Antioxidant and Anti-Proliferative Properties of Hagenia abyssinica Roots and Their Potentially Active Components



 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Chemicals and Reagents
2.3. Extraction, Separation and Identification
2.4. In vitro Antioxidant Assays of H. abyssinica
2.4.1. 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Free Radical Scavenging Assay
2.4.2. ABTS+ Radical Cation Scavenging Activity Assay
2.4.3. Ferric-Ion Reducing Antioxidant Power (FRAP) Assay
2.5. Determinations of Total Phenolic Content (TPC)
2.6. Determinations of Total Flavonoid Content (TFC)
2.7. Anti-Proliferative Activity of H. abyssinica
2.8. Statistical Analysis
3. Results and Discussion
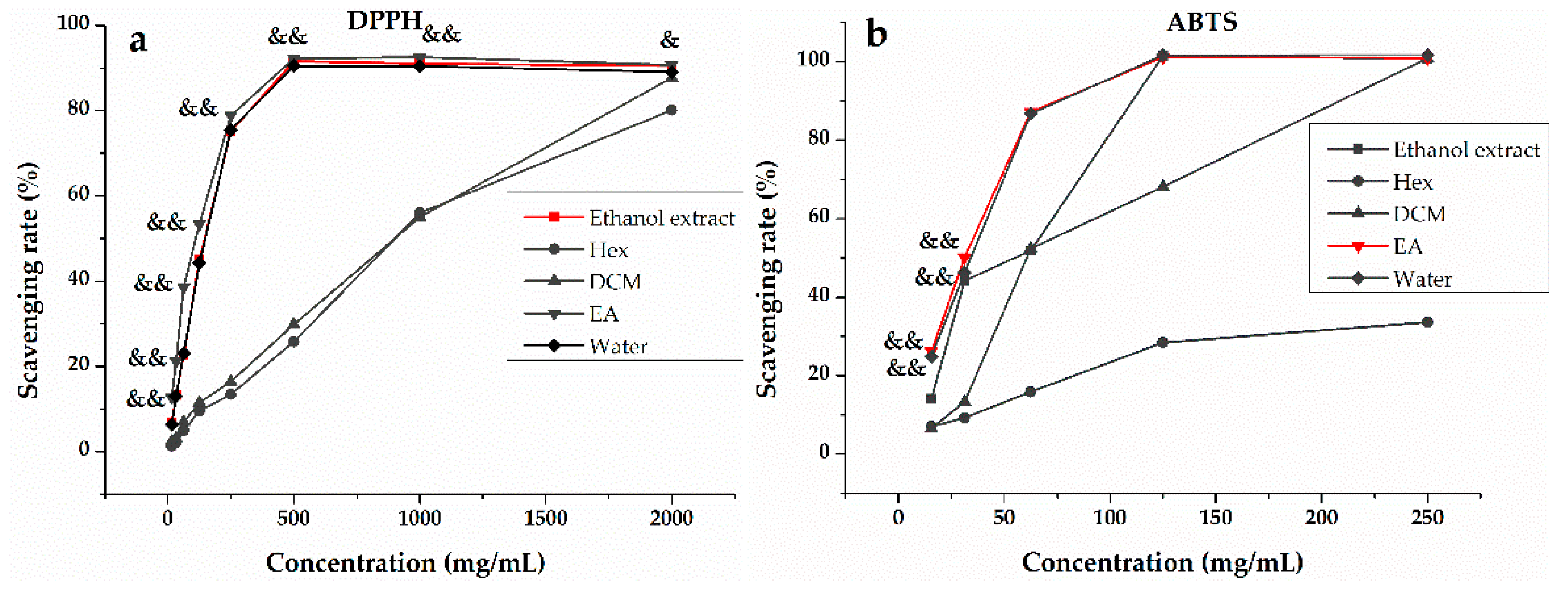
3.1. In Vitro Antioxidant Activity of H. abyssinica
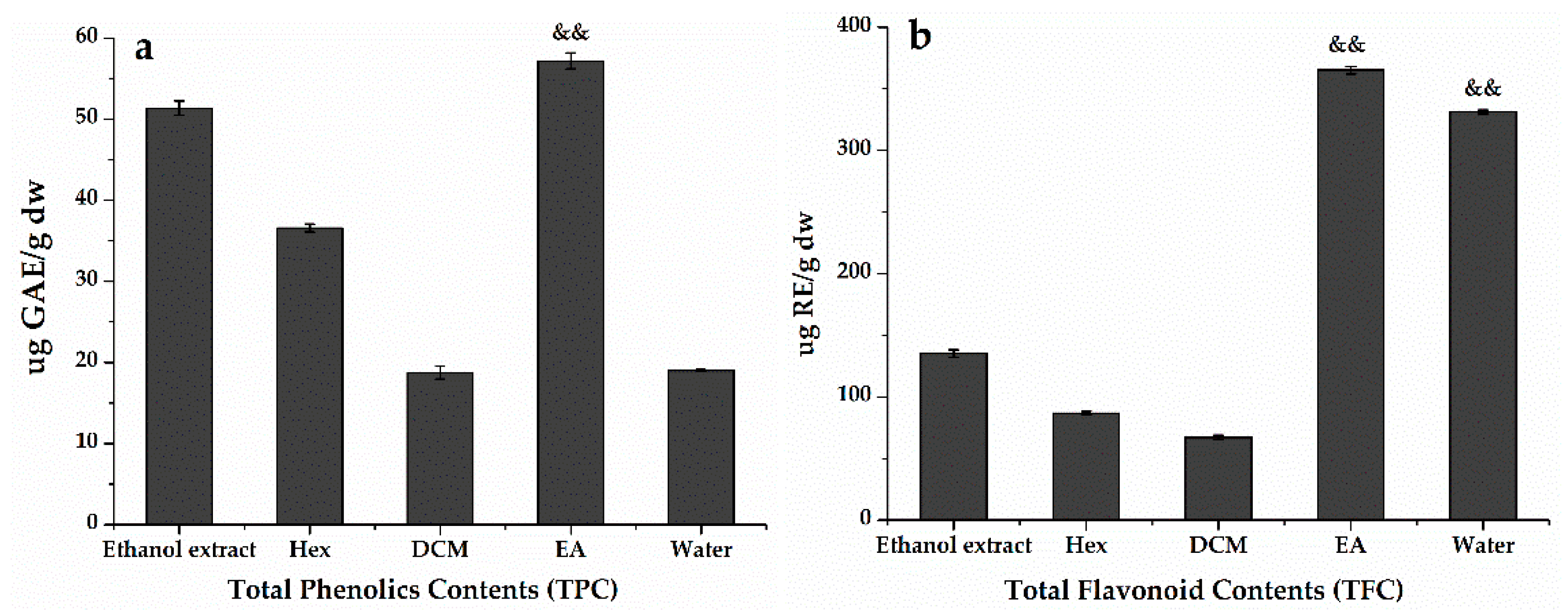
3.2. Total Phenolic and Flavonoid Contents of H. abyssinica
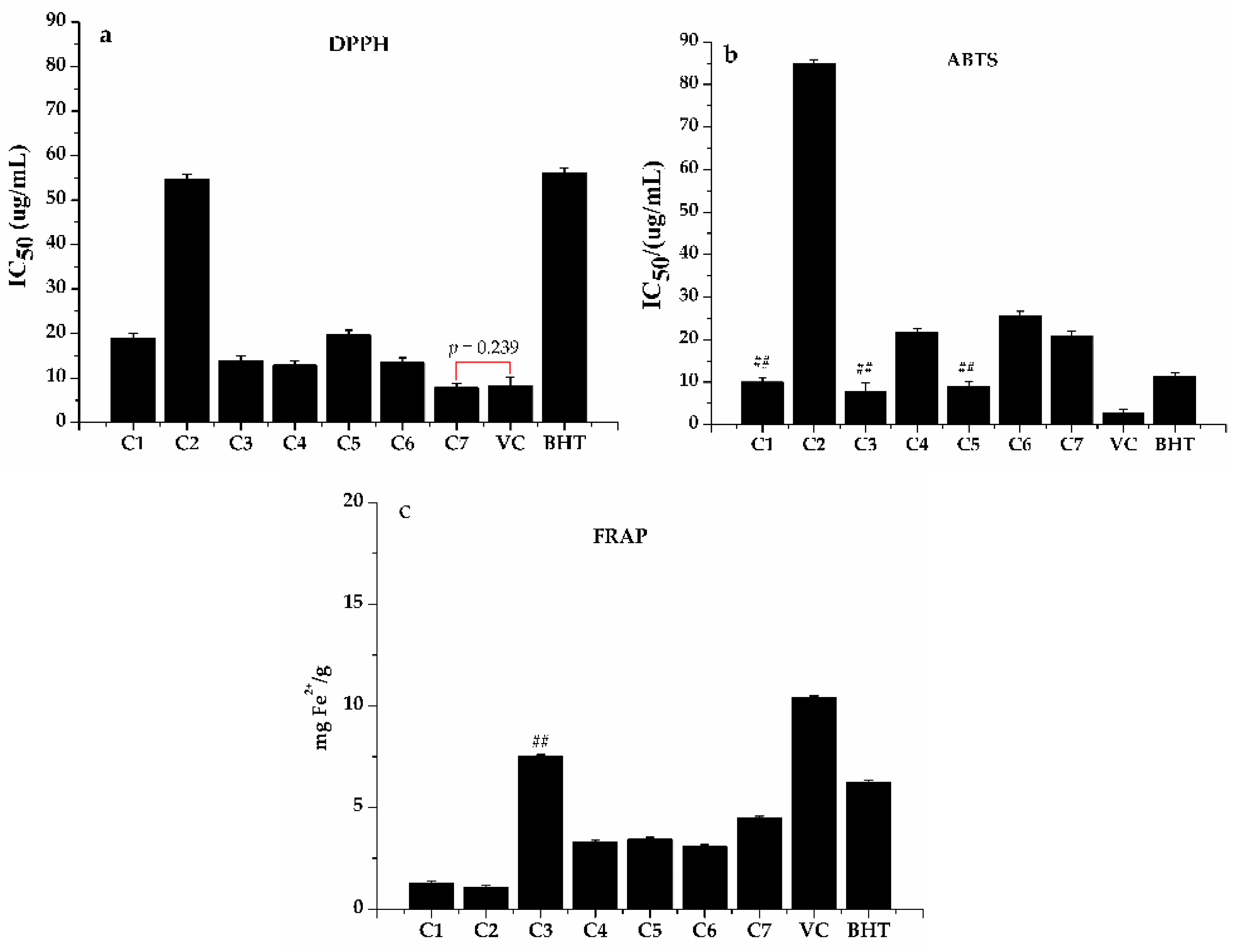
3.3. Anti-Proliferative Activity of H. abyssinica
3.4. Potential Antioxidants and Anti-Proliferative Compounds
3.5. The Structural Features and the Antioxidant and Anti-Proliferative Activities of Potential Active Compounds
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Feyissa, T.; Zhu, L.H.; Negash, L.; Welander, M. Regeneration and genetic transformation of Hagenia abyssinica (Bruce) J.F. Gmel. (Rosaceae) with rolB gene. Plant Cell Tiss. Organ Cult. 2007, 88, 277–288. [Google Scholar] [CrossRef]
- Negash, L. Indigenous Trees of Ethiopia: Biology, Uses and Propagation Techniques; The SLU Reprocentralen: Umeå, Sweden, 1995; pp. 125–147. [Google Scholar]
- Ayele, T.B.; Gailing, O.; Umer, M. Chloroplast DNA haplotype diversity and postglacial recolonization of Hagenia abyssinica (Bruce) J.F. Gmel. In Ethiopia. Plant Syst. Evol. 2009, 280, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Kokwaro, J.O. Medicinal Plants of East Africa; East African Literature Bureau: Nairobi, Kenya, 1976; pp. 292–294. [Google Scholar]
- Beentje, H.; Adamson, J.; Bhanderi, D. Kenya Trees Shrubs and Lianas; National Museums of Kenya: Nairobi, Kenya, 1994. [Google Scholar]
- Assefa, B.; Glatzel, G.; Buchmann, C. Ethnomedicinal uses of Hagenia abyssinica (Bruce) J.F. Gmel. Among rural communities of Ethiopia. J. Ethnobiolo. Ethnomed. 2010, 6, 20. [Google Scholar] [CrossRef] [Green Version]
- Woldemariam, T.Z.; Fell, A.F.; Linley, P.A.; Bibby, M.C.; Phillips, R.M. Evaluation of the anti-tumour action and acute toxicity of kosins from Hagenia abyssinica. J. Pharmaceut. Biomed. 1992, 10, 555–560. [Google Scholar] [CrossRef]
- Watt, J.M.; Breyer-Brandwijk, M.G. The Medicinal and Poisonous Plants of Southern and Eastern Africa, 2nd ed.; E. & S. Livingstone Ltd.: Scotland, UK, 1962. [Google Scholar]
- Farnsworth, N.R.; Bingel, A.S.; Cordell, G.A.; Crane, F.A.; Fong, F.A. Potential value of plants as source of new, antifertility agents I. J. Pharmacol. Sci. 1975, 64, 535–598. [Google Scholar] [CrossRef]
- Low, G.; Rogers, L.J.; Brumley, S.P.; Ehrlich, D. Visual deficits and retinotoxicity caused by the naturally occurring anthelmintics, Embelia ribes and Hagenia abyssinica. Toxicol. Appl. Pharmacol. 1985, 81, 220–230. [Google Scholar] [CrossRef]
- Abebe, D.; Ayehu, A. Medicinal plants and enigmatic health practices of Northern Ethiopia; Birhanena Selam Printing Enterprise: Addis Ababa, Ethiopia, 1993. [Google Scholar]
- Fassil, K.; Getachew, A. Utilization and conservation of medicinal plants in Ethiopia. In Proceedings of the Workshop on Development Utilization of Herbal Remedies in Ethiopia; Ethiopian Health and nutrition Institute: Addis Ababa, Ethiopia, 1996; pp. 46–52. [Google Scholar]
- Thomas, C. Oxygen Radicals and the Disease Process; CRC Press: Amsterdam, The Netherlands, 1998; pp. 1–282. [Google Scholar]
- Park, C.; Yeo, H.; Baskar, T.; Park, Y.; Park, J.; Lee, S.; Park, S. In vitro antioxidant and Antimicrobial properties of flower, leaf, and stem extracts of Korean Mint. Antioxidants 2019, 8, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matés, J.M.; Pérez-Gómez, C.; De Castro, I.N. Antioxidant enzymes and human diseases. Clin. Biochem. 1999, 32, 595–603. [Google Scholar] [CrossRef]
- Khan, N.; Afaq, F.; Mukhtar, H. Cancer chemoprevention through dietary antioxidants: Progress and promise. Antioxid. Redox Sign. 2008, 10, 475–510. [Google Scholar] [CrossRef] [PubMed]
- Lopez–Lazaro, M. Excessive superoxide anion generation plays a key role in carcinogenesis. Int. J. Cancer 2007, 120, 1378–1380. [Google Scholar] [CrossRef]
- Trachootham, D.; Alexandre, J.; Huang, P. Targeting cancer cells by ROS-mediated mechanisms: A radical therapeutic approach? Nat. Rev. Drug Discov. 2009, 8, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Trachootham, D.; Zhou, Y.; Zhang, H.; Demizu, Y.; Chen, Z.; Pelicano, H.; Chiao, P.J.; Achanta, G.; Arlinghaus, R.B.; Liu, J.; et al. Selective killing of oncogenically transformed cells through a ROS-mediated mechanism by β-phenylethyl isothiocyanate. Cancer Cell 2006, 10, 241–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nibret, E.; Wink, M. Trypanocidal and antileukaemic effects of the essential oils of Hagenia abyssinica, Leonotis ocymifolia, Moringa stenopetala, and their main individual constituents. Phytomedicine 2010, 17, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Lounasmaa, M.; Widén, C.J.; Huhtikangas, A. Phloroglucinol derivatives of Hagenia abyssinica. Phytochemistry 1973, 12, 2017–2025. [Google Scholar] [CrossRef]
- Lounasmaa, M.; Widén, C.J.; Huhtikangas, A. Phloroglucinol derivatives of Hagenia abyssinica. II. The structure determination of kosotoxin and protokosin. Acta Chem. Scand. B 1974, 28, 1200–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lounasmaa, M.; Varenne, P. Dérivés phloroglucinoliques d’ Hagenia abyssinica V1. Planta Med. 1978, 34, 153–159. [Google Scholar] [CrossRef]
- Wolde, T.; Bizuayehu, B.; Hailemariam, T.; Tiruha, K. Phytochemical analysis and antimicrobial activity of Hagenia abyssinica. Indian J. Pharm. Pharmacol. 2016, 3, 127–134. [Google Scholar] [CrossRef]
- Gul, M.Z.; Ahmad, F.; Kondapi, A.K.; Qureshi, I.A.; Ghazi, I.A. Antioxidant and anti-proliferative activities of Abrus precatorius leaf extracts-an in vitro study. BMC Complem. Altern. Med. 2013, 13, 53. [Google Scholar] [CrossRef] [Green Version]
- Tuasha, N.; Petros, B.; Asfaw, Z. Medicinal plants used by traditional healers to treat malignancies and other human ailments in Dalle District, Sidama Zone, Ethiopia. J. Ethnobiol. Ethnomed. 2018, 14, 15. [Google Scholar] [CrossRef] [Green Version]
- Yonas, A.; Zeleke, M.; Ahmed, Z.; Moti, Y.; Abdissa, B.; Sultan, S. Anti-schistosomal activities of Echinops kebericho Mesfin root and Hagenia abyssinica (Bruce) J.F Gmel flower part crude extracts in Swiss albino mice. Asian Pac. J. Trop. Med. 2018, 11, 26–31. [Google Scholar]
- Xu, Y.B.; Chen, G.L.; Guo, M.Q. Antioxidant and anti-inflammatory activities of the crude extracts of Moringa oleifera from Kenya and their correlations with flavonoids. Antioxidants 2019, 8, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzi, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as ameasure of ‘antioxidant power’: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.O.; Jeong, S.W.; Lee, C.Y. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem. 2003, 81, 321–326. [Google Scholar] [CrossRef]
- Anjum, S.; Gani, A.; Ahmad, M.; Shah, A.; Masoodi, F.A.; Shah, Y.; Asir, G. Antioxidant and anti-proliferative activity of walnut extract (Juglans regia L.) processed by different methods and identification of compounds using GC/MS and LC/MS technique. J. Food Process. Pres. 2016, 41, 1–9. [Google Scholar]
- Zhu, M.Z.; Wu, W.; Jiao, L.L.; Yang, P.F.; Guo, M.Q. Analysis of flavonoids in lotus (Nelumbo nucifera) leaves and their antioxidant activity using macroporous resin chromatography coupled with LC-MS/MS and antioxidant biochemical assays. Molecules 2015, 20, 10553–10565. [Google Scholar] [CrossRef] [Green Version]
- Schumacker, P.T. Sirt3 controls cancer metabolic reprogramming by regulating ROS and HIF. Cancer Cell 2011, 19, 299–300. [Google Scholar] [CrossRef] [Green Version]
- Li, F.L.; WU, X.; Liao, H.B.; Qiu, S.L.; Zhu, X.H.; Cui, L.; Wu, H. Dalbinol induces apoptosis of human colon cancer cells through ROS/ DVL/GSK-3β/β-catenin pathway. Chin. Pharm. Bull. 2016, 32, 1694–1698. [Google Scholar]
- Othman, A.; Mukhtar, N.J.; Ismail, N.S.; Chang, S.K. Phenolics, flavonoids content and antioxidant activities of 4 Malaysian herbal plants. Int. Food Res. J. 2014, 21, 759–766. [Google Scholar]
- Ma, Y.; Zhao, L.F.; Lv, Z.Y.; Sun, L.; Wu, W.; Liu, S.Y. Analysis of flavonoids and polyphenols in mulberry extracts by high-performance liquid chromatography quadrupole-qrbitrap mass spectrometry. J. Chin. Mass Spectrom. Soc. 2017, 38, 46–51. [Google Scholar]
- Yin, J.T.; Ma, Y.L.; Liang, C.; Gao, J.; Wang, H.R.; Zhang, L.T. A systematic study on the metabolites of dietary acacetin in vivo and in vitro based on UHPLC-Q-TOF-MS/MS analysis. J. Agric. Food Chem. 2019, 67, 5530–5543. [Google Scholar] [CrossRef] [PubMed]
- Marles, M.A.S.; Gruber, M.Y.; Scoles, G.J.; Muir, A.D. Pigmentation in the developing seed coat and seedling leaves of Brassica carinatais controlled at the dihydroflavonol reductase locus. Phytochemistry 2003, 62, 663–672. [Google Scholar] [CrossRef]
- Chang, C.C.; Lee, S.S. Rapid identification of flavonoid glycosides in Pasania kawakamii and Cyclobalanopsis morii via HPLC/MS and HPLC-SPE-NMR. Chem. Nat. Compd. 2012, 48, 689–692. [Google Scholar] [CrossRef]
- Guyot, S.; Vercauteren, J.; Cheynier, V. Structural determination of colourless and yellow dimers resulting from (+)-catechin coupling catalysed by grape polyphenoloxidase. Phytochemistry 1996, 42, 1279–1288. [Google Scholar] [CrossRef]
- Song, X.F.; Fan, B.L.; Zeng, X.L.; Li, T.T.; Shi, Y.M.; Zou, J.J.; Yang, J.; Chen, H.G. Simultaneous determination of seven active polyphenols in different Osmanthus fragrans cultivars by HPLC-MS/MS. Chin. J. Pharm. Anal. 2019, 39, 1811–1820. [Google Scholar]
- Sun, J.P.; Liang, F.; Yan, B.; Ping, L.; Duan, C.Q. Screening Non-colored Phenolics in red wines using liquid chromatography/ultraviolet and mass spectrometry/mass spectrometry libraries. Molecules 2007, 12, 679–693. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, Y.; Liang, Y.; Liang, K.; Zhang, F.; Xu, T.; Wang, M.; Song, H.; Liu, X.; Lu, B. Determination of phenolic acid profiles by HPLC-MS in vegetables commonly consumed in China. Food Chem. 2019, 276, 538–546. [Google Scholar] [CrossRef]
- Zhu, H.; Ma, X.; Kong, J.Y.; Zhang, M.L.; Kenttämaa, H.I. Identification of carboxylate, phosphate, and phenoxide functionalities in deprotonated molecules related to drug metabolites via ion–molecule reactions with water and diethylhydroxyborane. J. Am. Soc. Mass Spectrom. 2017, 28, 2189–2200. [Google Scholar] [CrossRef]
- Dao, P.T.A.; Quan, T.L.; Mai, N.T.T. Antioxidant constituents from the stem of Tetrastigma erusbescense Planch. (Vitaceae). Nat. Prod. Sci. 2014, 20, 22–28. [Google Scholar]
- Li, C.W.; Cui, C.B. One new and nine known flavonoids from choerospondias axillaries and their in vitro antitumor, anti-hypoxia and antibacterial activities. Molecules 2014, 19, 21363–21377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, M.X.; Chen, G.L.; Sun, B.Q.; Wu, J.L.; Guo, M.Q. Screening for natural inhibitors of human topoisomerases from medicinal plants with bio-affinity ultrafiltration and LC–MS. Phytochem. Rev. 2019, 1–31. [Google Scholar] [CrossRef]
- Singh, M.; Kaur, M.; Silakari, O. Flavones: An important scaffold for medicinal chemistry. Eur. J. Med. Chem. 2014, 84, 206–239. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.W.; Liu, P.Y.; Liu, D.; Sun, S.S.; Zhen, L.; YU, K.X.; Zhang, M.L.; Shi, Q.W. Research progress in structure-activity relationship of flavonoids. Chin. Tradit. Herb. Drugs 2015, 46, 3264–3271. [Google Scholar]
- Lopezlazaro, M. Flavonoids as anticancer agents: Structure-activity relationship study. Curr. Med. Chem. 2002, 2, 691. [Google Scholar]
- Habtemariam, S. Flavonoids as inhibitors or enhancers of the cytotoxicity of tumor necrosis factor-α in L-929 tumor cells. J. Nat. Prod. 1997, 60, 775–778. [Google Scholar] [CrossRef]
- Kurth, E.F.; Chan, F.L. Dihydroquercetin as an antioxidant. J. Am. Oil Chem. Soc. 1951, 28, 433–436. [Google Scholar] [CrossRef]
- Coskun, O.; Kanter, M.; Korkmaz, A.; Oter, S. Quercetin, a flavonoid antioxidant, prevents and protects streptozotocin-induced oxidative stress and β-cell damage in rat pancreas. Pharm. Res. 2005, 51, 117–123. [Google Scholar] [CrossRef]
- Fadeev, R.S.; Kaptsov, V.V.; Uminsky, A.A.; Akatov, V.S. Cytotoxic effect of dihydroquercetin and its derivatives in liposomal form and in the form of fat nanoscale emulsions. Biochem. Supp. 2011, 5, 45–50. [Google Scholar] [CrossRef]
- Massi, A.; Bortolini, O.; Ragno, D.; Bernardi, T.; Sacchetti, G.; Tacchini, M.; De Risi, C. Research progress in the modification of quercetin leading to anticancer agents. Molecules 2017, 22, 1270. [Google Scholar] [CrossRef]
- Liang, L.; Gao, C.; Luo, M.; Wang, W.; Zhao, C.; Zu, Y.; Efferth, T.; Fu, Y. Dihydroquercetin (DHQ) induced HO-1 and NQO1 expression against oxidative stress through the NRF2-dependent antioxidant pathway. J. Agric. Food Chem. 2013, 61, 2755–2761. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lim, M.J.; Kim, M.H.; Yu, C.H.; Yun, Y.S.; Ahn, J.; Song, J.Y. An effective strategy for increasing the radiosensitivity of human lung cancer cells by blocking nrf2-dependent antioxidant responses. Free Radic. Biol. Med. 2012, 53, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Gulati, N.; Laudet, B.; Zohrabian, V.M.; Murali, R.; Jhanwaruniyal, M. The anti-proliferative effect of quercetin in cancer cells is mediated via inhibition of the PI3K-AKT/PKB pathway. Anticancer Res. 2006, 26, 1177–1181. [Google Scholar] [PubMed]
- Hsu, Y.L.; Kuo, P.L.; Lin, C.C. Acacetin inhibits the proliferation of HepG2 by blocking cell cycle progression and inducing apoptosis. Biochem. Pharmacol. 2004, 67, 823–829. [Google Scholar] [CrossRef]
- Vijai, K.A.; Hala, N.E.; Shabana, I.K.; Troy, J.S.; Ikhlas, A.K.; Larry, A.W. Antioxidant constituents of Nymphaea caerulea flowers. Phytochemistry 2008, 69, 2061–2066. [Google Scholar]
- Amado, N.G.; Cerqueira, D.M.; Menezes, F.S.; Da, S.J.; Neto, V.M.; Abreu, J.G. Isoquercitrin isolated from Hyptis fasciculata reduces glioblastoma cell proliferation and changes β-catenin cellular localization. Anticancer Drug 2009, 20, 543–552. [Google Scholar] [CrossRef]
- Zhang, N.; Du, L.L.; Wang, D.; Liu, X.Q. Research progress of phenolic acids in traditional Chinese medicine. Mod. Chin. Med. 2006, 8, 25–28. [Google Scholar]
- Zhang, Y.; Wu, X.Q.; Ding, X.L. Study on the relationship between the structure of flavonoids and the scavenging efficacy of reactive oxygen species. Nat. Prod. Res. Dev. 1998, 10, 26–33. [Google Scholar]
- López-Posadas, R.; Ballester, I.; Abadía-Molina, A.C.; Suárez, M.D.; Zarzuelo, A.; Martínez-Augustin, O.; de Medina, F.S. Effectof flavonoids on rat splenocytes, a structure–activity relationship study. Biochem. Pharmacol. 2008, 76, 495–506. [Google Scholar] [CrossRef]
- Sun, J.; Huang, Y.; Sun, G.B.; Sun, X.B.; Qin, M.; Zhao, D. Study on in vitro antioxidant activity of flavonoids contained in hebei balmy chrysanthemum and structure-activity relationship. China J. Chin. Mater. Med. 2012, 37, 1958. [Google Scholar]
- Shen, K.H.; Hung, S.H.; Yin, L.T.; Huang, C.S.; Chao, C.H.; Liu, C.L.; Shih, Y.W. Acacetin, a flavonoid, inhibits the invasion and migration of human prostate cancer du145 cells via inactivation of the p38 MAPK signaling pathway. Mol. Cell. Biochem. 2010, 333, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Jeong, E.J.; Hwang, L.; Lee, M.; Lee, K.Y.; Ahn, M.J.; Sung, S.H. Neur-oprotective biflavonoids of Chamaecyparis obtusa leaves against glutamate-induced oxidative stress in HT22 hippocampal cells. Food Chem. Toxicol. 2014, 64, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, D.; Panda, P.K.; Tripathy, G.; Pattanayak, J.R.N. Anticancer activity of biflavonoids from lonicera japonica and benincasa hispida on human cancer cell lines. J. Pharm. Res. 2009, 2, 983–985. [Google Scholar]
- Han, J.; Weng, X.; Bi, K. Antioxidants from a Chinese medicinal herb-Lithospermum erythrorhizon. Food Chem. 2008, 106, 2–10. [Google Scholar] [CrossRef]
- Balasundram, N.; Sundram, K.; Samman, S. Phenolic compounds in plants and AGRI-industrial by-products: Antioxidant activity, occurrence, and potential uses. Food Chem. 2006, 99, 191–203. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, K.Z.; Song, J.R.; Hang, J.; Nie, W. Theoretical study on antioxidant activity of five phenolic acids. Food Sci. 2011, 32, 36–39. [Google Scholar]
- Lin, H.H.; Chen, J.H.; Huang, C.C.; Wang, C.J. Apoptotic effect of 3,4-dihydroxybenzoic acid on human gastric carcinoma cells involving JNK/p38 mapk signaling activation. Int. J. Cancer 2007, 120, 2306–2316. [Google Scholar] [CrossRef]
- Xiao, X.N.; Xu, W.H.; Zuo, D.; Liao, X.; Ming, J. The syner gistic antioxidant effect and structure-activity relationship of six flavonoids. Food Mach. 2017, 11, 7–10. [Google Scholar]
- Riceevans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Gong, J.Y.; Huang, J.; Xiao, G.N.; Chen, F.; Lee, B.; Ge, Q.; You, Y.; Liu, S. Antioxidant capacities of fractions of bamboo shaving extract and their antioxidant components. Molecules 2016, 21, 996. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Peak No. | Rt/min | [M – H]− | MS/MS Spectrum | Identification | Ref. |
|---|---|---|---|---|---|---|
| 1 | 6 | 46.47 | 303.24 | 285.13, 241.10, 217.08, 199.16, 174.68, 125.18 | dihydroquercetin | [38] |
| 2 | 9 | 76.12 | 283.23 | 268.06, 240.03, 239.03, 211.03, 212.04, 151.00, 117.03, 107.01 | acacetin | [39] |
| 3 | 8 | 66.76 | 301.50 | 179.11,151.07 | quercetin | [40] |
| 4 | 5 | 42.90 | 463.23 | 301.24, 300.14 | isoquercitin | [41] |
| 5 | 7 | 56.62 | 575.06 | 445.00, 423.00, 394.00, 271.00, 243.00, 229.00, 137.00, 125.00 | dehydrodicatechin A | [42] |
| 6 | 4 | 35.78 | 193.10 | 178.10,149.12,134.08 | trans-ferulic acid | [43] |
| 7 | 2 | 24.15 | 179.48 | 135.21 | caffeic acid | [44] |
| 8 | 1 | 14.55 | 153.11 | 109.02 | 3, 4-dihydroxybenzoic acid | [45] |
| 9 | 3 | 29.22 | 195.16 | 180.23, 136.11 | 2-methoxyterephthalic acid | [46] |
| 1H-NMR | 1H-1H COSYa | 13C-NMR | HMBCb (H→C) | |
|---|---|---|---|---|
| 2C | 3.98 (1H, m) | 4C-H | 79.5 | C-3C, 4C, 6′B |
| 3C | 3.98 (1H, m) | 3C-H | 66.9 | C-2C, 4C, 1′B |
| 4C | Ha: 2.50 (1H, m, 4axC) Hb: 2.94 (1H, m, 4exC) | 3C-H | 27.8 | C-2C, 3C, 4aA, 5A |
| 4aA | - | 100.5 | - | |
| 5A | - | 156.4 | - | |
| 6A | 5.54 (1H, d, J = 2.0 Hz) | 8A | 97.1 | C-4aA, 5A, 7A, 8A |
| 7A | - | 157.7 | - | |
| 8A | 5.90 (1H, d, J = 2.0 Hz) | 6A | 95.8 | C-4aA, 6A, 7A, 8aA |
| 8aA | - | 158.0 | - | |
| 1′B | - | 89.9 | - | |
| 2′B | Ha: 2.50 (1H, m) Hb: 2.67 (1H, m) | 3′B-H | 45.9 | C-2C, 1′B, 3′B, 6′B |
| 3′B | - | 2′B-H, 4′B-H | 95.3 | - |
| 4′B | - | 3′B-H | 194.1 | - |
| 5′B | 6.43 (1H, s) | 3′B-H | 112.9 | C-2′B, 1′B, 3′B, 6′B |
| 6′B | - | 164.4 | - | |
| 2F | 4.93 (1H, d, J = 6.5 Hz) | 83.5 | C-3F, 4F, 1′E, 2′E, 6′E, 8aD | |
| 3F | 4.11 (1H, m) | 67.9 | C-4aD | |
| 4F | Ha: 2.60 (1H, dd, J = 7.5, 16.5 Hz, 4axF) Hb: 2.85 (1H, dd, J = 5.0, 16.5 Hz, 4exF) | 28.3 | C-2F, 3F, 4aD, 5D, 8aD | |
| 4aD | - | 103.9 | ||
| 5D | - | 166.2 | ||
| 6D | 6.16 (1H, s) | 90.9 | C-5D, 8D | |
| 7D | - | 168.1 | ||
| 8D | - | 105.6 | ||
| 8aD | - | 155.1 | ||
| 1′E | - | 131.3 | ||
| 2′E | 6.84 (1H, d, J = 2.0 Hz, H-2′E) | 5′E | 114.8 | C-2F, 4′E, 6′E |
| 3′E | - | 146.5 | ||
| 4′E | - | 146.6 | ||
| 5′E | 6.79 (1H, dd, J = 8.0 Hz, H-) | 2′E, 6′E | 116.3 | C-1′E, 3′E |
| 6′E | 6.74 (1H, dd, J = 2.0, 8.0 Hz) | 5′E | 119.7 | C-2F, 2′E, 4′E |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, M.; Chen, G.; Zhang, Y.; Nahar, L.; Sarker, S.D.; Hu, G.; Guo, M. Antioxidant and Anti-Proliferative Properties of Hagenia abyssinica Roots and Their Potentially Active Components. Antioxidants 2020, 9, 143. https://doi.org/10.3390/antiox9020143
Fan M, Chen G, Zhang Y, Nahar L, Sarker SD, Hu G, Guo M. Antioxidant and Anti-Proliferative Properties of Hagenia abyssinica Roots and Their Potentially Active Components. Antioxidants. 2020; 9(2):143. https://doi.org/10.3390/antiox9020143
Chicago/Turabian StyleFan, Minxia, Guilin Chen, Yongli Zhang, Lutfun Nahar, Satyajit Dey Sarker, Guangwan Hu, and Mingquan Guo. 2020. "Antioxidant and Anti-Proliferative Properties of Hagenia abyssinica Roots and Their Potentially Active Components" Antioxidants 9, no. 2: 143. https://doi.org/10.3390/antiox9020143