How Does Pyridoxamine Inhibit the Formation of Advanced Glycation End Products? The Role of Its Primary Antioxidant Activity

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
3. Results
- Hydrogen-atom transfer (HAT): PM + R•→ PM(-H)• + RH, considering each possible PM hydrogen atom.
- Radical-adduct formation (RAF): PM + R•→ [PM-R]•, at each aromatic PM atom.
- Single-electron transfer (SET): PM + R•→ PM+• + R.
3.1. Thermodynamic Study
3.2. Kinetic Study
3.3. Comparison to Other Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AGE | Advanced glycation end product |
| ROS | Reactive oxygen species |
| RCS | Reactive carbonyl species |
| NMR | Nuclear magnetic resonance |
| DFT | Density functional theory |
| PM | Pyridoxamine |
| PL | Pyridoxal |
| PN | Pyridoxine |
| PLP | Pyridoxal 5’-phosphate |
| QM-ORSA | Quantum mechanics-based test for overall free radical scavenging activity |
| TS | Transition state |
| SMD | Solvation Model based on the quantum mechanical charge Density of a solute molecule interacting with a continuum |
| HAT | Hydrogen atom transfer |
| RAF | Radical adduct formation |
| SET | Single electron transfer |
| TST | Transition state theory |
References
- Bohlender, J.M.; Franke, S.; Stein, G.; Wolf, G. Advanced Glycation End Products and the Kidney. Am. J. Physiol. Ren. Physiol. 2005, 289, 645–659. [Google Scholar] [CrossRef] [PubMed]
- Stitt, A.W. The Maillard Reaction in Eye Diseases. Ann. N. Y. Acad. Sci. 2005, 1043, 582–597. [Google Scholar] [CrossRef] [PubMed]
- Kume, S.; Takeya, M.; Mori, T.; Araki, N.; Suzuki, H.; Horiuchi, S.; Kodama, T.; Miyauchi, Y.; Takahashi, K. Immunohistochemical and Ultrastructural Detection of Advanced Glycation End Products in Atherosclerotic Lesions of Human Aorta with a Novel Specific Monoclonal Antibody. Am. J. Pathol. 1995, 147, 654–667. [Google Scholar] [PubMed]
- Thorpe, S.R.; Baynes, J.W. Maillard Reaction Products in Tissue Proteins: New Products and New Perspectives. Amino Acids 2003, 25, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, M. Advanced Protein Glycosylation in Diabetes and Aging. Annu. Rev. Med. 1995, 46, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, P.; Cerami, A. Protein Glycation, Diabetes, and Aging. Recent. Prog. Horm. Res. 2001, 56, 1–21. [Google Scholar] [CrossRef]
- Reynolds, T.M. Chemistry of Nonenzymatic Browning II. Adv. Food Res. 1965, 14, 167–283. [Google Scholar]
- Rahbar, S.; Figarola, J.L. Inhibitors and Breakers of Advanced Glycation Endproducts (AGEs): A Review. Curr. Med. Chem. Immun. Enodcrinol. Metab. Agents 2002, 2, 135–161. [Google Scholar] [CrossRef]
- Monnier, V.M. Intervention Against the Maillard Reaction in Vivo. Arch. Biochem. Biophys. 2003, 419, 1–15. [Google Scholar] [CrossRef]
- Khalifah, R.G.; Baynes, J.W.; Hudson, B.G. Amadorins: Novel Post-Amadori Inhibitors of Advanced Glycation Reactions. Biochem. Biophys. Res. Commun. 1999, 257, 251–258. [Google Scholar] [CrossRef]
- Snell, E.E. The Vitamin B6 Group. V. The Reversible Interconversion of Pyridoxal and Pyridoxamine by Transamination Reactions. J. Am. Chem. Soc. 1945, 67, 194–197. [Google Scholar] [CrossRef]
- Degenhardt, T.P.; Alderson, N.L.; Arrington, D.D.; Beattie, R.J.; Basgen, J.M.; Steffes, M.W.; Thorpe, S.R.; Baynes, J.W. Pyridoxamine Inhibits Early Renal Disease and Dyslipidemia in the Streptozotocin-Diabetic Rat. Kidney Int. 2002, 61, 939–950. [Google Scholar] [CrossRef] [PubMed]
- Murakoshi, M.; Tanimoto, M.; Gohda, T.; Hagiwara, S.; Ohara, I.; Toyoda, H.; Ishikawa, Y.; Horikoshi, S.; Tomino, Y. Pleiotropic Effect of Pyridoxamine on Diabetic Complications Via CD36 Expression in KK-Ay/Ta Mice. Diabetes Res. Clin. Pract. 2009, 83, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Stitt, A.; Gardiner, T.A.; Alderson, N.L.; Canning, P.; Frizzell, N.; Duffy, N.; Boyle, C.; Januszewski, A.S.; Chachich, M.; Baynes, J.W.; et al. The AGE Inhibitor Pyridoxamine Inhibits Development of Retinopathy in Experimental Diabetes. Diabetes 2002, 51, 2826–2832. [Google Scholar] [CrossRef] [PubMed]
- Wu, E.; Liang, J.; Wu, M.; Chang, K. Pyridoxamine Prevents Age-Related Aortic Stiffening and Vascular Resistance in Association with Reduced Collagen Glycation. Exp. Gerontol. 2011, 46, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Adrover, M.; Vilanova, B.; Frau, J.; Muñoz, F.; Donoso, J. The Pyridoxamine Action on Amadori Compounds: A Reexamination of its Scavenging Capacity and Chelating Effect. Bioorg. Med. Chem. 2008, 16, 5557–5569. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.M.; Balenovich, W.; Hornich, L.H.M.; Richardson, M.R. Reactions of Metal Ions With Vitamins. IV. The Crystal Structure of a Zinc Complex of Pyridoxamine (Vitamin B6). Inorg. Chim. Acta 1980, 46, 199–203. [Google Scholar] [CrossRef]
- Ortega-Castro, J.; Adrover, M.; Frau, J.; Salvà, A.; Donoso, J.; Muñoz, F. DFT Studies on Schiff Base Formation of Vitamin B6 Analogues. Reaction Between a Pyridoxamine-Analogue and Carbonyl Compounds. J. Phys. Chem. A 2010, 114, 4634–4640. [Google Scholar] [CrossRef] [PubMed]
- Adrover, M.; Vilanova, B.; Muñoz, F.; Donoso, J. A Comparative Study of the Chemical Reactivity of Pyridoxamine, Ac-Phe-Lys and Ac-Cys with Various Glycating Carbonyl Compounds. Amino Acids 2009, 36, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Voziyan, P.A.; Metz, T.O.; Baynes, J.W.; Hudson, B.G. A Post-Amadori Inhibitor Pyridoxamine Also Inhibits Chemical Modification of Proteins by Scavenging Carbonyl Intermediates of Carbohydrate and Lipid Degradation. J. Biol. Chem. 2002, 277, 3397–3403. [Google Scholar] [CrossRef]
- Chetykrin, S.; Zhang, W.; Hudson, B.D.; Serianni, A.S.; Voziyan, P.V. Pyridoxamine Protects Protein from Functional Damage by 3-Deoxyglucosone: Mechanism of Action for Pyridoxamine. Biochemistry 2008, 47, 997–1006. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.C. Hydroxyl Radicals Assayed by Aromatic Hydroxylation and Deoxyribose Degradation. In Handbook of Methods for Oxygen Radical Research; Greenwald, R.A., Ed.; CRC Press: Boca Raton, FL, USA, 1986; pp. 177–180. [Google Scholar]
- Voziyan, P.A.; Hudson, B.G. Pyridoxamine: The Many Virtues of a Maillard Reaction Inhibitor. Ann. N. Y. Acad. Sci. 2005, 1043, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Chetykrin, S.; Mathis, M.E.; Ham, A.J.L.; Hachey, D.L.; Hudson, B.G.; Voziyan, P.V. Propagation of Protein Glycation Damage Involves Modification of Tryptophan Residues Via Reactive Oxygen Species: Inhibition by Pyridoxamine. Free Radic. Biol. Med. 2008, 44, 1276–1285. [Google Scholar]
- Jain, S.K.; Lim, G. Pyridoxine and Pyridoxamine Inhibits Superoxide Radicals and Prevents Lipid Peroxidation, Protein Glycosylation, and (Na++K+)-ATPase Activity Reduction in High Glucose-Treated Human Erythrocytes. Free Rad. Biol. Med. 2001, 30, 232–237. [Google Scholar] [CrossRef]
- Cao, G.; Alessio, H.M.; Cutler, R.G. Oxygen-Radical Absorbance Capacity Assay for Antioxidants. Free Radic. Biol. Med. 1993, 14, 303–311. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant Determinations by the Use of a Stable Free Radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Matxain, J.; Ristila, M.; Strid, A.; Eriksson, L.A. Theoretical Study of the Antioxidant Properties of Pyridoxine. J. Phys. Chem. A 2006, 110, 13068–13072. [Google Scholar] [CrossRef]
- Matxain, J.; Ristila, M.; Strid, A.; Eriksson, L.A. Evidence of High •OH Radical Quenching Efficiency by Vitamin B6. J. Phys. Chem. B 2009, 113, 9629–9632. [Google Scholar] [CrossRef]
- Galano, A.; Alvarez-Idaboy, J.R.; Francisco-Márquez, M.; Medina, M.E. A Quantum Chemical Study on the Free Radical Scavenging Activity of Tyrosol and Hydroxytyrosol. Theor. Chem. Acc. 2012, 131, 1–12. [Google Scholar] [CrossRef]
- León-Carmona, J.; Galano, A. Is Caffeine a Good Scavenger of Oxygenated Free Radicals? J. Phys. Chem. B 2011, 115, 4538–4546. [Google Scholar] [CrossRef]
- Galano, A.; Alvarez-Idaboy, J.R. Glutathione: Mechanism and Kinetics of Its Non-Enzymatic Defense Action Against Free Radicals. RSC Adv. 2011, 1, 1763–1771. [Google Scholar] [CrossRef]
- Galano, A.; Francisco-Márquez, M.; Alvarez-Idaboy, J.R. Canolol: A Promising Chemical Agent Against Oxidative Stress. J. Phys. Chem. B 2011, 115, 8590–8596. [Google Scholar] [CrossRef] [PubMed]
- Galano, A. On the Direct Scavenging Activity of Melatonin Towards Hydroxyl and a Series of Peroxyl Radicals. Phys. Chem. Chem. Phys. 2011, 13, 7147–7157. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Castro, J.; Adrover, M.; Frau, J.; Donoso, J.; Muñoz, F. Cu2+ Complexes of Some AGEs Inhibitors. Chem. Phys. Lett. 2009, 475, 277–284. [Google Scholar] [CrossRef]
- Ortega-Castro, J.; Frau, J.; Casasnovas, R.; Fernández, D.; Donoso, J.; Muñoz, F. High- and Low-Spin Fe(III) Complexes of Various AGE Inhibitors. J. Phys. Chem. A 2012, 116, 2961–2971. [Google Scholar] [CrossRef] [PubMed]
- Casasnovas, R.; Ortega-Castro, J.; Donoso, J.; Frau, J.; Muñoz, F. Theoretical Calculations of Stability Constants and pKa Values of Metal Complexes in Solution: Application to Pyridoxamine-Copper(II) Complexes and Their Biological Implications in AGE Inhibition. Phys. Chem. Chem. Phys. 2013, 15, 16303–16313. [Google Scholar] [CrossRef]
- Adrover, M.; Vilanova, B.; Muñoz, F.; Donoso, J. Pyridoxamine, A Scavenger Agent of Carbohydrates. Int. J. Chem. Kinet. 2007, 39, 154–167. [Google Scholar] [CrossRef]
- Adrover, M.; Vilanova, B.; Muñoz, F.; Donoso, J. Inhibition of Glycosylation Processes: The Reaction Between Pyridoxamine and Glucose. Chem. Biodivers. 2005, 2, 964–975. [Google Scholar] [CrossRef]
- Rose, R.C.; Bode, A.M. Biology of Free Radical Scavengers. FASEB J. 1993, 7, 1135–1142. [Google Scholar] [CrossRef]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin as a Naturall Ally Against Oxidative Stress: A Physicochemical Examination. J. Pineal Res. 2011, 51, 1–16. [Google Scholar] [CrossRef]
- Galano, A.; Alvarez-Idaboy, J.R. A Computational Methodology for Accurate Prediction of Rate Constants in Solution: Application to the Assessment of Primary Antioxidant Activity. J. Comput. Chem. 2013, 34, 2430–2445. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Schultz, N.E.; Truhlar, D.G. Design of Density Functionals by Combining the Method of Constraint Satisfaction with Parametrization for Thermochemistry, Thermochemical Kinetics, and Noncovalent Interactions. J. Chem. Theory Comput. 2006, 2, 364–382. [Google Scholar] [CrossRef] [PubMed]
- Black, G.; Simmie, J.M. Barrier Heights for H-Atom Abstraction by HO2• from n-Butanol—A Simple Yet Exacting Test for Model Chemistries? J. Comput. Chem. 2009, 31, 1236–1248. [Google Scholar]
- Furuncuoǧlu, T.; Uǧur, I.; Deǧirmenci, I.; Aviyente, V. Role of Chain Transfer Agents in Free Radical Polymerization Kinetics. Macromolecules 2010, 43, 1823–1835. [Google Scholar] [CrossRef]
- Zhao, Y.; Truhlar, D.G. How Well Can New-Generation Density Functionals Describe the Energetics of Bond-Dissociation Reactions Producing Radicals? J. Phys. Chem. A 2008, 112, 1095–1099. [Google Scholar] [CrossRef] [PubMed]
- Marenich, A.V.; Cramer, C.J.; Truhlar, D.G. Universal Solvation Model Based on Solute Electron Density and on a Continuum Model of the Solvent Defined by the Bulk Dielectric Constant and Atomic Surface Tensions. J. Phys. Chem. B 2009, 113, 6378–6396. [Google Scholar] [CrossRef] [PubMed]
- Vilanova, B.; Adrover, M.; Muñoz, F.; Donoso, J. Photo-Induced Processes in Vitamin B6 Compounds. Chem. Biodivers. 2004, 1, 1073–1090. [Google Scholar] [CrossRef] [PubMed]
- Okuno, Y. Theoretical Investigation of the Mechanism of the Baeyer-Villiger Reaction in Nonpolar Solvents. Chem. Eur. J. 1997, 3, 212–218. [Google Scholar] [CrossRef]
- Benson, S.W. The Foundations of Chemical Kinetics; Krieger: Malabar, FL, USA, 1982. [Google Scholar]
- Eckart, C. The Penetration of a Potential Barrier by Electrons. Phys. Rev. 1930, 35, 1303–1309. [Google Scholar] [CrossRef]
- Brown, R.L. A Method of Calculating Tunneling Corrections for Eckart Potential Barriers. J. Res. Natl. Bur. Stand. 1981, 86, 357–359. [Google Scholar] [CrossRef]
- Collins, F.C.; Kimball, G.E. Diffusion-Controlled Reaction Rates. J. Colloid Sci. 1949, 4, 425–437. [Google Scholar] [CrossRef]
- Smoluchowski, M.Z. Attempt for a Mathematical Theory of Kinetic Coagulation of Colloid Solutions. J. Phys. Chem. 1917, 92, 129–168. [Google Scholar]
- Stokes, G.G. Mathematical and Physical Papers; Cambridge University Press: Cambridge, UK, 1903; Volume 3, p. 55. [Google Scholar]
- Einstein, A. Motion of Suspended Particles in Stationary Liquids Required from the Molecular Kinetic Theory of Heat. Ann. Phys. 1905, 17, 549–560. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A.; et al. Gaussian09, Revision D.01; Gaussian, Inc.: Wallingford, CT, USA, 2013. [Google Scholar]
- Galano, A.; Martínez, A. Capsaicin, a Tasty Free Radical Scavenger: Mechanism of Action and Kinetics. J. Phys. Chem. B 2012, 116, 1200–1208. [Google Scholar] [CrossRef] [PubMed]
- Amić, A.; Marković, Z.; Klein, E.; Dimitrić Marković, J.M.; Milenković, D. Theoretical Study of the Thermodynamics of the Mechanisms Underlying Antiradical Activity of Cinnamic Acid Derivatives. Food Chem. 2018, 246, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Francisco-Marquez, M. Reactions of OOH Radical with β-Carotene, Lycopene, and Torulene: Hydrogen Atom Transfer and Adduct Formation Mechanisms. J. Phys. Chem. B 2009, 113, 11338–11345. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Francisco-Marquez, M. Peroxyl-Radical-Scavenging Activity of Garlic: 2-Propenesulfenic Acid versus Allicin. J. Phys. Chem. B 2009, 113, 16077–16081. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.W.; Doba, T.; Gabe, E.; Hughes, L.; Lee, F.L.; Prasad, L.; Ingold, K.U. Autoxidation of Biological Molecules. 4. Maximizing the Antioxidant Activity of Phenols. J. Am. Chem. Soc. 1985, 107, 7053–7065. [Google Scholar] [CrossRef]
- Galano, A.; León-Carmona, J.R.; Alvarez-Idaboy, J.R. Influence of the Environment on the Protective Effects of Guaiacol Derivatives against Oxidative Stress: Mechanisms, Kinetics, and Relative Antioxidant Activity. J. Phys. Chem. B 2012, 116, 7129–7137. [Google Scholar] [CrossRef]
- Galano, A.; Alvarez-Idaboy, J.R.; Francisco-Márquez, M. Physicochemical Insights on the Free Radical Scavenging Activity of Sesamol: Importance of the Acid/Base Equilibrium. J. Phys. Chem. B 2011, 115, 13101–13109. [Google Scholar] [CrossRef]
- Booth, A.A.; Khalifah, R.G.; Hudson, B.G. Thiamine Pyrophosphate and Pyridoxamine Inhibit the Formation of Antigenic Advanced Glycation End-Products: Comparison with Aminoguanidine. Biochem. Biophys. Res. Commun. 1996, 220, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Stocker, P.; Lesgards, J.F.; Vidal, N.; Chalier, F.; Prost, M. ESR Study of a Biological Assay on Whole Blood: Antioxidant Efficiency of Various Vitamins. Biochim. Biophys. Acta Gen. Subj. 2003, 1621, 1–8. [Google Scholar] [CrossRef]
- Merigliano, C.; Mascolo, E.; Burla, R.; Saggio, I.; Verni, F. The Relationship Between Vitamin B6, Diabetes and Cancer. Front. Genet. 2018, 9, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Voziyan, P.A.; Khalifah, R.G.; Thibaudeau, C.; Yildiz, A.; Jacob, J.; Serianni, A.S.; Hudson, B.G. Modification of Proteins In Vitro by Physiological Levels of Glucose. J. Biol. Chem. 2003, 278, 46616–46624. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, R.H.; Sarkar, P.; Mally, A.; Biemel, K.M.; Lederer, M.O.; Padayatti, P.S. Effect of Pyridoxamine on Chemical Modification of Proteins by Carbonyls in Diabetic Rats: Characterization of a Major Product from the Reaction of Pyridoxamine and Methylglyoxal. Arch. Biochem. Biophys. 2002, 402, 110–119. [Google Scholar] [CrossRef]
- Amarnath, V.; Amarnath, K.; Amarnath, K.; Davies, S.; Roberts, L.J. Pyridoxamine: An Extremely Potent Scavenger of 1,4-Dicarbonyls. Chem. Res. Toxicol. 2004, 17, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Ehrenshaft, M.; Jenns, A.E.; Chung, K.R.; Daub, M.E. SOR1, a Gene Required for Photosensitizer and Singlet Oxygen Resistance in Cercospora Fungi, Is Highly Conserved in Divergent Organisms. Mol. Cell 1998, 1, 603–609. [Google Scholar] [CrossRef]
- Bilski, P.; Li, M.Y.; Ehrenshaft, M.; Daub, M.E.; Chignell, C.F. Vitamin B6 (Pyridoxine) and Its Derivatives Are Efficient Singlet Oxygen Quenchers and Potential Fungal Antioxidants. Photochem. Photobiol. 2000, 71, 129–134. [Google Scholar] [CrossRef]
- Denslow, S.A.; Walls, A.A.; Daub, M.E. Regulation of Biosynthetic Genes and Antioxidant Properties of Vitamin B6 Vitamers During Plant Defense Responses. Physiol. Mol. Plant Pathol. 2005, 66, 244–255. [Google Scholar] [CrossRef]
- Danon, A.; Miersch, O.; Felix, G.; Camp, R.G.L.; Apel, K. Concurrent Activation of Cell Death-Regulating Signaling Pathways by Singlet Oxygen in Arabidopsis Thaliana. Plant J. 2005, 41, 68–80. [Google Scholar] [CrossRef]
- Anand, S.S. Protective Effect of Vitamin B6 in Chromium-Induced Oxidative Stress in Liver. J. Appl. Toxicol. 2005, 25, 440–443. [Google Scholar] [CrossRef] [PubMed]
- Gliszczyńska-Świglo, A. Antioxidant Activity of Water Soluble Vitamins in the TEAC (Trolox Equivalent Antioxidant Capacity) and the FRAP (Ferric Reducing Antioxidant Power) Assays. Food Chem. 2006, 96, 131–136. [Google Scholar] [CrossRef]
- Matxain, J.M.; Ristilä, M.; Strid, A.; Eriksson, L.A. Theoretical Study of the Reaction of Vitamin B6 with 1O2. Chem. Eur. J. 2007, 13, 4636–4642. [Google Scholar] [CrossRef] [PubMed]
- Škorňa, P.; Rimarčíc, J.; Poliak, P.; Lukeš, V.; Klein, E. Thermodynamic Study of Vitamin B6 Antioxidant Potential. Comput. Theor. Chem. 2016, 1077, 32–38. [Google Scholar] [CrossRef]
- Abdullah, K.M.; Qais, F.A.; Ahmad, I.; Hasan, H.; Naseem, I. Study of Pyridoxamine Against Glycation and Reactive Oxygen Species Production in Human Serum Albumin as Model Protein: An In Vitro & Ex Vivo Approach. Int. J. Biol. Macromol. 2018, 120, 1734–1743. [Google Scholar] [PubMed]



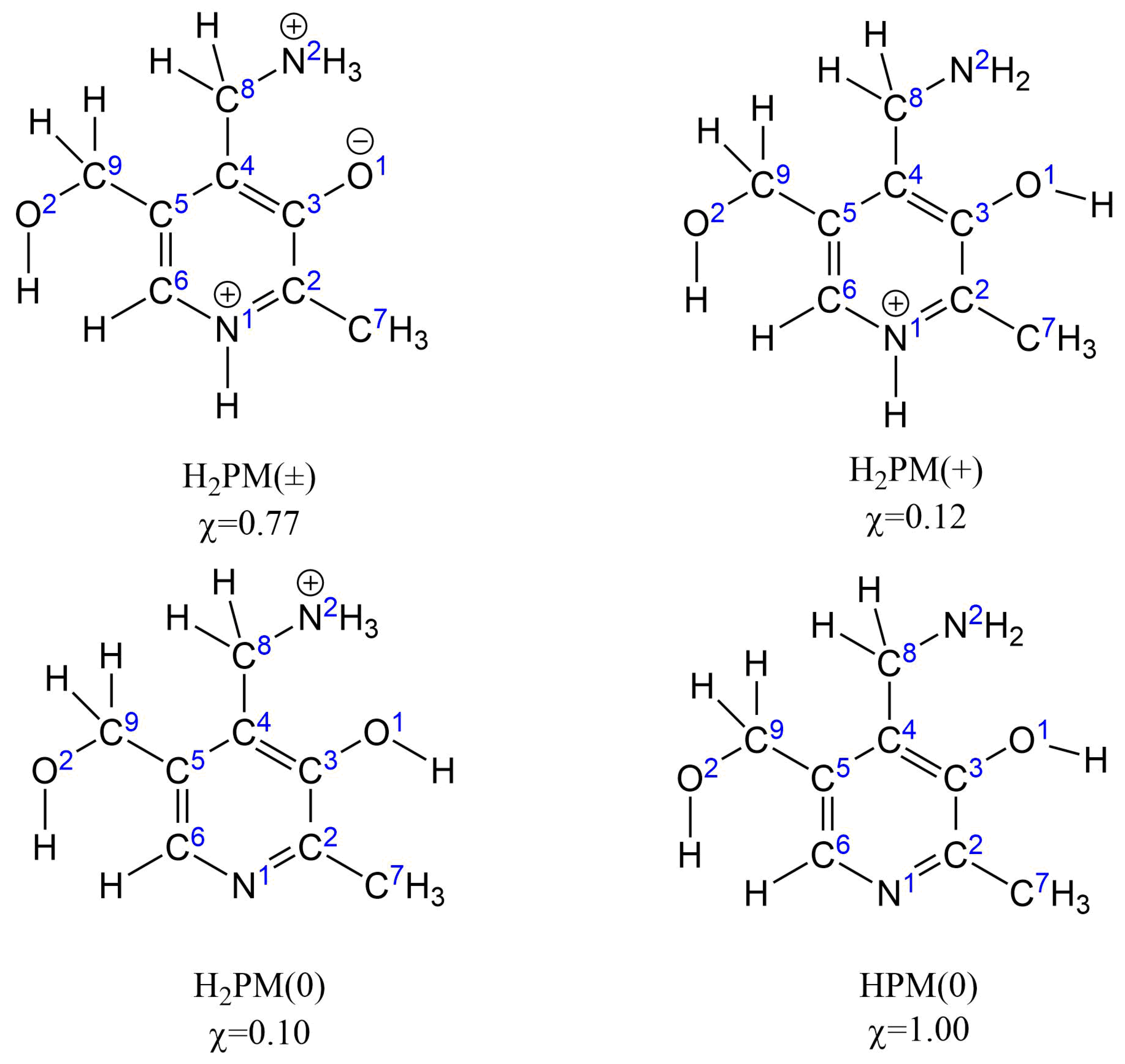{kind=link}
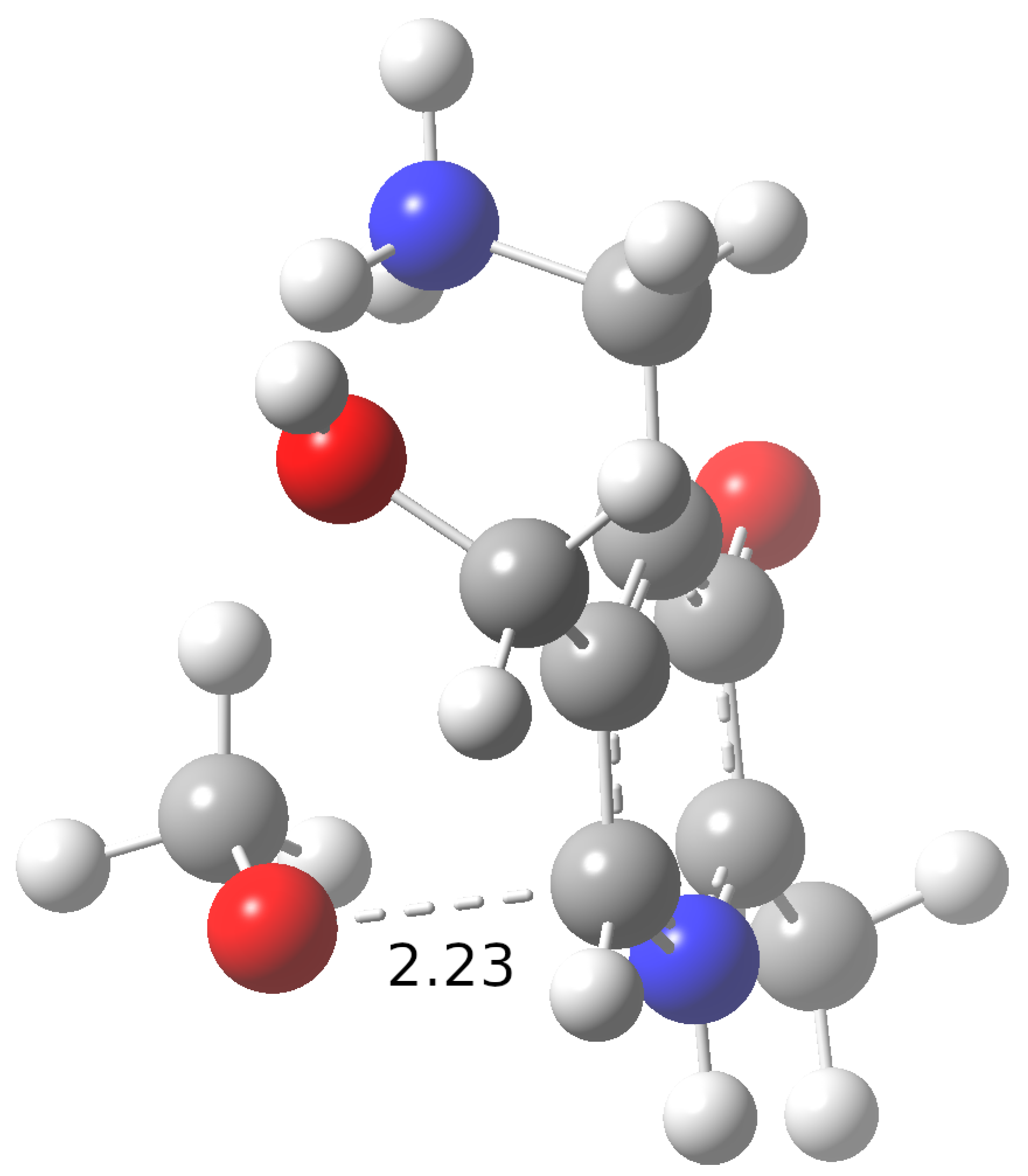{kind=link}
| H2PM(±) | H2PM(+) | H2PM(0) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| •OOH | •OOCH3 | •OCH3 | •OOH | •OOCH3 | •OCH3 | •OOH | •OOCH3 | •OCH3 | |
| HAT-C6 | 33.9 | 34.5 | 15.4 | 34.0 | 34.6 | 15.5 | 23.7 | 24.3 | 5.2 |
| HAT-C7 | −2.1 | −1.5 | −20.6 | 2.2 | 2.8 | −16.3 | 4.6 | 5.2 | −13.9 |
| HAT-C8 | −0.7 | −0.1 | −19.2 | −12.7 | −12.1 | −31.2 | 8.7 | 9.3 | −9.8 |
| HAT-C9 | −1.3 | −0.7 | −19.8 | −1.8 | −1.2 | −20.3 | 3.3 | 3.9 | −15.2 |
| HAT-N1 | −1.4 | −0.8 | −19.9 | 26.5 | 27.1 | 8.0 | - | - | − |
| HAT-N2 | 10.1 | 10.6 | −8.4 | 13.5 | 14.1 | −5.0 | 26.7 | 27.3 | 8.2 |
| HAT-O1 | - | - | - | 25.4 | 12.1 | −7.0 | 13.9 | 0.6 | −18.5 |
| HAT-O2 | 29.8 | 30.4 | 11.3 | 19.0 | 19.6 | 0.5 | 19.4 | 20.0 | 0.9 |
| RAF-N1 | - | - | - | - | - | - | 42.7 | - | 27.6 |
| RAF-C2 | −5.3 | −2.9 | −20.5 | 11.4 | 15.3 | −4.3 | 12.8 | 16.2 | −1.0 |
| RAF-C3 | 20.7 | 22.2 | 4.1 | 17.9 | 19.5 | 2.0 | 15.2 | 17.3 | 0.2 |
| RAF-C4 | 0.9 | 2.8 | −15.4 | 13.3 | 16.8 | −2.9 | 17.6 | 20.3 | 0.7 |
| RAF-C5 | 19.9 | 21.5 | 4.1 | 19.6 | 19.8 | 3.1 | 16.2 | 18.2 | 1.8 |
| RAF-C6 | −2.5 | 0.4 | −17.4 | 11.8 | 13.1 | −2.6 | 10.5 | 13.6 | −3.7 |
| SET | 26.3 | 28.1 | 20.8 | 62.0 | 63.8 | 56.5 | 41.5 | 43.3 | 36.0 |
| HPM(0) | |||
|---|---|---|---|
| •OOH | •OOCH3 | •OCH3 | |
| HAT-C6 | 23.1 | 24.6 | 4.8 |
| HAT-C7 | 5.6 | 7.0 | −12.8 |
| HAT-C8 | −0.2 | 1.2 | −18.5 |
| HAT-C9 | 0.3 | 1.7 | −18.0 |
| HAT-N2 | 19.2 | 20.6 | 0.9 |
| HAT-O1 | 6.9 | 8.3 | −11.4 |
| HAT-O2 | 19.7 | 21.1 | 1.3 |
| RAF-N1 | 42.6 | - | 29.3 |
| RAF-C2 | 11.1 | 15.0 | −1.8 |
| RAF-C3 | 14.8 | 19.3 | 0.0 |
| RAF-C4 | 13.8 | 17.2 | −1.8 |
| RAF-C5 | 16.1 | 18.8 | 1.1 |
| RAF-C6 | 10.7 | 15.6 | −3.6 |
| SET | 89.4 | 90.7 | 81.3 |
| H2PM(±) | H2PM(+) | H2PM(0) | HPM(0) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| •OOH | •OOCH3 | •OCH3 | •OOH | •OOCH3 | •OCH3 | •OCH3 | •OOH | •OCH3 | |
| HAT-C7 | 2.8 × 101 | 2.8 × 100 | 3.3 × 105 | - | - | 1.4 × 105 | 1.0 × 105 | - | 2.0 × 104 |
| HAT-C8 | 1.1 × 100 | 1.1 × 10−1 | 4.2 × 104 | 1.8 × 100 | 2.7 × 100 | 9.6 × 104 | 4.2 × 103 | 4.0 × 101 | 1.7 × 106 |
| HAT-C9 | 6.8 × 10−1 | 2.8 × 10−1 | 7.0 × 105 | 2.9 × 100 | 1.8 × 10−1 | 3.1 × 105 | 1.8 × 105 | 1.1 × 100 | 7.3 × 104 |
| HAT-N1 | - | - | 2.9 × 109 | - | - | - | - | - | - |
| HAT-N2 | - | - | 2.8 × 109 | - | - | 5.6 × 101 | - | - | 1.7 × 102 |
| HAT-O1 | - | - | - | - | - | 3.0 × 109 | 2.9 × 109 | - | 3.0 × 109 |
| HAT-O2 | - | - | - | - | - | 2.8 × 10−1 | 5.6 × 10-2 | - | - |
| RAF-C2 | 3.1 × 104 | 1.1 × 104 | 9.1 × 107 | - | - | 6.1 × 10−1 | 3.9 × 100 | - | 1.1 × 102 |
| RAF-C3 | - | - | - | - | - | - | 1.2 × 101 | - | 3.5 × 103 |
| RAF-C4 | 1.1 × 102 | - | 6.4 × 106 | - | - | 3.8 × 104 | 1.6 × 100 | - | 9.5 × 103 |
| RAF-C6 | 9.9 × 103 | 2.1 × 102 | 2.1 × 108 | - | - | 1.8 × 100 | 2.8 × 103 | - | 1.5 × 104 |
| Water | Pentyl Ethanoate | ||||
|---|---|---|---|---|---|
| •OOH | •OOCH3 | •OCH3 | •OOH | •OCH3 | |
| HAT-C7 | 2.1 × 101 (<0.1%) | 2.1 × 100 (<0.1%) | 2.8 × 105 (<0.1%) | ≈0 | 2.0 × 104 (<0.1%) |
| HAT-C8 | 1.1 × 100 (<0.1%) | 4.1 × 10−1 (<0.1%) | 4.4 × 104 (<0.1%) | 4.0 × 101 (97.3%) | 1.7 × 106 (<0.1%) |
| HAT-C9 | 8.7 × 10−1 (<0.1%) | 2.4 × 10−1 (<0.1%) | 6.0 × 105 (<0.1%) | 1.1 × 100 (2.7%) | 7.3 × 104 (<0.1%) |
| HAT-N1 | ≈0 | ≈0 | 2.2 × 109 (42.0%) | - | - |
| HAT-N2 | ≈0 | ≈0 | 2.2 × 109 (41.2%) | ≈0 | 1.7 × 102 (<0.1%) |
| HAT-O1 | ≈0 | ≈0 | 6.6 × 108 (12.4%) | ≈0 | 3.0 × 109 (99.9%) |
| HAT-O2 | ≈0 | ≈0 | 3.9 × 10-2 (<0.1%) | ≈0 | ≈0 |
| •OOH | •OOCH3 | •OCH3 | •OOH | •OCH3 | |
| RAF-C2 | 2.4 × 104 (75.6%) | 8.1 × 103 (98.0%) | 6.9 × 107 (1.3%) | ≈0 | 1.1 × 102 (<0.1%) |
| RAF-C3 | ≈0 | ≈0 | 1.2 × 100 (<0.1%) | ≈0 | 3.5 × 103 (<0.1%) |
| RAF-C4 | 8.6 × 101 (0.3%) | ≈0 | 4.9 × 106 (0.1%) | ≈0 | 9.5 × 103 (<0.1%) |
| RAF-C6 | 7.6 × 103 (24.0%) | 1.6 × 102 (1.9%) | 1.6 × 108 (3.1%) | ≈0 | 1.5 × 104 (<0.1%) |
| 3.2 × 104 | 8.3 × 103 | 5.3 × 109 | 4.2 × 101 | 3.0 × 109 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramis, R.; Ortega-Castro, J.; Caballero, C.; Casasnovas, R.; Cerrillo, A.; Vilanova, B.; Adrover, M.; Frau, J. How Does Pyridoxamine Inhibit the Formation of Advanced Glycation End Products? The Role of Its Primary Antioxidant Activity. Antioxidants 2019, 8, 344. https://doi.org/10.3390/antiox8090344
Ramis R, Ortega-Castro J, Caballero C, Casasnovas R, Cerrillo A, Vilanova B, Adrover M, Frau J. How Does Pyridoxamine Inhibit the Formation of Advanced Glycation End Products? The Role of Its Primary Antioxidant Activity. Antioxidants. 2019; 8(9):344. https://doi.org/10.3390/antiox8090344
Chicago/Turabian StyleRamis, Rafael, Joaquín Ortega-Castro, Carmen Caballero, Rodrigo Casasnovas, Antonia Cerrillo, Bartolomé Vilanova, Miquel Adrover, and Juan Frau. 2019. "How Does Pyridoxamine Inhibit the Formation of Advanced Glycation End Products? The Role of Its Primary Antioxidant Activity" Antioxidants 8, no. 9: 344. https://doi.org/10.3390/antiox8090344
APA StyleRamis, R., Ortega-Castro, J., Caballero, C., Casasnovas, R., Cerrillo, A., Vilanova, B., Adrover, M., & Frau, J. (2019). How Does Pyridoxamine Inhibit the Formation of Advanced Glycation End Products? The Role of Its Primary Antioxidant Activity. Antioxidants, 8(9), 344. https://doi.org/10.3390/antiox8090344