Structure-Dependent Effects of Bisphosphonates on Inflammatory Responses in Cultured Neonatal Mouse Calvaria

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Calvaria Culture
2.2. Effects of NBPs and Non-NBPs on PGE2 and NO Production in Calvaria
2.3. Reverse Transcription-PCR (RT-PCR) Analyses of Cultured Calvaria
2.4. Fluorescent Staining of Calvaria and Laser Scanning Confocal Microscopy
2.5. Western Blot Analysis in Cultured Calvaria
2.6. Effects of Non-NBPs on the Generation of Superoxide Anions in the Xanthine-Xanthine Oxidase System
2.7. Statistical Analysis
3. Results
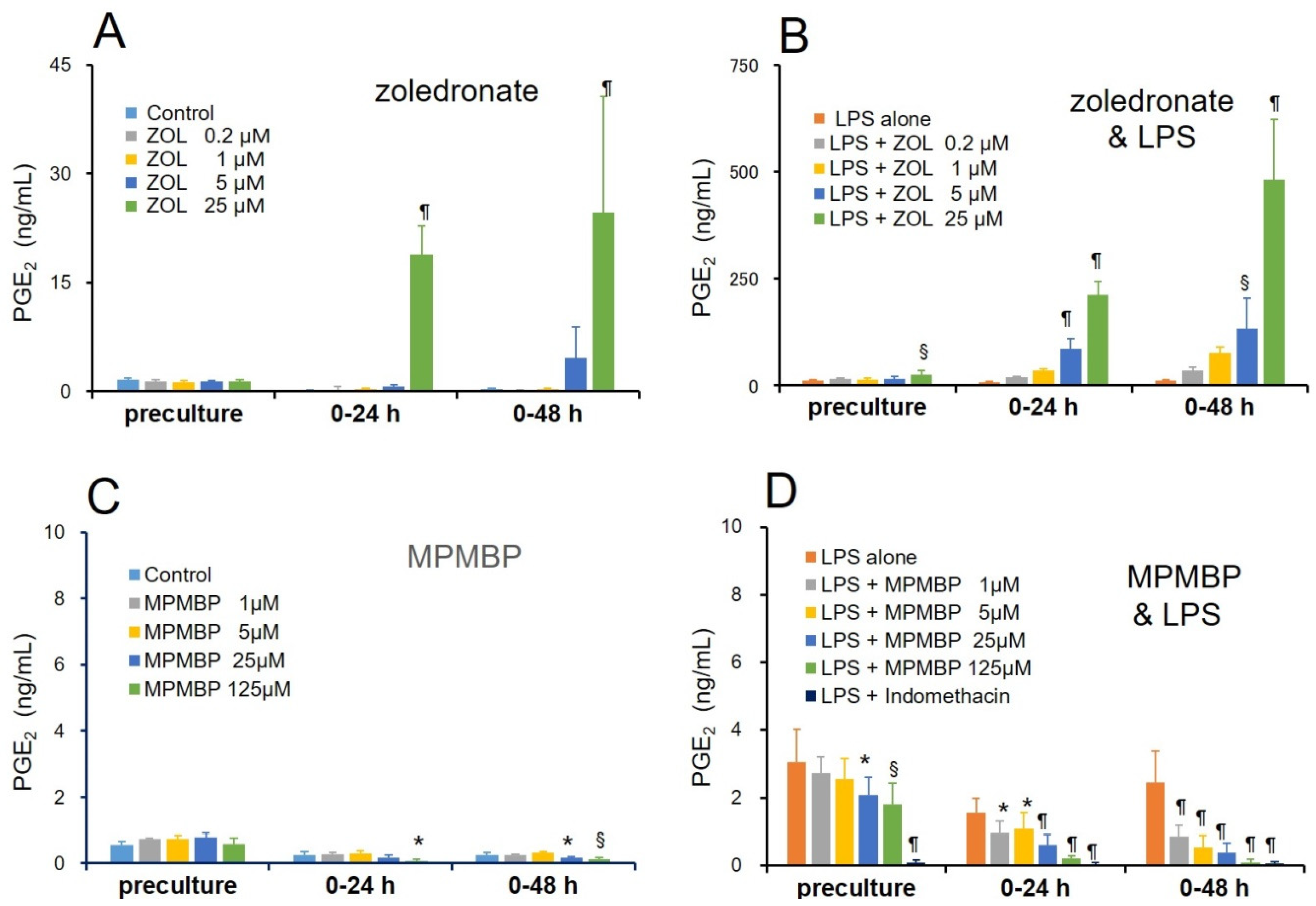
3.1. Effects of NBPs and Non-NBPs on PGE2 Production in Cultured Calvaria
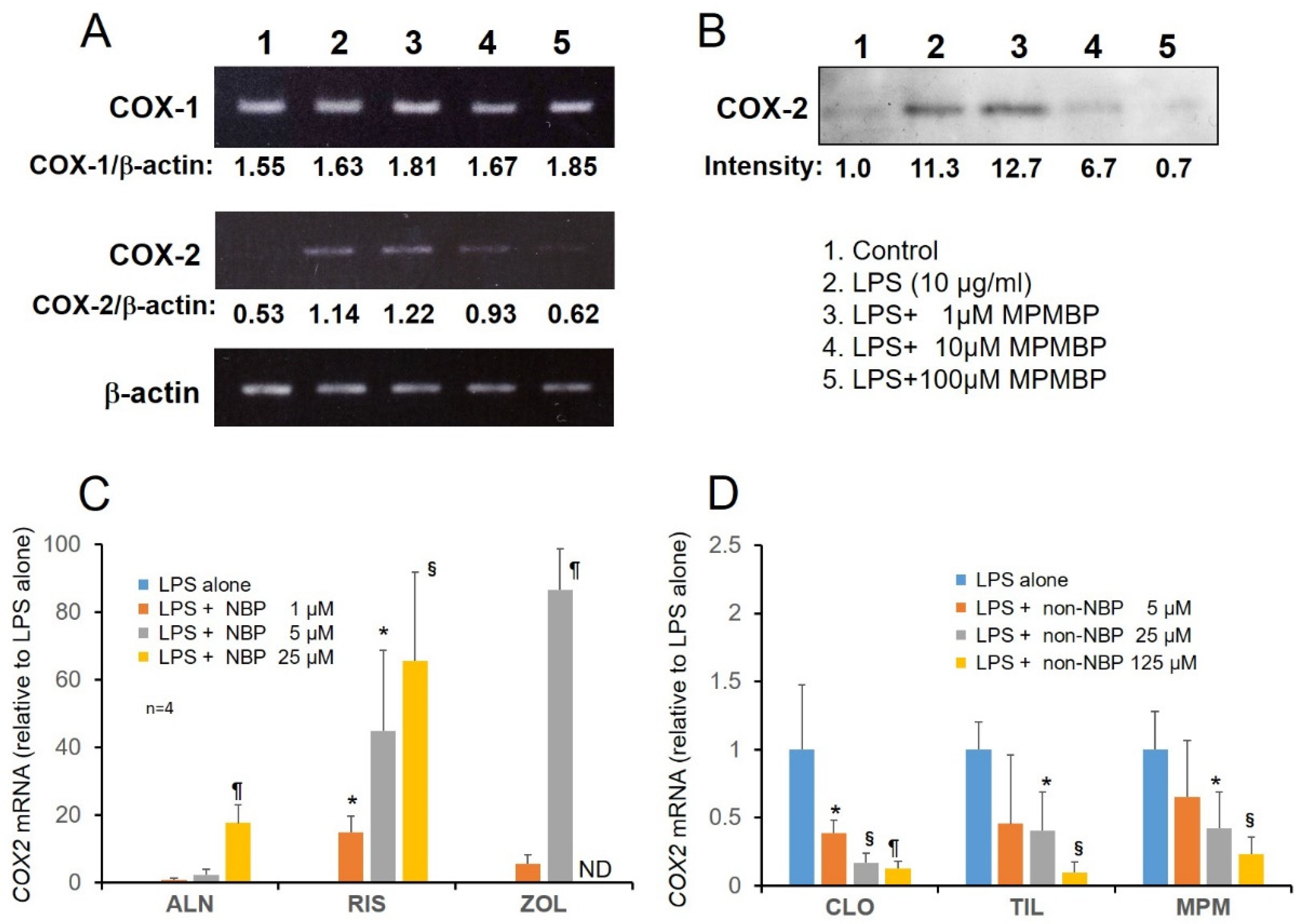
3.2. Effects of NBPs and Non-NBPs on COX-2 Expression in Cultured Calvaria
3.3. Effects of NBPs and Non-NBPs on the Production of NO and Expression of iNOS mRNA in Cultured Calvaria
3.4. Signaling Pathway through which MPMBP Inhibits the Production of Inflammatory Mediators
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Russell, R.G. Bisphosphonates: The first 40 years. Bone 2011, 49, 2–19. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.J.; Watts, D.J.; Russell, R.G. Overview of bisphosphonates. Cancer 1997, 80, 1652–1660. [Google Scholar] [CrossRef] [Green Version]
- Fleisch, H. Bisphosphonates in Bone Disease. From the Laboratory to the Patient, 4th ed.; Academic Press: San Diego, CA, USA, 2000. [Google Scholar]
- van Beek, E.R.; Cohen, L.H.; Leroy, I.M.; Ebetino, F.M.; Löwik, C.W.G.; Papapoulos, S.E. Differentiating the mechanisms of antiresorptive action of nitrogen containing bisphosphonates. Bone 2003, 33, 805–811. [Google Scholar] [CrossRef]
- Rogers, M.J.; Gordon, S.; Benford, H.L.; Coxon, F.P.; Luckman, S.P.; Monkkonen, J.; Frith, J.C. Cellular and molecular mechanisms of action of bisphosphonates. Cancer 2000, 88, 2961–2978. [Google Scholar] [CrossRef]
- Khosla, S.; Burr, D.; Cauley, J.; Dempster, D.W.; Ebeling, P.R.; Felsenberg, D.; Gagel, R.F.; Gilsanz, V.; Guise, T.; Koka, S.; et al. Bisphosphonate-associated osteonecrosis of the jaw: Report of a task force of the American society for bone and mineral research. J. Bone Miner. Res. 2007, 22, 1479–1491. [Google Scholar] [CrossRef] [Green Version]
- Durie, B.G.M.; Katz, M.; Crowley, J. Osteonecrosis of the jaw and bisphosphonates. N. Engl. J. Med. 2005, 353, 99–102. [Google Scholar]
- Bamias, A.; Kastritis, E.; Bamia, C.; Moulopoulos, L.A.; Melakopoulos, I.; Bozas, G.; Koutsoukou, V.; Gika, D.; Anagnostopoulos, A.; Papadimitriou, C.; et al. Osteonecrosis of the jaw in cancer after treatment with bisphosphonates: Incidence and risk factors. J. Clin. Oncol. 2006, 34, 8580–8585. [Google Scholar] [CrossRef]
- Hoff, A.O.; Toth, B.B.; Altundag, K.; Johnson, M.M.; Warneke, C.L.; Hu, M.; Nooka, A.; Sayegh, G.; Guarneri, V.; Desrouleaux, K.; et al. Frequency and risk factors associated with osteonecrosis of the jaw in cancer patients treated with intravenous bisphosphonates. J. Bone Miner. Res. 2008, 23, 826–836. [Google Scholar] [CrossRef]
- Hewitt, R.E.; Lissina, A.; Green, A.E.; Slay, E.S.; Price, D.A.; Sewell, A.K. The bisphosphonate acute phase response: Rapid and copious production of proinflammatory cytokines by peripheral blood γφ T cells in response to aminobisphosphonates is inhibited by statins. Clin. Exp. Immunol. 2005, 139, 101–111. [Google Scholar] [CrossRef]
- Kumar, V.; Shahi, A.K. Nitrogen containing bisphosphonates associated osteonecrosis of the jaws: A review for past 10 year literature. Dent. Res. J. 2014, 11, 147–153. [Google Scholar]
- Takizawa, A.; Chiba, M.; Ota, T.; Yasuda, M.; Suzuki, K.; Kanemitsu, T.; Itoh, T.; Shinoda, H.; Igarashi, K. The novel bisphosphonate disodium dihydrogen-4-[(methylthio) phenylthio] methane bisphosphonate increases bone mass in post-ovariectomy rats. J. Pharmacol. Sci. 2016, 131, 37–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanahashi, M.; Funaba, Y.; Tateishi, A.; Kawabe, N.; Nakadate-Matsushita, T. TRK-530 inhibits accumulation of superoxide anions derived from human polymorphonuclear leukocytes and bone resorption induced by activated osteoclasts. Pharmacology 1998, 56, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Takaoka, Y.; Nagai, H.; Mori, H.; Tanahashi, M. The effect of TRK-530 on experimental arthritis in mice. Biol. Pharm. Bull. 1997, 20, 1147–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanahashi, M.; Koike, J.; Kawabe, N.; Nakadate-Matsushita, T. Inhibitory effect of TRK-530 on inflammatory cytokines in bone marrow of rats with adjuvant arthritis. Pharmacology 1998, 56, 237–241. [Google Scholar] [CrossRef]
- Tanahashi, M.; Funaba, Y.; Itoh, M.; Kawabe, N.; Nakadate-Matsushita, T. Inhibitory effects of TRK-530 on rat adjuvant arthritis. Pharmacology 1998, 56, 242–251. [Google Scholar] [CrossRef]
- Attia, A.M.M.; Ibrahim, F.A.A.; Abd El-Latif, N.A.; Aziz, S.W.; Elwan, A.M.; Aziz, A.; Elgendy, A.; Elgengehy, F.T. Role of reactive nitrogen species and antioxidant defense systems in the pathogenesis of rheumatoid arthritis. Wulfenia 2015, 22, 120–135. [Google Scholar]
- Shinoda, H.; Takeyama, S.; Suzuki, K.; Murakami, S.; Yamada, S. Pharmacological topics of bone metabolism: A novel bisphosphonate for the treatment of periodontitis. J. Pharmacol. Sci. 2008, 106, 555–558. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Takeshita, F.; Yamamoto, K.; Yamada, S.; Shinoda, H.; Ochiya, T. In vivo imaging of osteoclast precursor recruitment to the inflammatory site where extensive bone destruction occurs. In Proceedings of the ASBMR 2010 Annual Meeting, Toronto, ON, Canada, 15–19 October 2010. MO0263. [Google Scholar]
- Nathan, C. Points of control in inflammation. Nature 2002, 420, 846–852. [Google Scholar] [CrossRef]
- Lee, K.W.; Jeong, S.Y.; Park, H.J.; Jung, H.J.; Cho, Y.W.; Yun, K.; Lee, K.T. Inhibition of LPS-induced NO and PGE2 production by asiatic acid via NF-kappa B inactivation in RAW 264.7 macrophages: Possible involvement of the IKK and MAPK pathways. Int. Immunopharmacol. 2008, 8, 431–441. [Google Scholar]
- Suzuki, K.; Takeyama, S.; Kikuchi, T.; Yamada, S.; Sodek, J.; Shinoda, H. Osteoclast responses to lipopolysaccharide, parathyroid hormone and bisphosphonates in neonatal murine calvaria analyzed by laser scanning confocal microscopy. J. Histochem. Cytochem. 2005, 53, 1525–1537. [Google Scholar] [CrossRef] [Green Version]
- Pikarsky, E.; Porat, R.M.; Stein, I.; Abramovitch, R.; Amit, S.; Kasem, S.; Gutkovich-Pyest, E.; Urieli-Shoval, S.; Galun, E.; Ben-Neriah, Y. NF-kappaB functions as a tumour promoter in inflammation-associated cancer. Nature 2004, 431, 461–466. [Google Scholar] [CrossRef]
- Hogg, N.; Darley-Usmar, V.M.; Wilson, M.T.; Moncada, S. Production of hydroxyl radicals from the simultaneous generation of superoxide and nitric oxide. Biochem. J. 1992, 281, 419–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosfield, D.J.; Zhang, Y.; Dougan, D.R.; Broun, A.; Tari, L.W.; Swanson, R.V.; Finn, J. Structural basis for bisphosphonate-mediated inhibition of isoprenoid biosynthesis. J. Biol. Chem. 2003, 279, 8526–8529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyaura, C.; Inada, M.; Matsumoto, C.; Ohshiba, T.; Uozumi, N.; Shimizu, T.; Ito, A. An essential role of cytosolic phospholipase A2alpha in prostaglandin E2-mediated bone resorption associated with inflammation. J. Exp. Med. 2003, 197, 1303–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wadleigh, D.J.; Herschman, H.R. Transcriptional regulation of the cyclooxygenase-2 gene by diverse ligands in murine osteoblasts. Biochem. Biophys. Res. Commun. 1999, 264, 865–870. [Google Scholar] [CrossRef]
- Watson, W.H.; Zhao, Y.; Chawla, R.K. S-adenosylmethionine attenuates the lipopolysaccharide-induced expression of the gene for the tumour necrosis factor-α. Biochem. J. 1999, 342, 21–25. [Google Scholar] [CrossRef]
- Kubes, P.; McCafferty, D.M. Nitric oxide and intestinal inflammation. Am. J. Med. 2000, 109, 150–158. [Google Scholar] [CrossRef]
- Gu, Q.; Yang, H.; Shi, Q. Macrophages and bone inflammation. J. Orthop. Translat. 2017, 10, 86–93. [Google Scholar] [CrossRef]
- Tamai, R.; Suzuki, K.; Mashima, I.; Kiyoura, Y. MPMBP down-regulates Toll-like receptor (TLR) 2 ligand-induced proinflammatory cytokine production by inhibiting NF-κB but not AP-1 activation. Int. Immunopharmacol. 2020, 79, 106085. [Google Scholar] [CrossRef]
- Ischiropoulos, H. Biological tyrosine nitration: A pathophysiological function of nitric oxide and reactive oxygen species. Arch. Biochem. Biophys. 1998, 356, 1–11. [Google Scholar] [CrossRef]
- Gochman, E.; Mahajna, J.; Reznick, A.Z. NF-κB activation by peroxynitrite through IκBα-dependent phosphorylation versus nitration in colon cancer cells. Anticancer Res. 2011, 31, 1607–1617. [Google Scholar] [PubMed]
- Janssen-Heininger, Y.M.; Macara, I.; Mossman, B.T. Cooperativity between oxidants and tumor necrosis factor in the activation of nuclear factor (NF)-kappaB: Requirement of Ras/mitogen-activated protein kinases in the activation of NF-kappaB by oxidants. Am. J. Respir. Cell Mol. Biol. 1999, 20, 942–952. [Google Scholar] [CrossRef] [PubMed]
- Yakovlev, V.A.; Barani, I.J.; Rabender, C.S.; Black, S.M.; Leach, J.K.; Graves, P.R.; Kellogg, G.E.; Mikkelsen, R.B. Tyrosine nitration of IκBα: A novel mechanism for NF-κB activation. Biochemistry 2007, 46, 11671–11683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaoka, M.; Maeda, T.; Chatani, M.; Handa, K.; Yamakawa, T.; Kiyohara, S.; Negishi-Koga, T.; Kato, Y.; Takami, M.; Niida, S.; et al. A delphinidin-enriched maqui berry extract improves bone metabolism and protects against bone loss in osteopenic mouse models. Antioxidants 2019, 8, 386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabó, C.; Módis, K. Pathophysiological roles of peroxynitrite in circulatory shock. Shock 2010, 34, 4–14. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, S.; Gorren, A.C.F.; Schmidt, K.; Werner, E.R.; Hansert, B.; Bohle, D.S.; Mayer, B. Metabolic fate of peroxynitrite in aqueous solution: Reaction with nitric oxide and pH-dependent decomposition to nitrite and oxygen in a 2:1 stoichiometry. J. Biol. Chem. 1997, 272, 3465–3470. [Google Scholar] [CrossRef] [Green Version]
- Estévez, A.G.; Spear, N.; Manuel, S.M.; Radi, R.; Henderson, C.E.; Barbeito, L.; Beckman, J.S. Nitric oxide and superoxide contribute to motor neuron apoptosis induced by trophic factor deprivation. J. Neurosci. 1998, 18, 923–931. [Google Scholar] [CrossRef]
- Marla, S.S.; Lee, J.; Groves, J.T. Peroxynitrite rapidly permeates phospholipid membranes. Proc. Natl. Acad. Sci. USA 1997, 94, 14243–14248. [Google Scholar] [CrossRef] [Green Version]
- Mazel, P.; Henderson, L.F.; Axelrod, J. S-demethylation by microsomal enzymes. J. Pharmacol. Exp. Ther. 1964, 143, 1–6. [Google Scholar]
- Larsen, G.L.; Bakke, J.E.; Feil, V.J.; Huwe, J.K. In vitro metabolism of the methylthio group of 2-methylthiobenzothiazole by rat liver. Xenobiotica 1988, 18, 313–322. [Google Scholar] [CrossRef]
- Liu, S.F.; Ye, X.; Malik, A.B. Inhibition of NF-κB activation by pyrrolidine dithiocarbamate prevents in vivo expression of proinflammatory genes. Circulation 1999, 100, 1330–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.F.; Ye, X.; Malik, A.B. In vivo inhibition of nuclear factor-kappa B activation prevents inducible nitric oxide synthase expression and systemic hypotension in a rat model of septic shock. J. Immunol. 1997, 159, 3976–3983. [Google Scholar] [PubMed]
- Kalyan, S.; Huebbe, P.; Esatbeyoglu, T.; Niklowitz, P.; Côté, H.C.F.; Rimbach, G.; Kabelitz, D. Nitrogen-bisphosphonate therapy is linked to compromised coenzyme Q10 and vitamin E status in postmenopausal women. J. Clin. Endocrinol. Metab. 2014, 99, 1307–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Littarru, G.P.; Nakamura, R.; Ho, L.; Folkers, K.; Kuzell, W.C. Deficiency of coenzyme Q 10 in gingival tissue from patients with periodontal disease. Proc. Natl. Acad. Sci. USA 1971, 68, 2332–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, S.; Sunitha, J.; Hans, M. Role of coenzyme Q (10) as an antioxidant and bioenergizer in periodontal diseases. Indian J. Pharmacol. 2010, 42, 334–337. [Google Scholar] [CrossRef] [Green Version]
- Dallner, G.; Sindelar, P.J. Regulation of ubiquinone metabolism. Free Radic. Biol. Med. 2000, 29, 285–294. [Google Scholar] [CrossRef]
- Nawarskas, J.J. HMG-CoA Reductase inhibitors and coenzyme Q10. Cardiol. Rev. 2005, 13, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Elmberger, P.G.; Kalen, A.; Appelkvist, E.L.; Dallner, G. In vitro and in vivo synthesis of dolichol and other main mevalonate products in various organs of the rat. Eur. J. Biochem. 1987, 168, 1–11. [Google Scholar] [CrossRef]
- Marx, R.E. Pamidronate (Aredia) and zoledronate (Zometa) induced avascular necrosis of the jaws: A growing epidemic. J. Oral Maxillofac. Surg. 2003, 9, 1115–1117. [Google Scholar] [CrossRef]
- Lesclous, P.; Abi, N.S.; Carrel, J.P.; Baroukh, B.; Lombardi, T.; Willi, J.P.; Rizzoli, R.; Saffar, J.L.; Samson, J. Bisphosphonate-associated osteonecrosis of the jaw: A key role of inflammation? Bone 2009, 45, 843–852. [Google Scholar] [CrossRef] [Green Version]
- Iannitti, T.; Rosini, S.; Lodi, D.; Frediani, B.; Rottigni, V.; Palmieri, B. Bisphosphonates: Focus on inflammation and bone loss. Am. J. Ther. 2012, 19, 228–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khosla, S.; Bilezikian, J.P.; Dempster, D.W.; Lewiecki, M.E.; Miller, P.D.; Neer, R.M.; Recker, R.R.; Shane, E.; Shoback, D.; Potts, J.T. Benefits and risks of bisphosphonate therapy for osteoporosis. J. Clin. Endocrinol. Metab. 2012, 97, 2272–2282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cremers, S.; Papapoulos, S. Pharmacology of bisphosphonates. Bone 2011, 49, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Rodan, G.A.; Martin, T.J. Therapeutic approaches to bone diseases. Science 2000, 289, 1508–1514. [Google Scholar] [CrossRef]
- Sandhu, J.K.; Robertson, S.; Birnboim, H.C.; Goldstein, R. Distribution of protein nitrotyrosine in synovial tissues of patients with rheumatoid arthritis and osteoarthritis. J. Rheumatol. 2003, 30, 1173–1181. [Google Scholar]
- Hokugo, A.; Kanayama, K.; Sun, S.; Morinaga, K.; Sun, Y.; Wu, Q.; Sasaki, H.; Okawa, H.; Evans, C.; Ebetino, F.H.; et al. Rescue bisphosphonate treatment of alveolar bone improves extraction socket healing and reduces osteonecrosis in zoledronate-treated mice. Bone 2019, 123, 115–128. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequences (5′-3′) | Product Size (bp) | Accession Number | |
|---|---|---|---|---|
| Forward | Reverse | |||
| COX-1 | acc tgc ccc tat gtt tcc tt | cct tcc aag tgc tgg gat ta | 421 | NM_008969.4 |
| COX-2 | aag aca gat cat aag cga gga c | gcc agt gat aga gtg tgt tga a | 532 | NM_011198.4 |
| iNOS | ctg ttc tca gcc caa caa tac a | gct ggt agg ttc ctg ttg ttt c | 423 | NM_010927.4 |
| eNOS | agg aag gag tct agc aac aca gac | gtg agt cag ccc tgg tag taa ttg | 369 | NM_008713.4 |
| nNOS | gct gtc cta tac agc ttc cag agt | caa agt tgt ctc tga ggt ctg gtc | 700 | NM_008712.3 |
| Actb | tgt tac caa ctg gga cga ca | atg cca cag gat tcc ata cc | 597 | NM_007393.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, K.; Takeyama, S.; Murakami, S.; Nagaoka, M.; Chiba, M.; Igarashi, K.; Shinoda, H. Structure-Dependent Effects of Bisphosphonates on Inflammatory Responses in Cultured Neonatal Mouse Calvaria. Antioxidants 2020, 9, 503. https://doi.org/10.3390/antiox9060503
Suzuki K, Takeyama S, Murakami S, Nagaoka M, Chiba M, Igarashi K, Shinoda H. Structure-Dependent Effects of Bisphosphonates on Inflammatory Responses in Cultured Neonatal Mouse Calvaria. Antioxidants. 2020; 9(6):503. https://doi.org/10.3390/antiox9060503
Chicago/Turabian StyleSuzuki, Keiko, Sadaaki Takeyama, Shinobu Murakami, Masahiro Nagaoka, Mirei Chiba, Kaoru Igarashi, and Hisashi Shinoda. 2020. "Structure-Dependent Effects of Bisphosphonates on Inflammatory Responses in Cultured Neonatal Mouse Calvaria" Antioxidants 9, no. 6: 503. https://doi.org/10.3390/antiox9060503