Antioxidants Reduce Muscular Dystrophy in the dy2J/dy2J Mouse Model of Laminin α2 Chain-Deficient Muscular Dystrophy

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Human Tissues
2.3. Treatments
2.4. Stand Ups and Grip Strength
2.5. Tissue Collection
2.6. Histology and Immunohistochemistry
2.7. Morphometric Evaluation of Fibrosis by Fast Green and Sirius Red Staining
2.8. Colorimetric Evaluation of Fibrosis by Fast Green/Sirius Red Staining
2.9. Dihydroethidium Staining for ROS Detection
2.10. Real Time PCR Analysis
2.11. Statistical Analysis and Data Availability
3. Results
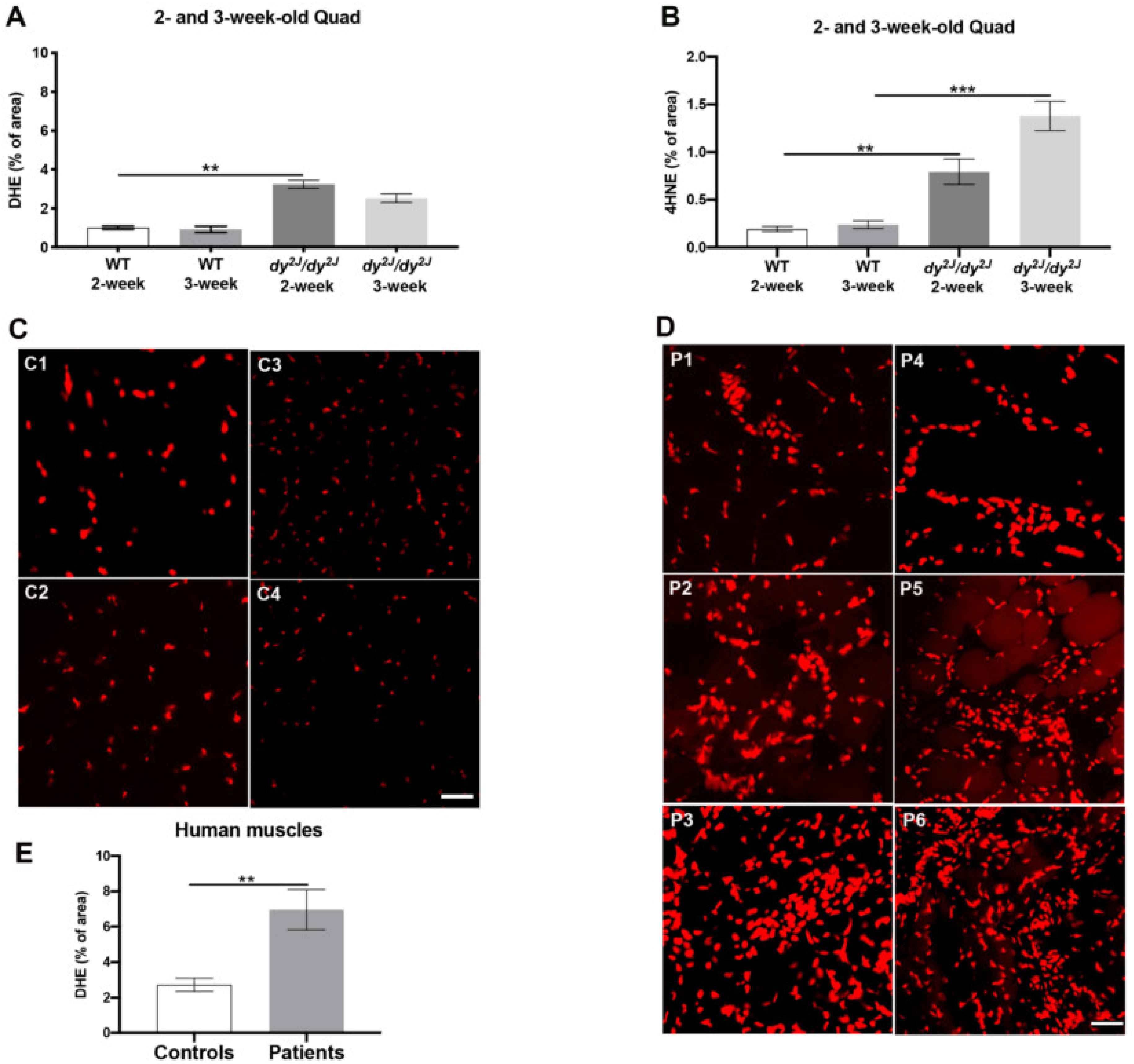
3.1. Significantly Increased ROS Levels in Skeletal Muscles of dy2J/dy2J Mice and LAMA2-CMD Patients
3.2. Muscle Force Loss is Prevented by NAC Treatment
3.3. Skeletal Muscle Histology is Preserved by Both NAC and Vitamin E Treatment
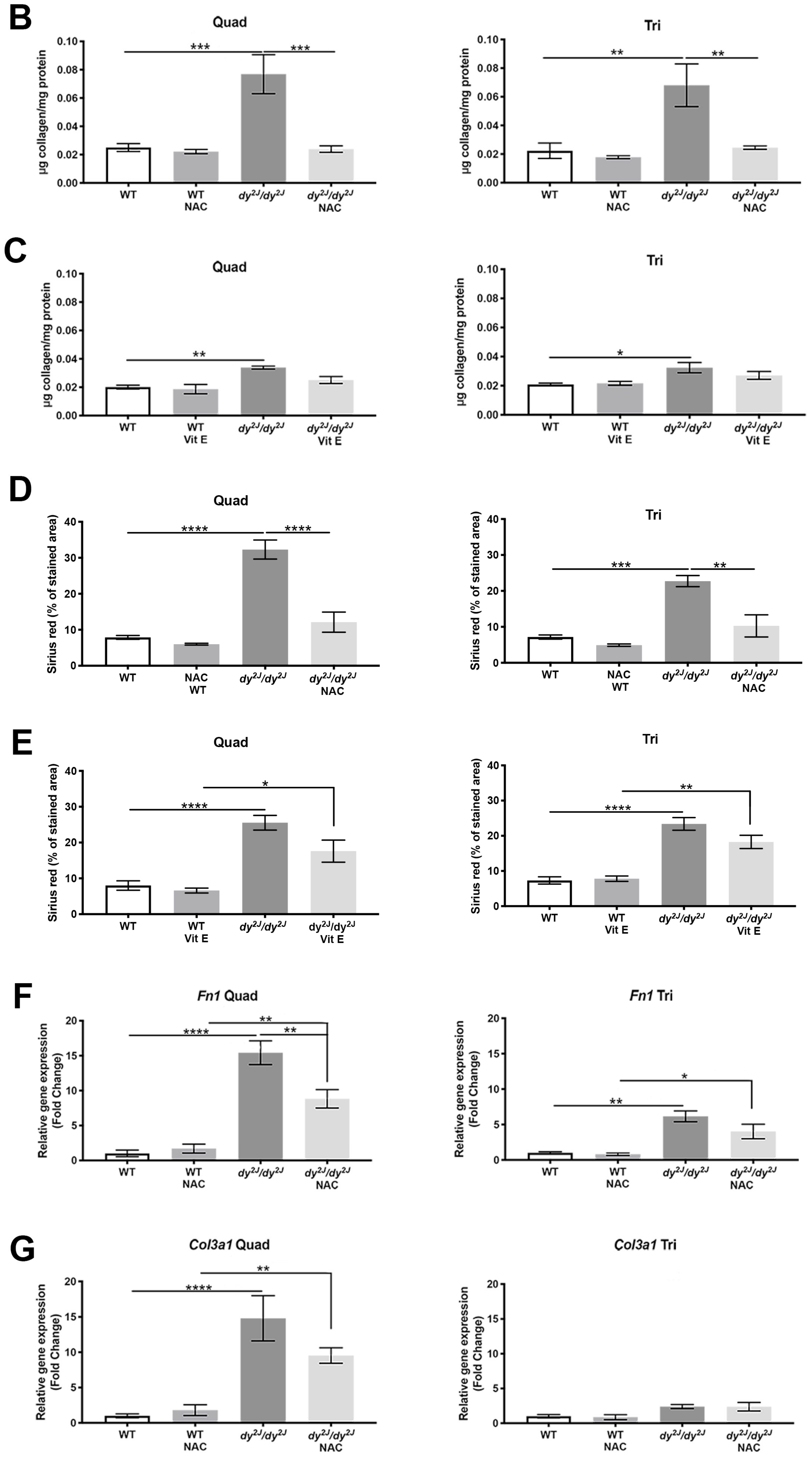
3.4. Fibrosis is Prevented in NAC Treated dy2J/dy2J Muscles
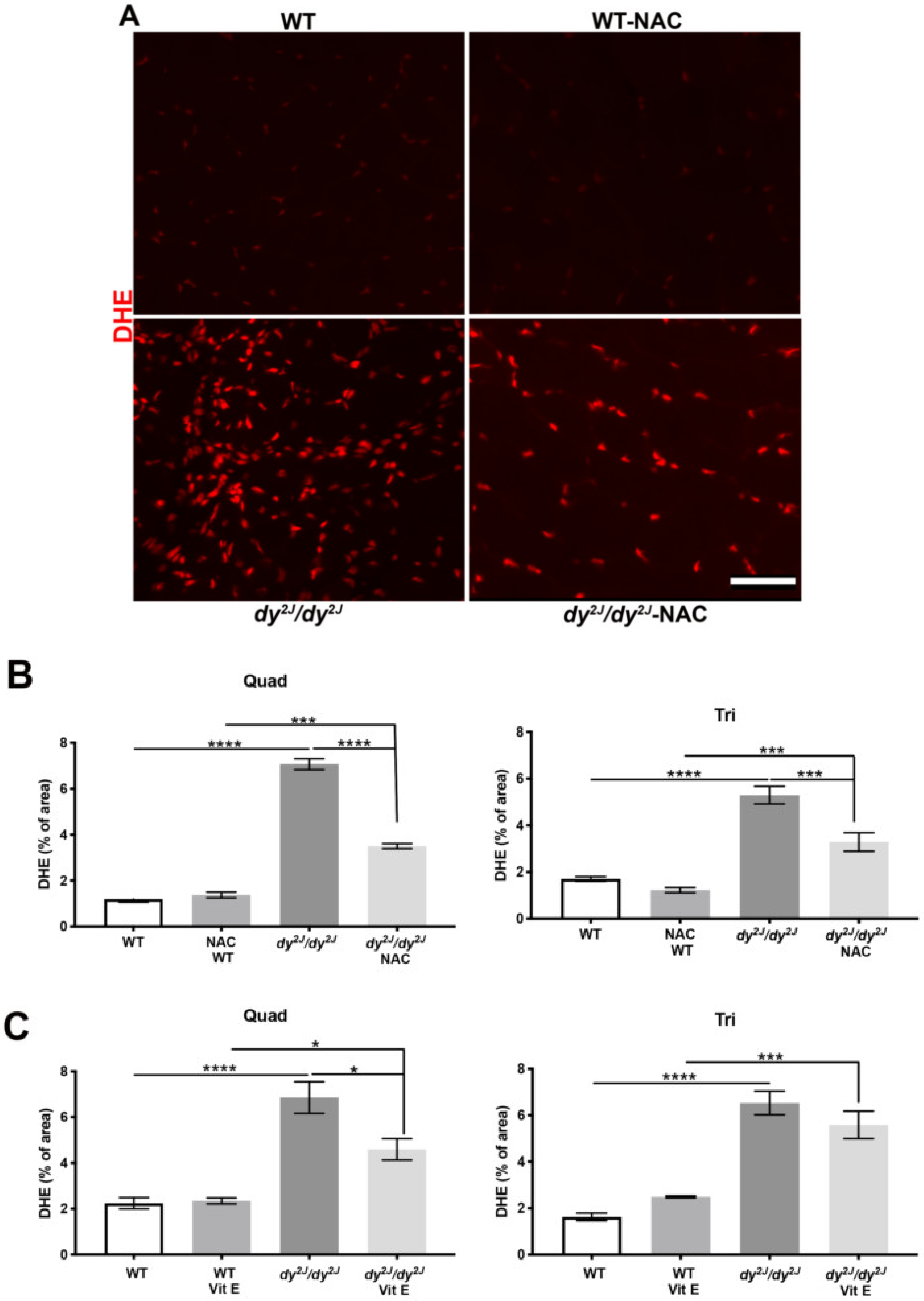
3.5. Inflammation and Increased ROS Levels are Prevented in Response to NAC and Vitamin E
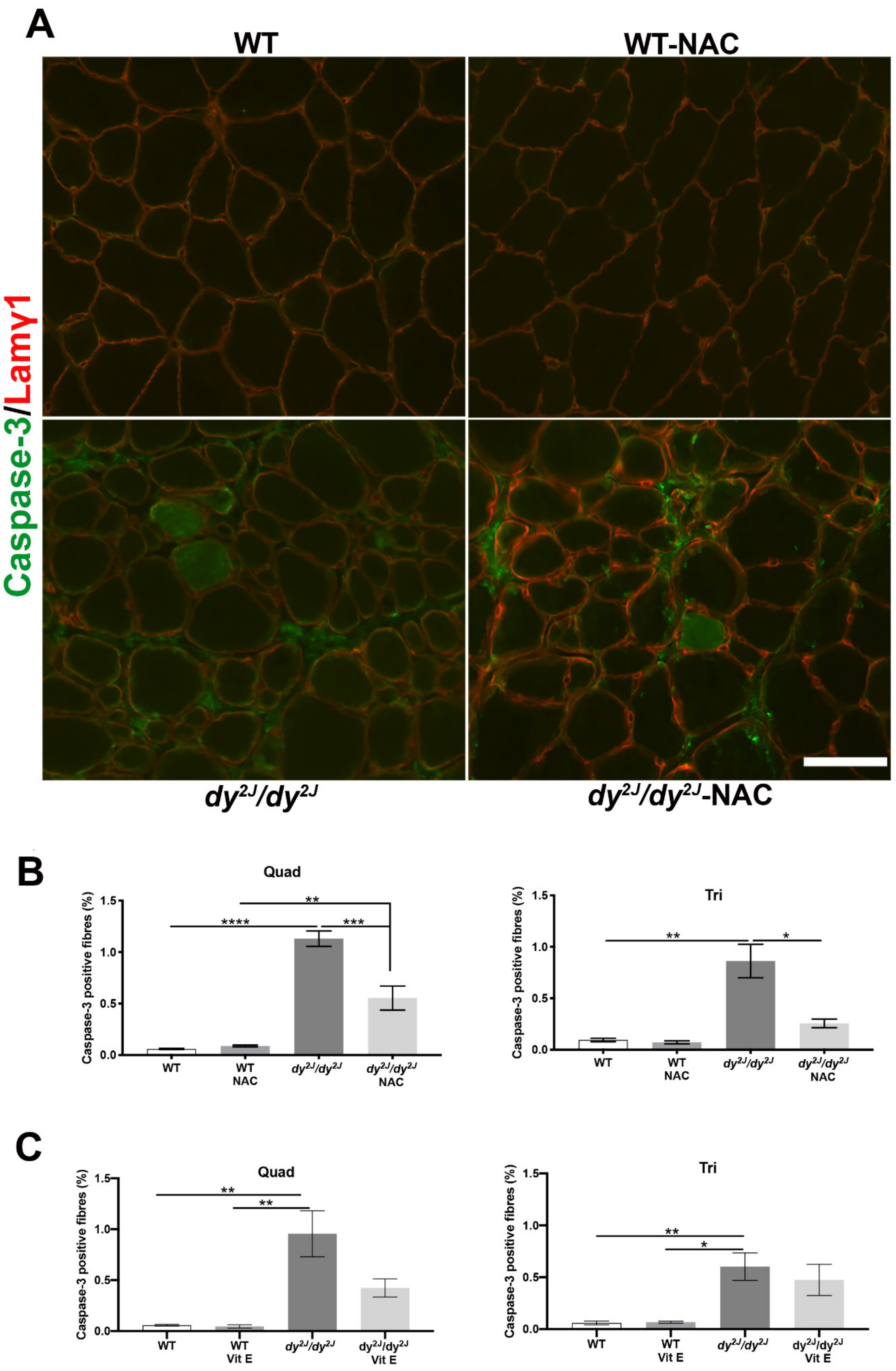
3.6. Apoptosis is Prevented upon NAC Treatment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Durbeej, M. Laminin-alpha2 Chain-Deficient Congenital Muscular Dystrophy: Pathophysiology and Development of Treatment. Curr. Top. Membr. 2015, 76, 31–60. [Google Scholar] [PubMed]
- Patton, B.L.; Miner, J.H.; Chiu, A.Y.; Sanes, J.R. Distribution and function of laminins in the neuromuscular system of developing, adult, and mutant mice. J. Cell. Biol. 1997, 139, 1507–1521. [Google Scholar] [CrossRef] [PubMed]
- Petrof, B.J.; Shrager, J.B.; Stedman, H.H.; Kelly, A.M.; Sweeney, H.L. Dystrophin protects the sarcolemma from stresses developed during muscle contraction. Proc. Natl. Acad. Sci. USA 1993, 90, 3710–3714. [Google Scholar] [CrossRef] [PubMed]
- Gawlik, K.I.; Durbeej, M. Skeletal muscle laminin and MDC1A: Pathogenesis and treatment strategies. Skelet. Muscle 2011, 1, 9. [Google Scholar] [CrossRef] [PubMed]
- Miyagoe, Y.; Hanaoka, K.; Nonaka, I.; Hayasaka, M.; Nabeshima, Y.; Arahata, K.; Nabeshima, Y.; Takeda, S. Laminin alpha2 chain-null mutant mice by targeted disruption of the Lama2 gene: A new model of merosin (laminin 2)-deficient congenital muscular dystrophy. FEBS Lett. 1997, 415, 33–39. [Google Scholar] [CrossRef]
- Xu, H.; Wu, X.R.; Wewer, U.M.; Engvall, E. Murine muscular dystrophy caused by a mutation in the laminin alpha 2 (Lama2) gene. Nat. Genet. 1994, 8, 297–302. [Google Scholar] [CrossRef]
- Sunada, Y.; Bernier, S.M.; Utani, A.; Yamada, Y.; Campbell, K.P. Identification of a novel mutant transcript of laminin alpha 2 chain gene responsible for muscular dystrophy and dysmyelination in dy2J mice. Hum Mol. Genet. 1995, 4, 1055–1061. [Google Scholar] [CrossRef]
- Pasteuning-Vuhman, S.; Putker, K.; Tanganyika-de Winter, C.L.; van der Boertje Meulen, J.W.; van Vliet, L.; Overzier, M.; Plomp, J.J.; Aartsma-Rus, A.; van Putten, M. Natural disease history of the dy2J mouse model of laminin alpha2 (merosin)-deficient congenital muscular dystrophy. PLoS ONE 2018, 13, e0197388. [Google Scholar] [CrossRef]
- De Oliveira, B.M.; Matsumura, C.Y.; Fontes-Oliveira, C.C.; Gawlik, K.I.; Acosta, H.; Wernhoff, P.; Durbeej, M. Quantitative proteomic analysis reveals metabolic alterations, calcium dysregulation, and increased expression of extracellular matrix proteins in laminin alpha2 chain-deficient muscle. Mol. Cell. Proteomics 2014, 13, 3001–3013. [Google Scholar] [CrossRef]
- Hager, M.; Bigotti, M.G.; Meszaros, R.; Carmignac, V.; Holmberg, J.; Allamand, V.; Åkerlund, M.; Kalamajski, S.; Brancaccio, A.; Mayer, U.; et al. Cib2 binds integrin alpha7Bbeta1D and is reduced in laminin alpha2 chain-deficient muscular dystrophy. J. Biol. Chem. 2008, 283, 24760–24769. [Google Scholar] [CrossRef]
- Fontes-Oliveira, C.C.; Steinz, M.; Schneiderat, P.; Mulder, H.; Durbeej, M. Bioenergetic Impairment in Congenital Muscular Dystrophy Type 1A and Leigh Syndrome Muscle Cells. Sci. Rep. 2017, 7, 45272. [Google Scholar] [CrossRef] [PubMed]
- Rando, T.A. Oxidative stress and the pathogenesis of muscular dystrophies. Am. J. Phys. Med. Rehabil. 2002, 81, S175–S186. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.H.; Ow, J.R.; Yang, N.D.; Taneja, R. Oxidative Stress-Mediated Skeletal Muscle Degeneration: Molecules, Mechanisms, and Therapies. Oxid. Med. Cell. Longev. 2016. [Google Scholar] [CrossRef]
- Kerksick, C.; Willoughby, D. The antioxidant role of glutathione and N-acetyl-cysteine supplements and exercise-induced oxidative stress. J. Int. Soc. Sports Nutr. 2005, 2, 38–44. [Google Scholar] [CrossRef]
- Heard, K.J. Acetylcysteine for acetaminophen poisoning. N. Engl. J. Med. 2008, 359, 285–292. [Google Scholar] [CrossRef]
- Tirouvanziam, R.; Conrad, C.K.; Bottiglieri, T.; Herzenberg, L.A.; Moss, R.B.; Herzenberg, L.A. High-dose oral N-acetylcysteine, a glutathione prodrug, modulates inflammation in cystic fibrosis. Proc. Natl. Acad. Sci. USA 2006, 103, 4628–4633. [Google Scholar] [CrossRef]
- Slattery, J.; Kumar, N.; Delhey, L.; Berk, M.; Dean, O.; Spielholz, C.; Frye, R. Clinical trials of N-acetylcysteine in psychiatry and neurology: A systematic review. Neurosci. Biobehav. Rev. 2015, 55, 294–321. [Google Scholar]
- Howard, A.C.; McNeil, A.K.; McNeil, P.L. Promotion of plasma membrane repair by vitamin E. Nat. Commun. 2011, 2, 1–8. [Google Scholar] [CrossRef]
- Hill, K.E.; Montine, T.J.; Motley, A.K.; Li, X.; May, J.M.; Burk, R.F. Combined deficiency of vitamins E and C causes paralysis and death in guinea pigs. Am. J. Clin. Nutr. 2003, 77, 1484–1488. [Google Scholar] [CrossRef]
- Thomas, P.K.; Cooper, J.M.; King, R.H.; Workman, J.M.; Schapira, A.H.; Goss-Sampson, M.A.; Muller, D.P. Myopathy in vitamin E deficient rats: Muscle fibre necrosis associated with disturbances of mitochondrial function. J. Anat. 1993, 183, 451–461. [Google Scholar] [PubMed]
- Korner, Z.; Fontes-Oliveira, C.C.; Holmberg, J.; Carmignac, V.; Durbeej, M. Bortezomib partially improves laminin alpha2 chain-deficient muscular dystrophy. Am. J. Pathol. 2014, 184, 1518–1528. [Google Scholar] [CrossRef] [PubMed]
- Leiter, J.R.; Peeler, J.; Anderson, J.E. Exercise-induced muscle growth is muscle-specific and age-dependent. Muscle Nerve. 2011, 43, 828–838. [Google Scholar] [CrossRef] [PubMed]
- Gawlik, K.I.; Harandi, V.M.; Cheong, R.Y.; Petersen, A.; Durbeej, M. Laminin alpha1 reduces muscular dystrophy in dy(2J) mice. Matrix Biol. 2018, 70, 36–49. [Google Scholar] [CrossRef]
- Fontes-Oliveira, C.C.; Soares Oliveira, B.M.; Korner, Z.; Harandi, V.M.; Durbeej, M. Effects of metformin on congenital muscular dystrophy type 1A disease progression in mice: A gender impact study. Sci. Rep. 2018, 8, 16302. [Google Scholar] [CrossRef]
- Nguyen, H.X.; Tidball, J.G. Null mutation of gp91phox reduces muscle membrane lysis during muscle inflammation in mice. J. Physiol. 2003, 553, 833–841. [Google Scholar] [CrossRef]
- Moreira Soares Oliveira, B.; Gawlik, K.I.; Durbeej, M.; Holmberg, J. Exploratory Profiling of Urine MicroRNAs in the dy(2J)/dy(2J) Mouse Model of LAMA2-CMD: Relation to Disease Progression. PLoS Curr. 2018, 10. [Google Scholar]
- Victor, V.M.; Rocha, M.; De la Fuente, M. Regulation of macrophage function by the antioxidant N-acetylcysteine in mouse-oxidative stress by endotoxin. Int. Immunopharmacol. 2003, 3, 97–106. [Google Scholar] [CrossRef]
- Cao, L.; Li, L.; Zuo, Z. N-acetylcysteine reverses existing cognitive impairment and increased oxidative stress in glutamate transporter type 3 deficient mice. Neuroscience 2012, 220, 85–89. [Google Scholar] [CrossRef]
- Durieux, A.M.; Fernandes, C.; Murphy, D.; Labouesse, M.A.; Giovanoli, S.; Meyer, U.; Li, Q.; So, P.W.; McAlonan, G. Targeting Glia with N-Acetylcysteine Modulates Brain Glutamate and Behaviors Relevant to Neurodevelopmental Disorders in C57BL/6J Mice. Front. Behav. Neurosci. 2015, 9, 343. [Google Scholar] [CrossRef]
- Mancio, R.D.; Hermes, T.A.; Macedo, A.B.; Mizobuti, D.S.; Valduga, A.H.; Rupcic, I.F.; Minatel, E. Vitamin E treatment decreases muscle injury in mdx mice. Nutrition 2017, 43–44, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Korner, Z.; Durbeej, M. Bortezomib Does Not Reduce Muscular Dystrophy in the dy2J/dy2J Mouse Model of Laminin alpha2 Chain-Deficient Muscular Dystrophy. PLoS ONE 2016, 11, e0146471. [Google Scholar] [CrossRef]
- Kemaladewi, D.U.; Benjamin, J.S.; Hyatt, E.; Ivakine, E.A.; Cohn, R.D. Increased polyamines as protective disease modifiers in congenital muscular dystrophy. Hum. Mol. Genet. 2018, 27, 1905–1912. [Google Scholar] [CrossRef] [PubMed]
- Geranmayeh, F.; Clement, E.; Feng, L.H.; Sewry, C.; Pagan, J.; Mein, R.; Abbs, S.; Brueton, L.; Childs, A.M.; Jungbluth, H.; et al. Genotype-phenotype correlation in a large population of muscular dystrophy patients with LAMA2 mutations. Neuromuscul. Disord. 2010, 20, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Quijano-Roy, S.; Sparks, S.E.; Rutkowski, A. LAMA2-Related Muscular Dystrophy; Adam, M.P., Ardinger, H.H., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Stephens, K., Eds.; GeneReviews: Seattle, WA, USA, 1993.
- Hayashi, Y.K.; Tezak, Z.; Momoi, T.; Nonaka, I.; Garcia, C.A.; Hoffman, E.P.; Arahata, K. Massive muscle cell degeneration in the early stage of merosin-deficient congenital muscular dystrophy. Neuromuscul. Disord. 2001, 11, 350–359. [Google Scholar] [CrossRef]
- Mukasa, T.; Momoi, T.; Momoi, M.Y. Activation of caspase-3 apoptotic pathways in skeletal muscle fibers in laminin alpha2-deficient mice. Biochem. Biophys. Res. Commun. 1999, 260, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Yurchenco, P.D.; McKee, K.K.; Reinhard, J.R.; Ruegg, M.A. Laminin-deficient muscular dystrophy: Molecular pathogenesis and structural repair strategies. Matrix Biol. 2018, 71–72, 174–187. [Google Scholar] [CrossRef]
- Gawlik, K.I. At the Crossroads of Clinical and Preclinical Research for Muscular Dystrophy-Are We Closer to Effective Treatment for Patients? Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef]
- Lewis, P.; Sheehan, D.; Soares, R.; Coelho, A.V.; O’Halloran, K.D. Redox Remodeling Is Pivotal in Murine Diaphragm Muscle Adaptation to Chronic Sustained Hypoxia. Am. J. Respir. Cell Mol. Biol. 2016, 55, 12–23. [Google Scholar] [CrossRef]
- Shortt, C.M.; Fredsted, A.; Chow, H.B.; Williams, R.; Skelly, J.R.; Edge, D.; Bradford, A.; O’Halloran, K.D. Reactive oxygen species mediated diaphragm fatigue in a rat model of chronic intermittent hypoxia. Exp. Physiol. 2014, 99, 688–700. [Google Scholar] [CrossRef]
- Rodriguez, B.M.; Khouzami, L.; Decostre, V.; Varnous, S.; Pekovic-Vaughan, V.; Hutchison, C.J.; Pecker, F.; Bonne, G.; Muchir, A. N-acetyl cysteine alleviates oxidative stress and protects mice from dilated cardiomyopathy caused by mutations in nuclear A-type lamins gene. Hum. Mol. Genet. 2018, 27, 3353–3360. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, N.P.; Pham, C.; Gervasio, O.L.; Allen, D.G. N-Acetylcysteine ameliorates skeletal muscle pathophysiology in mdx mice. J. Physiol. 2008, 586, 2003–2014. [Google Scholar] [CrossRef] [PubMed]
- Pinniger, G.J.; Terrill, J.R.; Assan, E.B.; Grounds, M.D.; Arthur, P.G. Pre-clinical evaluation of N-acetylcysteine reveals side effects in the mdx mouse model of Duchenne muscular dystrophy. J. Physiol. 2017, 595, 7093–7107. [Google Scholar] [CrossRef] [PubMed]
- Terrill, J.R.; Radley-Crabb, H.G.; Grounds, M.D.; Arthur, P.G. N-Acetylcysteine treatment of dystrophic mdx mice results in protein thiol modifications and inhibition of exercise induced myofibre necrosis. Neuromuscul. Disord. 2012, 22, 427–434. [Google Scholar] [CrossRef]
- Moraes, L.H.; Bollineli, R.C.; Mizobuti, D.S.; Silveira Ldos, R.; Marques, M.J.; Minatel, E. Effect of N-acetylcysteine plus deferoxamine on oxidative stress and inflammation in dystrophic muscle cells. Redox Rep. 2015, 20, 109–115. [Google Scholar] [CrossRef]
- Matuszczak, Y.; Farid, M.; Jones, J.; Lansdowne, S.; Smith, M.A.; Taylor, A.A.; Reid, M.B. Effects of N-acetylcysteine on glutathione oxidation and fatigue during handgrip exercise. Muscle Nerve. 2005, 32, 633–638. [Google Scholar] [CrossRef]
- Reid, M.B.; Stokic, D.S.; Koch, S.M.; Khawli, F.A.; Leis, A.A. N-acetylcysteine inhibits muscle fatigue in humans. J. Clin. Invest. 1994, 94, 2468–2474. [Google Scholar] [CrossRef]
- Todd, J.J.; Lawal, T.A.; Witherspoon, J.W.; Chrismer, I.C.; Razaqyar, M.S.; Punjabi, M.; Elliott, J.S.; Tounkara, F.; Kuo, A.; Shelton, M.O.; et al. Randomized controlled trial of N-acetylcysteine therapy for RYR1-related myopathies. Neurology 2020. [Google Scholar] [CrossRef]
- Berk, M.; Malhi, G.S.; Gray, L.J.; Dean, O.M. The promise of N-acetylcysteine in neuropsychiatry. Trends Pharmacol. Sci. 2013, 34, 167–177. [Google Scholar] [CrossRef]
- Dill, C.; Prigent, H.; Behin, A.; Piemonte, F.; Bertini, E.; Orlikowski, D.; Estournet, B.; Ferreiro, A. Launching the first clinical trial in SEPN1-related myopathy: The SELNAC study. Neuromuscul. Disord. 2015, 25, 270. [Google Scholar] [CrossRef]
- Niki, E. Role of vitamin E as a lipid-soluble peroxyl radical scavenger: In vitro and in vivo evidence. Free Radic. Biol. Med. 2014, 66, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Aboul-Soud, M.A.; Al-Othman, A.M.; El-Desoky, G.E.; Al-Othman, Z.A.; Yusuf, K.; Ahmad, J.; Al-Khedhairy, A.A. Hepatoprotective effects of vitamin E/selenium against malathion-induced injuries on the antioxidant status and apoptosis-related gene expression in rats. J. Toxicol. Sci. 2011, 36, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Berneske, G.M.; Butson, A.R.; Gauld, E.N.; Levy, D. Clinical trial of high dosage vitamin E in human muscular dystrophy. Can. Med. Assoc. J. 1960, 82, 418–421. [Google Scholar] [PubMed]
- Passerieux, E.; Hayot, M.; Jaussent, A.; Carnac, G.; Gouzi, F.; Pillard, F.; Picot, M.C.; Bocker, K.; Hugon, G.; Pincemail, J.; et al. Effects of vitamin C, vitamin E, zinc gluconate, and selenomethionine supplementation on muscle function and oxidative stress biomarkers in patients with facioscapulohumeral dystrophy: A double-blind randomized controlled clinical trial. Free Radic. Biol. Med. 2015, 81, 158–169. [Google Scholar] [CrossRef]
- Erb, M.; Meinen, S.; Barzaghi, P.; Sumanovski, L.T.; Courdier-Fruh, I.; Ruegg, M.A.; Meier, T. Omigapil ameliorates the pathology of muscle dystrophy caused by laminin-alpha2 deficiency. J. Pharmacol. Exp. Ther. 2009, 331, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Girgenrath, M.; Beermann, M.L.; Vishnudas, V.K.; Homma, S.; Miller, J.B. Pathology is alleviated by doxycycline in a laminin-alpha2-null model of congenital muscular dystrophy. Ann. Neurol. 2009, 65, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Elbaz, M.; Yanay, N.; Aga-Mizrachi, S.; Brunschwig, Z.; Kassis, I.; Ettinger, K.; Barak, V.; Nevo, Y. Losartan, a therapeutic candidate in congenital muscular dystrophy: Studies in the dy(2J) /dy(2J) mouse. Ann. Neurol. 2012, 71, 699–708. [Google Scholar] [CrossRef]
- Elbaz, M.; Yanay, N.; Laban, S.; Rabie, M.; Mitrani-Rosenbaum, S.; Nevo, Y. Life or death by NFkappaB, Losartan promotes survival in dy2J/dy2J mouse of MDC1A. Cell Death Dis. 2015, 6, e1690. [Google Scholar] [CrossRef]
- Meinen, S.; Lin, S.; Ruegg, M.A. Angiotensin II type 1 receptor antagonists alleviate muscle pathology in the mouse model for laminin-alpha2-deficient congenital muscular dystrophy (MDC1A). Skelet. Muscle 2012, 2, 18. [Google Scholar] [CrossRef] [PubMed]
- Connolly, A.M.; Keeling, R.M.; Streif, E.M.; Pestronk, A.; Mehta, S. Complement 3 deficiency and oral prednisolone improve strength and prolong survival of laminin alpha2-deficient mice. J. Neuroimmunol. 2002, 127, 80–87. [Google Scholar] [CrossRef]
- Hayes, A.; Williams, D.A. Examining potential drug therapies for muscular dystrophy utilising the dy/dy mouse: I. Clenbuterol. J. Neurol. Sci. 1998, 157, 122–128. [Google Scholar] [CrossRef]
- Accorsi, A.; Kumar, A.; Rhee, Y.; Miller, A.; Girgenrath, M. IGF-1/GH axis enhances losartan treatment in Lama2-related muscular dystrophy. Hum. Mol. Genet. 2016, 25, 4624–4634. [Google Scholar] [CrossRef] [PubMed]
- Meinen, S.; Lin, S.; Thurnherr, R.; Erb, M.; Meier, T.; Ruegg, M.A. Apoptosis inhibitors and mini-agrin have additive benefits in congenital muscular dystrophy mice. EMBO Mol. Med. 2011, 3, 465–479. [Google Scholar] [CrossRef]
- Yamauchi, J.; Kumar, A.; Duarte, L.; Mehuron, T.; Girgenrath, M. Triggering regeneration and tackling apoptosis: A combinatorial approach to treating congenital muscular dystrophy type 1 A. Hum. Mol. Genet. 2013, 22, 4306–4317. [Google Scholar] [CrossRef] [PubMed]
- Grandjean, E.M.; Berthet, P.; Ruffmann, R.; Leuenberger, P. Efficacy of oral long-term N-acetylcysteine in chronic bronchopulmonary disease: A meta-analysis of published double-blind, placebo-controlled clinical trials. Clin. Ther. 2000, 22, 209–221. [Google Scholar] [CrossRef]
- Bailey, B.; McGuigan, M.A. Management of anaphylactoid reactions to intravenous N-acetylcysteine. Ann. Emerg. Med. 1998, 31, 710–715. [Google Scholar] [CrossRef]
- Arakawa, M.; Ito, Y. N-acetylcysteine and neurodegenerative diseases: Basic and clinical pharmacology. Cerebellum 2007, 6, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Tardiolo, G.; Bramanti, P.; Mazzon, E. Overview on the Effects of N-Acetylcysteine in Neurodegenerative Diseases. Molecules 2018, 23. [Google Scholar] [CrossRef]
- Hardan, A.Y.; Fung, L.K.; Libove, R.A.; Obukhanych, T.V.; Nair, S.; Herzenberg, L.A.; Frazier, T.W.; Tirouvanziam, R. A randomized controlled pilot trial of oral N-acetylcysteine in children with autism. Biol. Psychiatry 2012, 71, 956–961. [Google Scholar] [CrossRef]
- Hagos, F.T.; Empey, P.E.; Wang, P.; Ma, X.; Poloyac, S.M.; Bayir, H.; Kochanek, P.M.; Bell, M.J.; Clark, R.S.B. Exploratory Application of Neuropharmacometabolomics in Severe Childhood Traumatic Brain Injury. Crit. Care Med. 2018, 46, 1471–1479. [Google Scholar] [CrossRef] [PubMed]
- Nobili, V.; Manco, M.; Devito, R.; Ciampalini, P.; Piemonte, F.; Marcellini, M. Effect of vitamin E on aminotransferase levels and insulin resistance in children with non-alcoholic fatty liver disease. Aliment. Pharmacol. Ther. 2006, 24, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Feranchak, A.P.; Sontag, M.K.; Wagener, J.S.; Hammond, K.B.; Accurso, F.J.; Sokol, R.J. Prospective, long-term study of fat-soluble vitamin status in children with cystic fibrosis identified by newborn screen. J. Pediatr. 1999, 135, 601–610. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Individual | Muscle | Age at Biopsy | LAMA2 Mutation | Laminin α2- Deficiency | DHE Positive Area (%) |
|---|---|---|---|---|---|
| Patients | Quadriceps | 4 years | Unknown | Partial | 4.94 |
| Unknown | 17 years | Unknown | Complete | 5.27 | |
| Unknown | 22 days | Homo c.2208 + 2T>C | Complete | 10.55 | |
| Radial forearm | 29 years | Homo c.2230C>T | Partial | 4.23 | |
| Deltoid | 2 years | Unknown | Complete | 6.42 | |
| Deltoid | 3.5 years | Unknown | Complete | 10.27 | |
| Controls | Radial forearm | 33 years | - | - | 3.56 |
| Quadriceps | 17 years | - | - | 3.10 | |
| Vastus | 2 months | - | - | 2.34 | |
| Unknown | 2.5 years | - | - | 1.88 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
M. Harandi, V.; Moreira Soares Oliveira, B.; Allamand, V.; Friberg, A.; Fontes-Oliveira, C.C.; Durbeej, M. Antioxidants Reduce Muscular Dystrophy in the dy2J/dy2J Mouse Model of Laminin α2 Chain-Deficient Muscular Dystrophy. Antioxidants 2020, 9, 244. https://doi.org/10.3390/antiox9030244
M. Harandi V, Moreira Soares Oliveira B, Allamand V, Friberg A, Fontes-Oliveira CC, Durbeej M. Antioxidants Reduce Muscular Dystrophy in the dy2J/dy2J Mouse Model of Laminin α2 Chain-Deficient Muscular Dystrophy. Antioxidants. 2020; 9(3):244. https://doi.org/10.3390/antiox9030244
Chicago/Turabian StyleM. Harandi, Vahid, Bernardo Moreira Soares Oliveira, Valérie Allamand, Ariana Friberg, Cibely C. Fontes-Oliveira, and Madeleine Durbeej. 2020. "Antioxidants Reduce Muscular Dystrophy in the dy2J/dy2J Mouse Model of Laminin α2 Chain-Deficient Muscular Dystrophy" Antioxidants 9, no. 3: 244. https://doi.org/10.3390/antiox9030244
APA StyleM. Harandi, V., Moreira Soares Oliveira, B., Allamand, V., Friberg, A., Fontes-Oliveira, C. C., & Durbeej, M. (2020). Antioxidants Reduce Muscular Dystrophy in the dy2J/dy2J Mouse Model of Laminin α2 Chain-Deficient Muscular Dystrophy. Antioxidants, 9(3), 244. https://doi.org/10.3390/antiox9030244