
BCG Vaccination-Associated Lower HbA1c and Increased CD25 Expression on CD8+ T Cells in Patients with Type 1 Diabetes in Ghana

 , ,
, ,  ,
,  , , , , , , , , and add
Show full author list
, , , , , , , , and add
Show full author list
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Cohort Characterization
2.2. BCG Vaccination
2.3. Serum Measurements
2.3.1. Random C-Peptide Measurement
2.3.2. T1D Autoantibodies
2.3.3. Analysis of Targeted Metabolites by Liquid Chromatography–Tandem Mass Spectrometry (LC-MS/MS)
2.4. T Cell Phenotyping by Flow Cytometry
2.5. Statistical Analysis
3. Results
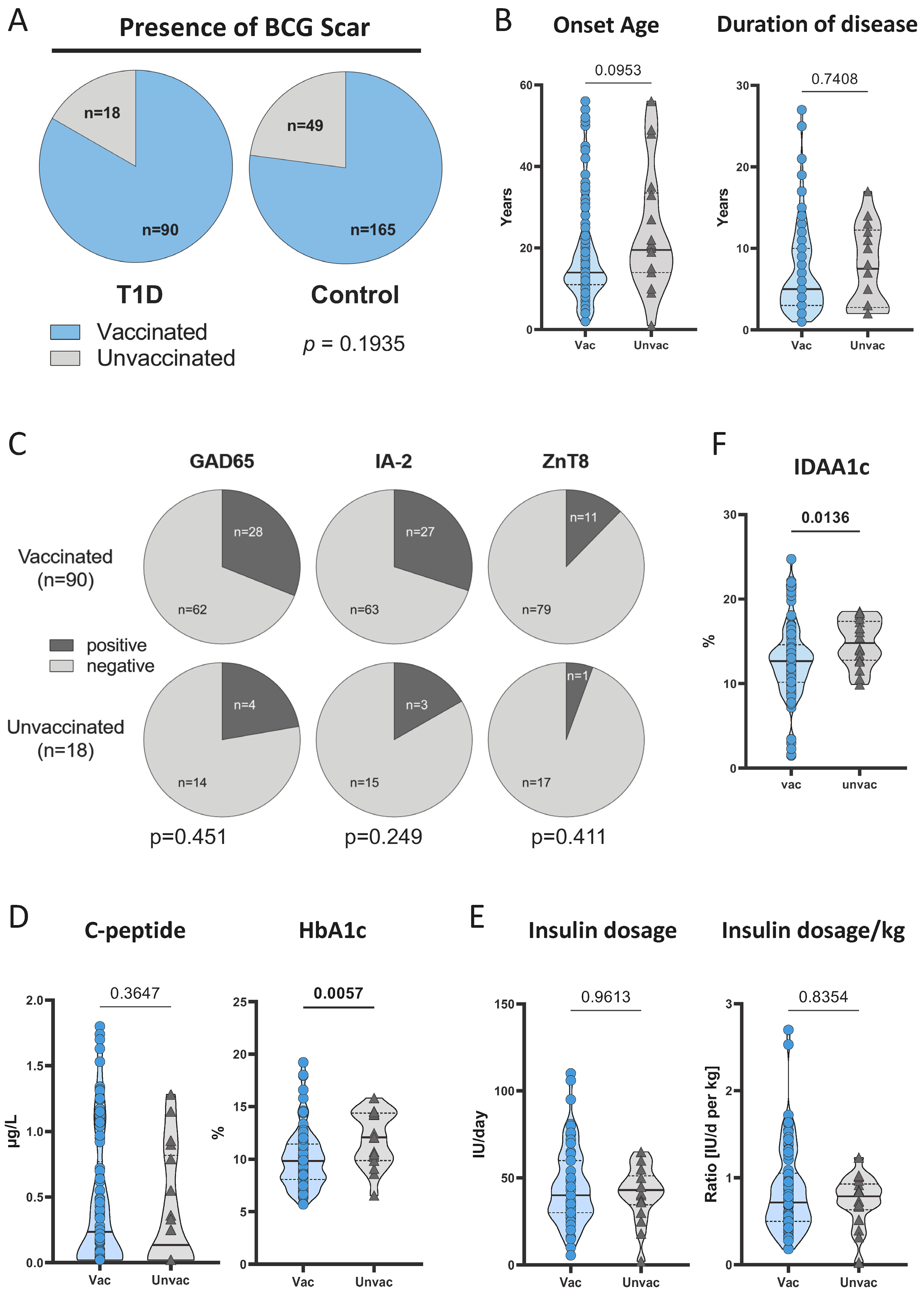
3.1. Study Characteristics and BCG Vaccination Rates
3.2. BCG-Vaccinated T1D Patients Had Lower HbA1c Compared to Unvaccinated Patients
3.3. CD25 Expression on CD8+ T Cells Is Higher in BCG-Vaccinated T1D Patients

3.4. CD25 Expression on CD8+ T Cells Correlates with HbA1c in T1D Patients
3.5. Glucose Metabolites Differentiate BCG-Vaccinated Patients from Unvaccinated Patients
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glynn, J.R.; Dube, A.; Fielding, K.; Crampin, A.C.; Karonga Prevention Trial, G.; Kanjala, C.; Fine, P.E.M. The effect of BCG revaccination on all-cause mortality beyond infancy: 30-year follow-up of a population-based, double-blind, randomised placebo-controlled trial in Malawi. Lancet Infect. Dis. 2021, 21, 1590–1597. [Google Scholar] [CrossRef] [PubMed]
- Welaga, P.; Debpuur, C.; Aaby, P.; Hodgson, A.; Azongo, D.K.; Benn, C.S.; Oduro, A.R. Is the decline in neonatal mortality in northern Ghana, 1996–2012, associated with the decline in the age of BCG vaccination? An ecological study. BMJ Open 2018, 8, e023752. [Google Scholar] [CrossRef] [PubMed]
- Adesanya, O.A.; Uche-Orji, C.I.; Adedeji, Y.A.; Joshua, J.I.; Adesola, A.A.; Chukwudike, C.J. Bacillus Calmette-Guerin (BCG): The adroit vaccine. AIMS Microbiol. 2021, 7, 96–113. [Google Scholar] [CrossRef]
- Fritschi, N.; Curtis, N.; Ritz, N. Bacille Calmette Guerin (BCG) and new TB vaccines: Specific, cross-mycobacterial and off-target effects. Paediatr. Respir. Rev. 2020, 36, 57–64. [Google Scholar] [CrossRef]
- Klinger, D.; Blass, I.; Rappoport, N.; Linial, M. Significantly Improved COVID-19 Outcomes in Countries with Higher BCG Vaccination Coverage: A Multivariable Analysis. Vaccines 2020, 8, 378. [Google Scholar] [CrossRef]
- Gong, W.; An, H.; Wang, J.; Cheng, P.; Qi, Y. The Natural Effect of BCG Vaccination on COVID-19: The Debate Continues. Front. Immunol. 2022, 13, 953228. [Google Scholar] [CrossRef]
- Pittet, L.F.; Messina, N.L.; Orsini, F.; Moore, C.L.; Abruzzo, V.; Barry, S.; Bonnici, R.; Bonten, M.; Campbell, J.; Croda, J.; et al. Randomized Trial of BCG Vaccine to Protect against Covid-19 in Health Care Workers. N. Engl. J. Med. 2023, 388, 1582–1596. [Google Scholar] [CrossRef]
- Harada, M.; Kishimoto, Y.; Makino, S. Prevention of overt diabetes and insulitis in NOD mice by a single BCG vaccination. Diabetes Res. Clin. Pract. 1990, 8, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Huppmann, M.; Baumgarten, A.; Ziegler, A.G.; Bonifacio, E. Neonatal Bacille Calmette-Guerin vaccination and type 1 diabetes. Diabetes Care 2005, 28, 1204–1206. [Google Scholar] [CrossRef]
- Rousseau, M.C.; El-Zein, M.; Conus, F.; Legault, L.; Parent, M.E. Bacillus Calmette-Guerin (BCG) Vaccination in Infancy and Risk of Childhood Diabetes. Paediatr. Perinat. Epidemiol. 2016, 30, 141–148. [Google Scholar] [CrossRef]
- Kuhtreiber, W.M.; Tran, L.; Kim, T.; Dybala, M.; Nguyen, B.; Plager, S.; Huang, D.; Janes, S.; Defusco, A.; Baum, D.; et al. Long-term reduction in hyperglycemia in advanced type 1 diabetes: The value of induced aerobic glycolysis with BCG vaccinations. NPJ Vaccines 2018, 3, 23. [Google Scholar] [CrossRef] [PubMed]
- Kuhtreiber, W.M.; Faustman, D.L. BCG Therapy for Type 1 Diabetes: Restoration of Balanced Immunity and Metabolism. Trends Endocrinol. Metab. 2019, 30, 80–92. [Google Scholar] [CrossRef]
- Keefe, R.C.; Takahashi, H.; Tran, L.; Nelson, K.; Ng, N.; Kuhtreiber, W.M.; Faustman, D.L. BCG therapy is associated with long-term, durable induction of Treg signature genes by epigenetic modulation. Sci. Rep. 2021, 11, 14933. [Google Scholar] [CrossRef] [PubMed]
- Pozzilli, P. BCG vaccine in insulin-dependent diabetes mellitus. Lancet 1997, 349, 1520–1521. [Google Scholar] [CrossRef] [PubMed]
- Allen, H.F.; Klingensmith, G.J.; Jensen, P.; Simoes, E.; Hayward, A.; Chase, H.P. Effect of Bacillus Calmette-Guerin vaccination on new-onset type 1 diabetes. A randomized clinical study. Diabetes Care 1999, 22, 1703–1707. [Google Scholar] [CrossRef] [PubMed]
- Baguune, B.; Ndago, J.A.; Adokiya, M.N. Immunization dropout rate and data quality among children 12–23 months of age in Ghana. Arch. Public Health 2017, 75, 18. [Google Scholar] [CrossRef] [PubMed]
- Sarfo-Kantanka, O.; Asamoah-Boaheng, M.; Arthur, J.; Agyei, M.; Barnes, N.A.; Tenkorang, E.Y.; Midodzi, W. Trends in type 1 diabetes diagnosis in Ghana. Int. Health 2022, 14, 442–446. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association Professional Practice Committee; El Sayed, N.A.; Aleppo, G.; Bannuru, R.R.; Bruemmer, D.; Collins, B.S.; Ekhlaspour, L.; Gaglia, J.L.; Hilliard, M.E.; Johnson, E.L.; et al. 2. Diagnosis and Classification of Diabetes: Standards of Care in Diabetes-2024. Diabetes Care 2024, 47, S20–S42. [Google Scholar] [CrossRef]
- World Health Organization. BCG vaccine: WHO position paper, February 2018—Recommendations. Vaccine 2018, 36, 3408–3410. [Google Scholar] [CrossRef]
- Phillips, R.O.; Phanzu, D.M.; Beissner, M.; Badziklou, K.; Luzolo, E.K.; Sarfo, F.S.; Halatoko, W.A.; Amoako, Y.; Frimpong, M.; Kabiru, A.M.; et al. Effectiveness of routine BCG vaccination on buruli ulcer disease: A case-control study in the Democratic Republic of Congo, Ghana and Togo. PLoS Negl. Trop. Dis. 2015, 9, e3457. [Google Scholar] [CrossRef]
- Birk, N.M.; Nissen, T.N.; Ladekarl, M.; Zingmark, V.; Kjærgaard, J.; Jensen, T.M.; Jensen, S.K.; Thostesen, L.M.; Kofoed, P.E.; Stensballe, L.G.; et al. The association between Bacillus Calmette-Guerin vaccination (1331 SSI) skin reaction and subsequent scar development in infants. BMC Infect. Dis. 2017, 17, 540. [Google Scholar] [CrossRef]
- Zaharia, O.P.; Strassburger, K.; Strom, A.; Bonhof, G.J.; Karusheva, Y.; Antoniou, S.; Bodis, K.; Markgraf, D.F.; Burkart, V.; Mussig, K.; et al. Risk of diabetes-associated diseases in subgroups of patients with recent-onset diabetes: A 5-year follow-up study. Lancet Diabetes Endocrinol. 2019, 7, 684–694. [Google Scholar] [CrossRef]
- Zaharia, O.P.; Bobrov, P.; Strassburger, K.; Bodis, K.; Karusheva, Y.; Scholz, M.; Markgraf, D.F.; Burkart, V.; Schloot, N.C.; Mussig, K.; et al. Metabolic Characteristics of Recently Diagnosed Adult-Onset Autoimmune Diabetes Mellitus. J. Clin. Endocrinol. Metab. 2018, 103, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Campos, C.G.; Veras, H.C.T.; de Aquino Ribeiro, J.A.; Costa, P.; Araujo, K.P.; Rodrigues, C.M.; de Almeida, J.R.M.; Abdelnur, P.V. New Protocol Based on UHPLC-MS/MS for Quantitation of Metabolites in Xylose-Fermenting Yeasts. J. Am. Soc. Mass Spectrom. 2017, 28, 2646–2657. [Google Scholar] [CrossRef]
- Xu, J.; Chen, Q.; Cai, M.; Han, X.; Lu, H. Ultra-high performance liquid chromatography coupled to tandem mass spectrometry-based metabolomics study of diabetic distal symmetric polyneuropathy. J. Diabetes Investig. 2023, 14, 1110–1120. [Google Scholar] [CrossRef]
- Takahashi, H.; Kuhtreiber, W.M.; Keefe, R.C.; Lee, A.H.; Aristarkhova, A.; Dias, H.F.; Ng, N.; Nelson, K.J.; Bien, S.; Scheffey, D.; et al. BCG vaccinations drive epigenetic changes to the human T cell receptor: Restored expression in type 1 diabetes. Sci. Adv. 2022, 8, eabq7240. [Google Scholar] [CrossRef]
- Kuhtreiber, W.M.; Takahashi, H.; Keefe, R.C.; Song, Y.; Tran, L.; Luck, T.G.; Shpilsky, G.; Moore, L.; Sinton, S.M.; Graham, J.C.; et al. BCG Vaccinations Upregulate Myc, a Central Switch for Improved Glucose Metabolism in Diabetes. iScience 2020, 23, 101085. [Google Scholar] [CrossRef]
- Seddiki, N.; Santner-Nanan, B.; Martinson, J.; Zaunders, J.; Sasson, S.; Landay, A.; Solomon, M.; Selby, W.; Alexander, S.I.; Nanan, R.; et al. Expression of interleukin (IL)-2 and IL-7 receptors discriminates between human regulatory and activated T cells. J. Exp. Med. 2006, 203, 1693–1700. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, X.; Geng, X.; Zhao, H. The IL-2A receptor pathway and its role in lymphocyte differentiation and function. Cytokine Growth Factor Rev. 2022, 67, 66–79. [Google Scholar] [CrossRef]
- Cammann, C.; Rath, A.; Reichl, U.; Lingel, H.; Brunner-Weinzierl, M.; Simeoni, L.; Schraven, B.; Lindquist, J.A. Early changes in the metabolic profile of activated CD8(+) T cells. BMC Cell Biol. 2016, 17, 28. [Google Scholar] [CrossRef]
- Donnelly, R.P.; Finlay, D.K. Glucose, glycolysis and lymphocyte responses. Mol. Immunol. 2015, 68, 513–519. [Google Scholar] [CrossRef]
- Locasale, J.W. New concepts in feedback regulation of glucose metabolism. Curr. Opin. Syst. Biol. 2018, 8, 32–38. [Google Scholar] [CrossRef]
- Rabinowitz, J.D.; Enerback, S. Lactate: The ugly duckling of energy metabolism. Nat. Metab. 2020, 2, 566–571. [Google Scholar] [CrossRef]
- Airaghi, L.; Tedeschi, A. Negative association between occurrence of type 1 diabetes and tuberculosis incidence at population level. Acta Diabetol. 2006, 43, 43–45. [Google Scholar] [CrossRef]
- Park, J.Y.; Choe, Y.J.; Lim, Y.; Kim, H.; Kim, J. Association between the incidence of type 1 diabetes mellitus and tuberculosis or bacillus Calmette-Guerin immunization in children and adolescents. Ann. Pediatr. Endocrinol. Metab. 2023, 28, 251–257. [Google Scholar] [CrossRef]
- Corsenac, P.; Parent, M.E.; Mansaray, H.; Benedetti, A.; Richard, H.; Stager, S.; Rousseau, M.C. Early life Bacillus Calmette-Guerin vaccination and incidence of type 1, type 2, and latent autoimmune diabetes in adulthood. Diabetes Metab. 2022, 48, 101337. [Google Scholar] [CrossRef]
- Karaci, M. Chapter 4—The Protective Effect of the BCG Vaccine on the Development of Type 1 Diabetes in Humans. In The Value of BCG and TNF in Autoimmunity; Faustman, D.L., Ed.; Academic Press: Amsterdam, The Netherlands, 2014; pp. 52–62. [Google Scholar]
- Doupis, J.; Kolokathis, K.; Markopoulou, E.; Efthymiou, V.; Festas, G.; Papandreopoulou, V.; Kallinikou, C.; Antikidou, D.; Gemistou, G.; Angelopoulos, T. The Role of Pediatric BCG Vaccine in Type 1 Diabetes Onset. Diabetes Ther. 2021, 12, 2971–2976. [Google Scholar] [CrossRef]
- Elliott, J.F.; Marlin, K.L.; Couch, R.M. Effect of bacille Calmette-Guérin vaccination on C-peptide secretion in children newly diagnosed with IDDM. Diabetes Care 1998, 21, 1691–1693. [Google Scholar] [CrossRef]
- Dias, H.F.; Mochizuki, Y.; Kuhtreiber, W.M.; Takahashi, H.; Zheng, H.; Faustman, D.L. Bacille Calmette Guerin (BCG) and prevention of types 1 and 2 diabetes: Results of two observational studies. PLoS ONE 2023, 18, e0276423. [Google Scholar] [CrossRef]
- Chang, Y.C.; Lin, C.J.; Hsiao, Y.H.; Chang, Y.H.; Liu, S.J.; Hsu, H.Y. Therapeutic Effects of BCG Vaccination on Type 1 Diabetes Mellitus: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. J. Diabetes Res. 2020, 2020, 8954125. [Google Scholar] [CrossRef]
- Adamczyk, M.; Bartosinska, J.; Raczkiewicz, D.; Kowal, M.; Surdacka, A.; Krasowska, D.; Michalak-Stoma, A.; Krasowska, D. The Expression of Activation Markers CD25 and CD69 Increases during Biologic Treatment of Psoriasis. J. Clin. Med. 2023, 12, 6573. [Google Scholar] [CrossRef]
- Moorlag, S.; Arts, R.J.W.; van Crevel, R.; Netea, M.G. Non-specific effects of BCG vaccine on viral infections. Clin. Microbiol. Infect. 2019, 25, 1473–1478. [Google Scholar] [CrossRef]
- Kumar, N.P.; Padmapriyadarsini, C.; Rajamanickam, A.; Bhavani, P.K.; Nancy, A.; Jayadeepa, B.; Selvaraj, N.; Asokan, D.; Renji, R.M.; Venkataramani, V.; et al. BCG vaccination induces enhanced frequencies of memory T cells and altered plasma levels of common gammac cytokines in elderly individuals. PLoS ONE 2021, 16, e0258743. [Google Scholar] [CrossRef]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Benn, C.S.; Joosten, L.A.B.; Jacobs, C.; Van Loenhout, J.; Xavier, R.J.; Aaby, P.; Van Der Meer, J.W.M.; et al. Long-Lasting Effects of BCG Vaccination on Both Heterologous Th1/Th17 Responses and Innate Trained Immunity. J. Innate Immun. 2014, 6, 152–158. [Google Scholar] [CrossRef]
- Goodridge, H.S.; Ahmed, S.S.; Curtis, N.; Kollmann, T.R.; Levy, O.; Netea, M.G.; Pollard, A.J.; van Crevel, R.; Wilson, C.B. Harnessing the beneficial heterologous effects of vaccination. Nat. Rev. Immunol. 2016, 16, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Obar, J.J.; Molloy, M.J.; Jellison, E.R.; Stoklasek, T.A.; Zhang, W.; Usherwood, E.J.; Lefrancois, L. CD4+ T cell regulation of CD25 expression controls development of short-lived effector CD8+ T cells in primary and secondary responses. Proc. Natl. Acad. Sci. USA 2010, 107, 193–198. [Google Scholar] [CrossRef]
- Herndler-Brandstetter, D.; Schwaiger, S.; Veel, E.; Fehrer, C.; Cioca, D.P.; Almanzar, G.; Keller, M.; Pfister, G.; Parson, W.; Wurzner, R.; et al. CD25-expressing CD8+ T cells are potent memory cells in old age. J. Immunol. 2005, 175, 1566–1574. [Google Scholar] [CrossRef]
- Palmer, C.S.; Ostrowski, M.; Balderson, B.; Christian, N.; Crowe, S.M. Glucose metabolism regulates T cell activation, differentiation, and functions. Front. Immunol. 2015, 6, 1. [Google Scholar] [CrossRef]
- Bermingham, K.M.; Mazidi, M.; Franks, P.W.; Maher, T.; Valdes, A.M.; Linenberg, I.; Wolf, J.; Hadjigeorgiou, G.; Spector, T.D.; Menni, C.; et al. Characterisation of Fasting and Postprandial NMR Metabolites: Insights from the ZOE PREDICT 1 Study. Nutrients 2023, 12, 2638. [Google Scholar] [CrossRef] [PubMed]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Tannahill, G.M.; Iraci, N.; Gaude, E.; Frezza, C.; Pluchino, S. Metabolic reprograming of mononuclear phagocytes in progressive multiple sclerosis. Front. Immunol. 2015, 6, 106. [Google Scholar] [CrossRef] [PubMed]
- Atlante, A.; de Bari, L.; Bobba, A.; Amadoro, G. A disease with a sweet tooth: Exploring the Warburg effect in Alzheimer’s disease. Biogerontology 2017, 18, 301–319. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Eugenin, E.A.; Subbian, S. Immunometabolism in Tuberculosis. Front. Immunol. 2016, 7, 150. [Google Scholar] [CrossRef] [PubMed]
- Rawat, B.S.; Kumar, D.; Soni, V.; Rosenn, E.H. Therapeutic Potentials of Immunometabolomic Modulations Induced by Tuberculosis Vaccination. Vaccines 2022, 10, 2127. [Google Scholar] [CrossRef]
- Arts, R.J.; Joosten, L.A.; Netea, M.G. Immunometabolic circuits in trained immunity. Semin. Immunol. 2016, 28, 425–430. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aniagyei, W.; Mohayideen, S.; Sarfo-Kantanka, O.; Bittner, S.; Vivekanandan, M.M.; Arthur, J.F.; Boateng, A.O.; Yeboah, A.; Ahor, H.S.; Asibey, S.O.; et al. BCG Vaccination-Associated Lower HbA1c and Increased CD25 Expression on CD8+ T Cells in Patients with Type 1 Diabetes in Ghana. Vaccines 2024, 12, 452. https://doi.org/10.3390/vaccines12050452
Aniagyei W, Mohayideen S, Sarfo-Kantanka O, Bittner S, Vivekanandan MM, Arthur JF, Boateng AO, Yeboah A, Ahor HS, Asibey SO, et al. BCG Vaccination-Associated Lower HbA1c and Increased CD25 Expression on CD8+ T Cells in Patients with Type 1 Diabetes in Ghana. Vaccines. 2024; 12(5):452. https://doi.org/10.3390/vaccines12050452
Chicago/Turabian StyleAniagyei, Wilfred, Sumaya Mohayideen, Osei Sarfo-Kantanka, Sarah Bittner, Monika M. Vivekanandan, Joseph F. Arthur, Agnes O. Boateng, Augustine Yeboah, Hubert S. Ahor, Shadrack O. Asibey, and et al. 2024. "BCG Vaccination-Associated Lower HbA1c and Increased CD25 Expression on CD8+ T Cells in Patients with Type 1 Diabetes in Ghana" Vaccines 12, no. 5: 452. https://doi.org/10.3390/vaccines12050452