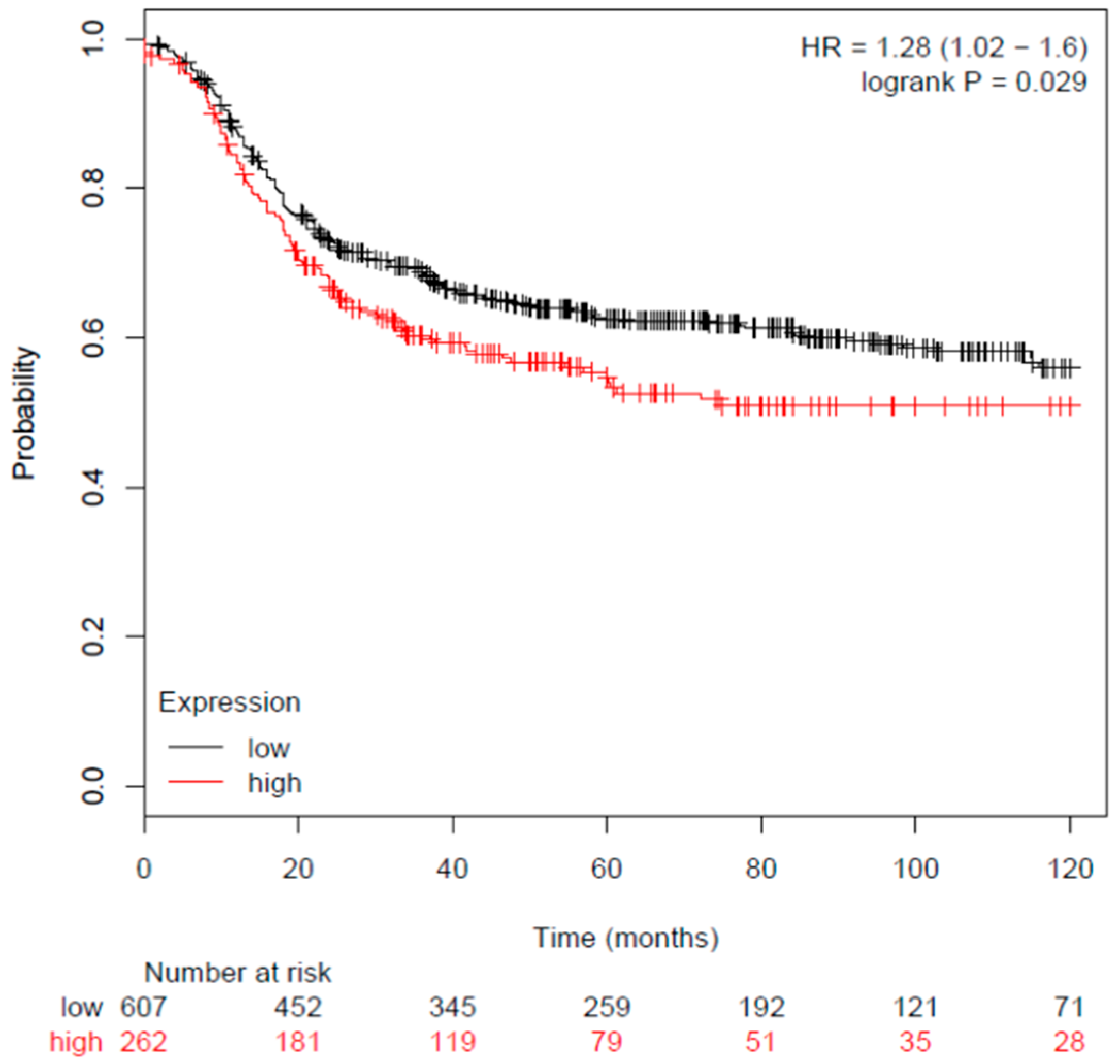
3.2. Breast Cancer Patient Visfatin Gene Expression and Survival Rate
3.3. Effects of cinnamaldehyde (CA) on Visfatin-Induced Breast Cancer Cell
3.3.1. Effect of Visfatin on Breast Cancer Cell Viability
3.3.2. Effect of CA on Breast Cancer Cell Viability
3.3.3. Anti-Proliferative Effect of cinnamaldehyde (CA) and FK866 Combined with Visfatin-Induced Breast Cancer Cell Viability
3.3.4. Anti-Colony Effect of CA Combined with Visfatin-Induced Breast Cancer Cells
3.4. Effects of CA and FK866 Combined with Visfatin on Proliferation-Related Protein Expression
3.5. Effects of CA and FK866 Combined with Visfatin on Intracellular and Extracellular Nicotinamide phosphoribosyl transferase (NAMPT) Protein Expression
3.6. Effect of CA on Visfatin-Induced Proliferation Xenograft Animal Model
3.6.1. Xenograft Animal Model Tumor Change
3.6.2. Xenograft Animal Model Protein Expression
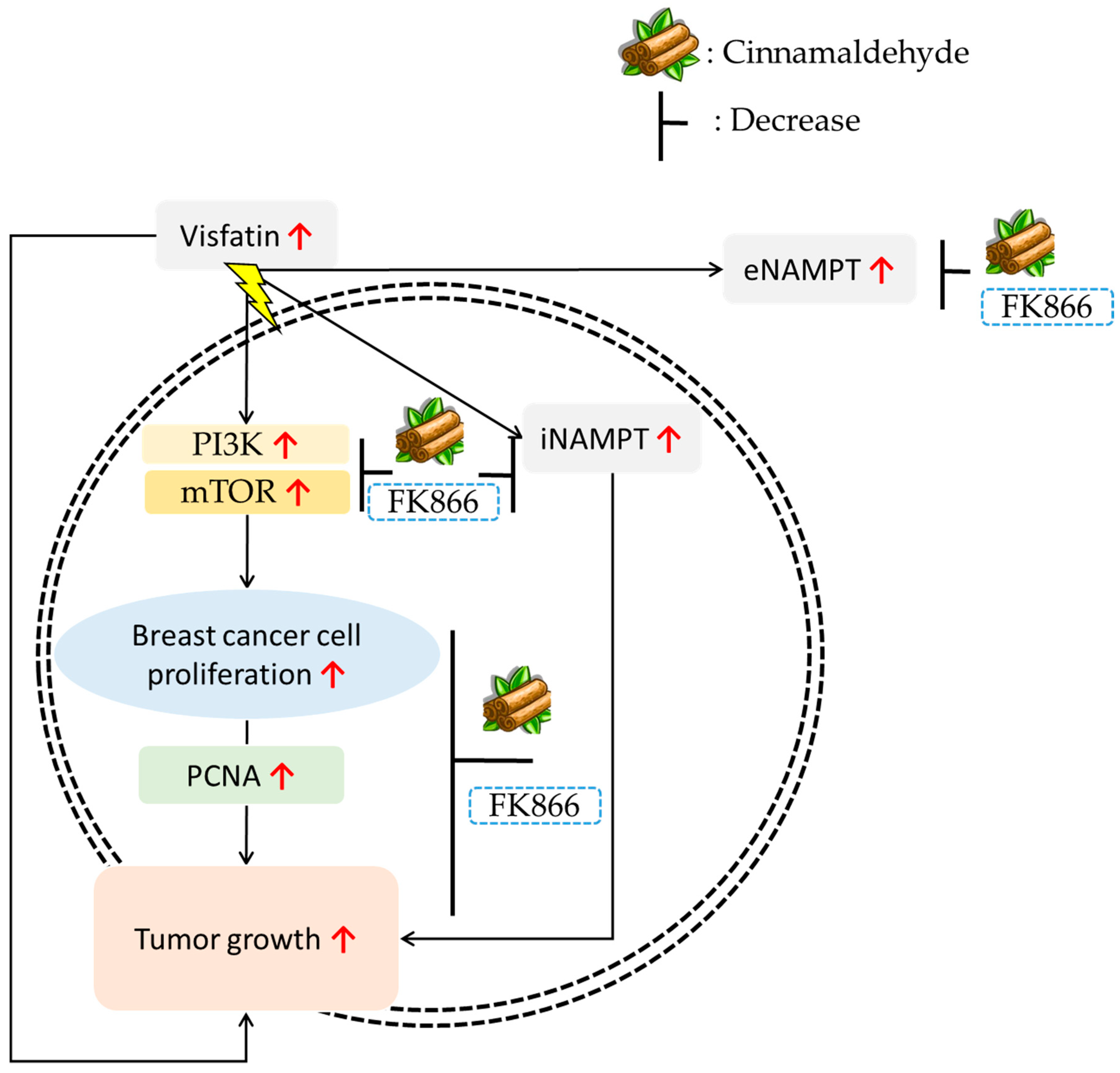
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Winters, S.; Martin, C.; Murphy, D.; Shokar, N.K. Breast Cancer Epidemiology, Prevention, and Screening. Prog. Mol. Biol. Transl. Sci. 2017, 151, 1–32. [Google Scholar] [PubMed]
- Guo, W.; Key, T.J.; Reeves, G.K. Adiposity and breast cancer risk in postmenopausal women: Results from the UK Biobank prospective cohort. Int. J. Cancer 2018, 143, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Renehan, A.G.; Tyson, M.; Egger, M.; Heller, R.F.; Zwahlen, M. Body-mass index and incidence of cancer: A systematic review and meta-analysis of prospective observational studies. Lancet 2008, 371, 569–578. [Google Scholar] [CrossRef]
- Wee, C.C.; Huang, A.; Huskey, K.W.; McCarthy, E.P. Obesity and the likelihood of sexual behavioral risk factors for HPV and cervical cancer. Obesity 2008, 16, 2552–2555. [Google Scholar] [CrossRef] [Green Version]
- Hung, A.C.; Lo, S.; Hou, M.-F.; Lee, Y.-C.; Tsai, C.-H.; Chen, Y.-Y.; Liu, W.; Su, Y.-H.; Lo, Y.-H.; Wang, C.-H.; et al. Extracellular Visfatin-Promoted Malignant Behavior in Breast Cancer Is Mediated Through c-Abl and STAT3 Activation. Clin. Cancer Res. 2016, 22, 4478–4490. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, X.; Liu, K.R.; Zhang, J.N.; Liu, Y.; Zhu, Y. Visfatin derived from ascites promotes ovarian cancer cell migration through Rho/ROCK signaling-mediated actin polymerization. Eur. J. Cancer Prev. 2015, 24, 231–239. [Google Scholar] [CrossRef]
- Carbone, F.; Liberale, L.; Bonaventura, A.; Vecchie, A.; Casula, M.; Cea, M.; Monacelli, F.; Caffa, I.; Bruzzone, S.; Montecucco, F.; et al. Regulation and Function of Extracellular Nicotinamide Phosphoribosyltransferase/Visfatin. Compr. Physiol. 2017, 7, 603–621. [Google Scholar]
- Hug, C.; Lodish, H.F. Medicine. Visfatin: A new adipokine. Science 2005, 307, 366–367. [Google Scholar] [CrossRef]
- Sheikhpour, R. Visfatin and its role in breast cancer. Middle East J. Cancer 2017, 8, 171–177. [Google Scholar]
- Mohammadi, M.; Mianabadi, F.; Mehrad-Majd, H. Circulating visfatin levels and cancers risk: A systematic review and meta-analysis. J. Cell. Physiol. 2019, 234, 5011–5022. [Google Scholar] [CrossRef]
- Lin, Y.T.; Chen, L.K.; Jian, D.Y.; Hsu, T.C.; Huang, W.C.; Kuan, T.T.; Wu, S.Y.; Kwok, C.F.; Ho, L.T.; Juan, C.C. Visfatin Promotes Monocyte Adhesion by Upregulating ICAM-1 and VCAM-1 Expression in Endothelial Cells via Activation of p38-PI3K-Akt Signaling and Subsequent ROS Production and IKK/NF-kappaB Activation. Cell. Physiol. Biochem. 2019, 52, 1398–1411. [Google Scholar] [PubMed]
- Gholinejad, Z.; Kheiripour, N.; Nourbakhsh, M.; Ilbeigi, D.; Behroozfar, K.; Hesari, Z.; Golestani, A.; Shabani, M.; Einollahi, N. Extracellular NAMPT/Visfatin induces proliferation through ERK1/2 and AKT and inhibits apoptosis in breast cancer cells. Peptides 2017, 92, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, B.E.; Sharif, T.; Martell, E.; Dai, C.; Kim, Y.; Lee, P.W.K.; Gujar, S.A. NAD+ salvage pathway in cancer metabolism and therapy. Pharmacol. Res. 2016, 114, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Kulikova, V.; Shabalin, K.; Nerinovski, K.; Dolle, C.; Niere, M.; Yakimov, A.; Redpath, P.; Khodorkovskiy, M.; Migaud, M.E.; Ziegler, M.; et al. Generation, Release, and Uptake of the NAD Precursor Nicotinic Acid Riboside by Human Cells. J. Biol. Chem. 2015, 290, 27124–27137. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.H.; Ismail, I.A.; Kang, S.M.; Han, D.C.; Kwon, B.M. Cinnamaldehydes in Cancer Chemotherapy. Phytother. Res. 2016, 30, 754–767. [Google Scholar] [CrossRef]
- Zhao, H.; Xie, Y.; Yang, Q.; Cao, Y.; Tu, H.; Cao, W.; Wang, S. Pharmacokinetic study of cinnamaldehyde in rats by GC–MS after oral and intravenous administration. J. Pharm. Biomed. Anal. 2014, 89, 150–157. [Google Scholar] [CrossRef]
- Paul, M.B. Cinnamaldehyde—Molecule of the Month August 2006 [Archived Version]; Hillsborough College: Sheffield, UK, 2017. [Google Scholar]
- Koppikar, S.J.; Choudhari, A.S.; Suryavanshi, S.A.; Kumari, S.; Chattopadhyay, S.; Kaul-Ghanekar, R. Aqueous cinnamon extract (ACE-c) from the bark of Cinnamomum cassia causes apoptosis in human cervical cancer cell line (SiHa) through loss of mitochondrial membrane potential. BMC Cancer 2010, 10, 210. [Google Scholar] [CrossRef] [Green Version]
- Tian, F.; Yu, C.T.; Ye, W.D.; Wang, Q. Cinnamaldehyde induces cell apoptosis mediated by a novel circular RNA hsa_circ_0043256 in non-small cell lung cancer. Biochem. Biophys. Res. Commun. 2017, 493, 1260–1266. [Google Scholar] [CrossRef]
- Wu, C.; Zhuang, Y.; Jiang, S.; Tian, F.; Teng, Y.; Chen, X.; Zheng, P.; Liu, S.; Zhou, J.; Wu, J.; et al. Cinnamaldehyde induces apoptosis and reverses epithelial-mesenchymal transition through inhibition of Wnt/beta-catenin pathway in non-small cell lung cancer. Int. J. Biochem. Cell Biol. 2017, 84, 58–74. [Google Scholar] [CrossRef]
- Zheng, Y.; Geng, S.; Meng, M.; Du, Z.; Yao, J.; Li, Z.; Zhang, Q.; Zhang, Z.; Duan, Y.; Du, G. The Edible Adlay Seed Oil Containing Cinnamaldehyde and Gingerol Prevents DMBA/TPA-Induced Skin Carcinogenesis by Cooperative Ablation of Lipid Rafts. J. Arcinogene. Mutagene 2017, 8, 302. [Google Scholar] [CrossRef]
- Aggarwal, B.B. Targeting inflammation-induced obesity and metabolic diseases by curcumin and other nutraceuticals. Annu. Rev. Nutr. 2010, 30, 173–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assiri, A.M.; Kamel, H.F. Evaluation of diagnostic and predictive value of serum adipokines: Leptin, resistin and visfatin in postmenopausal breast cancer. Obes. Res. Clin. Pract. 2016, 10, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Tang, S.H.; Zhou, X.C.; Ye, Y.H.; Xu, X.Q.; Li, R.Z. Preoperative serum visfatin levels and prognosis of breast cancer among Chinese women. Peptides 2014, 51, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Dalamaga, M. Nicotinamide phosphoribosyl-transferase/visfatin: A missing link between overweight/obesity and postmenopausal breast cancer? Potential preventive and therapeutic perspectives and challenges. Med. Hypotheses 2012, 79, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, C.; Tennekoon, K.H.; Karunanayake, E.H.; De Silva, K.; Amarasinghe, I.; Wijayasiri, A. Circulating leptin, soluble leptin receptor, free leptin index, visfatin and selected leptin and leptin receptor gene polymorphisms in sporadic breast cancer. Endocr. J. 2017, 64, 393–401. [Google Scholar] [CrossRef] [Green Version]
- El-Benhawy, S.A.; Abd El Moneim, N.A.; Ebeid, S.A. Serum Adipocytokines (Visfatin and Resistin): New Biomarkers of Breast Carcinogenesis. Middle East J. Cancer 2015, 6, 253–265. [Google Scholar]
- Gyorffy, B.; Lanczky, A.; Eklund, A.C.; Denkert, C.; Budczies, J.; Li, Q.; Szallasi, Z. An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1,809 patients. Breast Cancer Res. Treat. 2010, 123, 725–731. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, A.N.; Ahmad, R.; Khan, M.A. Evaluation and Comparison of the In Vitro Cytotoxic Activity of Withania somnifera Methanolic and Ethanolic Extracts against MDA-MB-231-GFPand Vero Cell Lines. Sci. Pharm. 2015, 84, 41–59. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.-H.; Chu, S.-C.; Yang, S.-F.; Hsieh, Y.-S.; Lee, C.-Y.; Chen, P.-N. Induction of apoptotic but not autophagic cell death by Cinnamomum cassia extracts on human oral cancer cells. J. Cell. Physiol. 2019, 234, 5289–5303. [Google Scholar] [CrossRef]
- Chen, H.-Y.; Huang, T.-C.; Shieh, T.-M.; Wu, C.-H.; Lin, L.-C.; Hsia, S.-M. Isoliquiritigenin Induces Autophagy and Inhibits Ovarian Cancer Cell Growth. Int. J. Mol. Sci. 2017, 18, 2025. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gao, C.; Zhang, Y.; Gao, J.; Teng, F.; Tian, W.; Yang, W.; Yan, Y.; Xue, F. Visfatin stimulates endometrial cancer cell proliferation via activation of PI3K/Akt and MAPK/ERK1/2 signalling pathways. Gynecol. Oncol. 2016, 143, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Gowder, S.J.B. Safety assessment of food flavor-cinnamaldehyde. Biosafety 2014, 3, e147. [Google Scholar] [CrossRef]
- Wani, K.; Kadu, B.; Mansara, P.; Gupta, P.; Deore, A.; Chikate, R.; Poddar, P.; Dhole, S.; Kaul-Ghanekar, R. Synthesis, Characterization and In Vitro Study of Biocompatible Cinnamaldehyde Functionalized Magnetite Nanoparticles (CPGF Nps) For Hyperthermia and Drug Delivery Applications in Breast Cancer. PLoS ONE 2014, 9, e107315. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, R.; Liu, H.; Liu, C.; Wang, L.; Ma, R.; Chen, B.; Li, L.; Niu, J.; Fu, M.; Zhang, D.; et al. Cinnamaldehyde in diabetes: A review of pharmacology, pharmacokinetics and safety. Pharmacol. Res. 2017, 122, 78–89. [Google Scholar] [CrossRef]
- Verdin, E. NAD+ in aging, metabolism, and neurodegeneration. Science 2015, 350, 1208–1213. [Google Scholar] [CrossRef]
- Bi, T.Q.; Che, X.M. Nampt/PBEF/visfatin and cancer. Cancer Biol. Ther. 2010, 10, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.-C.; Song, J.; Lee, A.; Cho, D.; Kim, T.S. Visfatin Promotes Wound Healing through the Activation of ERK1/2 and JNK1/2 Pathway. Int. J. Mol. Sci. 2018, 19, 3642. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.R.; Park, H.J.; Bae, Y.H.; Ahn, S.C.; Wee, H.J.; Yun, I.; Jang, H.O.; Bae, M.K.; Bae, S.K. Curcumin down-regulates visfatin expression and inhibits breast cancer cell invasion. Endocrinology 2012, 153, 554–563. [Google Scholar] [CrossRef] [Green Version]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiang, Y.-F.; Chen, H.-Y.; Huang, K.-C.; Lin, P.-H.; Hsia, S.-M. Dietary Antioxidant Trans-Cinnamaldehyde Reduced Visfatin-Induced Breast Cancer Progression: In Vivo and In Vitro Study. Antioxidants 2019, 8, 625. https://doi.org/10.3390/antiox8120625
Chiang Y-F, Chen H-Y, Huang K-C, Lin P-H, Hsia S-M. Dietary Antioxidant Trans-Cinnamaldehyde Reduced Visfatin-Induced Breast Cancer Progression: In Vivo and In Vitro Study. Antioxidants. 2019; 8(12):625. https://doi.org/10.3390/antiox8120625
Chicago/Turabian StyleChiang, Yi-Fen, Hsin-Yuan Chen, Ko-Chieh Huang, Po-Han Lin, and Shih-Min Hsia. 2019. "Dietary Antioxidant Trans-Cinnamaldehyde Reduced Visfatin-Induced Breast Cancer Progression: In Vivo and In Vitro Study" Antioxidants 8, no. 12: 625. https://doi.org/10.3390/antiox8120625