1. Introduction
Plague, caused by the bacterium,
Yersinia pestis, is a zoonotic disease of wild rodents that has severely impacted mammalian populations since its introduction into North America in the early 1900s [
1]. In fact, the disease probably contributed to the demise of several endangered species or at least hinders their recovery [
2], including the black-footed ferret (
Mustela nigripes) and the Utah prairie dog (
Cynomys parvidens). Although plague in humans is rare in the U.S. (3–10 cases per year), its occurrence causes alarm, and outbreaks in rodents can result in park closures or curtailment of other human activities. For these reasons, methods to manage sylvatic plague are highly desired. Although pesticides are effective in reducing fleas that transmit the disease, their application is labor intensive and costly, and the effects of the general insecticides used (e.g., deltamethrin) are indiscriminate, killing other arthropods. As an alternative, we have developed a highly efficacious, orally-delivered sylvatic plague vaccine (SPV) that protects prairie dogs against plague challenge.
Our approach is similar to the successful oral vaccination program for rabies in European and North American carnivores that utilizes vaccinia as a vector for rabies glycoprotein [
3,
4]. Using a similar orthopoxvirus, raccoonpox (RCN), we have designed and tested several vaccine constructs that express antigens known to be protective against plague [
5,
6]. Our first vaccine construct expressed the F1 capsular antigen (designated RCN-F1) and was shown to be protective against plague in both laboratory mice [
5] and prairie dogs [
7]. Due to the occurrence of F1 negative strains of
Y. pestis, we designed a second vaccine construct expressing a truncated version of the
lcrV gene (herein designated RCN-V307). Studies confirmed the protective efficacy of truncated V constructs against F1 negative strains [
8] and further demonstrated that combined administration of RCN-F1 and RCN-V307 increased protection against plague in both mice [
9] and prairie dogs [
6]. However, for a large-scale vaccination program, a single vaccine construct expressing both antigens is more desirable as it would significantly reduce costs of production.
Here, we describe the construction of an RCN-vectored vaccine expressing both F1 and V307 antigens (designated RCN-F1/V307). The efficacy of RCN-F1/V307 was compared to that of RCN-F1, RCN-V307, and combined RCN-F1 + RCN-V307 in a mouse challenge model that included both F1 positive and F1 negative strains of Y. pestis. We also evaluated plague protection in prairie dogs that were orally vaccinated after consuming baits containing the dual antigen construct.
2. Experimental
2.1. Cells and Viruses
Rat embryonic fibroblasts [Rat-2 (ATCC #CRL-1764)] and African green monkey kidney epithelial [BSC-1 (ATCC #CCL-26) and Vero (ATCC#CCL-18)] cells were maintained at 37 °C and 5% CO
2 in M199 medium supplemented with 0.01 g/L L-glutamine and 5% fetal bovine serum (FBS) and were used for culturing virus. Raccoon poxvirus (RCN) Herman strain [
10] was mixed 1:1 with trypsin-versene solution (0.05% trypsin; 0.02% EDTA in Earle’s Balanced Salt Solution) and incubated for 15 min at 37 °C prior to inoculation onto cells.
2.2. Construction of pTK Transfer Vectors
Construction of RCN-F1 and RCN-V307 viruses was described previously [
5,
9]. For the RCN-F1/V307 construct, both F1 and
lcrV (V307, coding for a 307-aa C-terminally truncated V antigen) genes from
Y. pestis were cloned into the pTK shuttle vector, so that the introduced genes and upstream
cis-acting elements were flanked by RCN thymidine kinase gene (
tk) sequences (
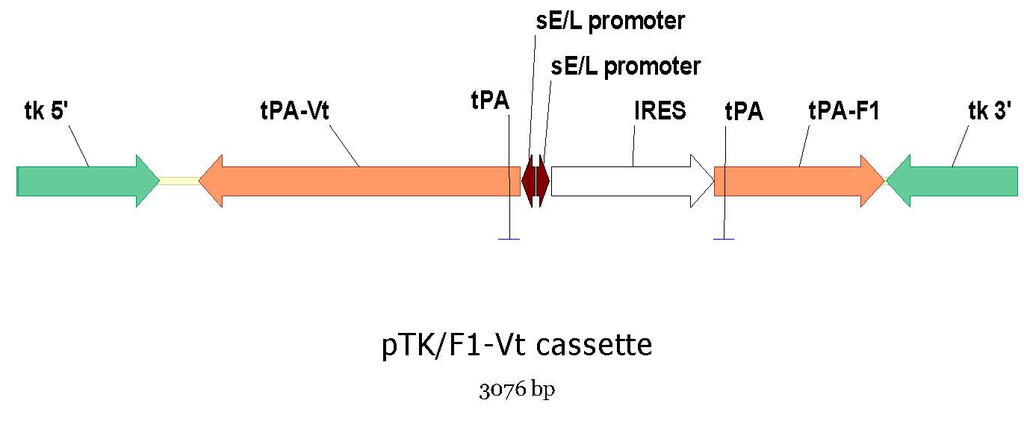
Figure 1). The internal ribosomal entry site (IRES) from encephalomyocarditis virus (EMCV-IRES) preceding the V307 gene was removed to avoid the interferences observed when two IRES are present in the same construct. In an attempt to increase antigen expression, the synthetic strong early/late promoter (sE/L) was used to express both F1 and V307 genes. The tissue plasminogen activator (tPA) secretory signal sequence was inserted in-frame before both F1 and V307 genes (
Figure 1).
Figure 1.
Schematic of the plasmid (pTK) used to produce the dual antigen plague vaccine construct, raccoon poxvirus expressing F1 and V antigens (RCN-F1/V307). The internal ribosomal entry site (IRES) was used as a translation enhancer for the F1 gene. The F1 and V genes were fused to the tissue plasminogen activator (tPA) secretory signal.
Figure 1.
Schematic of the plasmid (pTK) used to produce the dual antigen plague vaccine construct, raccoon poxvirus expressing F1 and V antigens (RCN-F1/V307). The internal ribosomal entry site (IRES) was used as a translation enhancer for the F1 gene. The F1 and V genes were fused to the tissue plasminogen activator (tPA) secretory signal.
2.3. Generation of Recombinant RCN Constructs
The
Y. pestis genes were inserted into the RCN genome by homologous recombination as previously described [
5]. Briefly, BSC-1 cells were infected with wild type RCN and transfected with the pTK shuttle vector. Three days post-infection/transfection, cells were harvested and the lysate was re-plated on Rat-2 cells in medium containing 5-bromo-2-deoxyuridine (BrdU) for selection of TK-RCNV. Purified viral clones were then amplified for large scale purification in Vero cells as described previously [
5] and passaged 10 times to confirm the stability of the insert using PCR.
In vitro expression was determined by western blot as described previously, using polyclonal mouse anti-V and polyclonal rabbit anti-F1 antibodies, secondary antibodies conjugated to alkaline-phosphatase (KPL Inc., Gaithersburg, MD, USA) and BCIP/NBT revelation (KPL Inc.).
2.4. Mouse Vaccination and Challenge Studies
All animal studies were conducted at the USGS National Wildlife Health Center (NWHC) in accordance with NWHC’s animal care and use committee (NWHC #EP050614-A1 and #EP100421). First, we compared the protective efficacy of the rRCN single and dual antigen constructs in mice. Five groups of twenty-four A/J mice (4 week-old) received one of the following treatments: RCN-F1, RCN-V307, a simultaneous injection of RCN-F1 and RCN-V307, the RCN-F1/V307 dual construct or PBS (negative control). Treatments were injected intramuscularly (i.m.) in the thigh with a dose of 5 × 107 pfu in 0.1 mL. Animals that received two rRCN constructs were inoculated with each virus in separate thighs. All groups received a boost vaccination (same formulations/dosage/route) four weeks later. To determine the kinetics of vaccine immune responses and protection, six animals in each group were challenged at 7, 14, 28, and 56 days post booster vaccination (dpb), with Y. pestis (CO92 isolate, provided by U.S. Army Medical Research Institute of Infectious Diseases-USAMRIID) at a dose of 1.4 × 104 mouse LD50s by intradermal (i.d.) injection in the abdominal region. The mice were monitored for 21 days for signs of illness or death. Animals with obvious clinical signs of plague (labored breathing, severe lethargy, etc.) were humanely euthanized.
To confirm the efficacy of the dual antigen construct against varying doses of plague, groups of six A/J mice each were vaccinated with RCN-F1/V307, same dose and route as described above, and boosted four weeks later. Three groups of four mice each served as controls. The mice were all bled on 21 dpb and each group of vaccine-treated and control mice were challenged as above with varying doses of Y. pestis CO92 strain (1.4 × 103, 1.4 × 104, or 8.0 × 105 mouse LD50s).
The third study examined the efficacy of the RCN-F1/V307 dual antigen construct against a Y. pestis F1 negative strain. Three groups of 24 A/J mice were vaccinated and boosted with either RCN-F1, simultaneous injections of RCN-F1 and RCN-V307, or RCN-F1/V307 as described above. At 21 and 42 dpb, 6 animals in each group were challenged as above with an F1 negative strain of Y. pestis (Java 9, YERS022, provided by USAMRIID) at a dose of 1.4 × 104 or 1.4 × 105 LD50s.
2.5. Prairie Dog Oral Vaccination and Challenge Study
Finally, we evaluated the efficacy of the RCN-F1/V307 dual antigen construct delivered via edible baits in prairie dogs (PDs), our primary target animal. Adult black-tailed prairie dogs (
Cynomys ludovicianus) were captured from wild colonies near Wall, SD, USA (43.992N, 102.241W) and dusted with carbaryl prior to shipment to NWHC. Upon arrival, they were inspected for external parasites (none found), injected with an anthelminthic (200 μg/kg of Ivermectin, Merck & Co., Inc, West Point, PA, USA), then treated with Advantage flea control (Imidacloprid; Bayer HealthCare, Shawnee Mission, KS) and inserted with micropchips (Avid Identification Systems, Inc., Folsom, LA, USA) to identify individuals. Blood was also drawn upon arrival to test for antibodies to plague antigens as described below, and the animals were group-housed and fed as previously described [
6]. The animals were randomly assigned to one of five treatment groups and housed in separate rooms.
RCN constructs (RCN-F1/V307 or RCN-TK-) were mixed into 4 g baits and offered to PDs for voluntary consumption. Three groups were fed baits containing RCN-F1/V307; one of these groups (group 4) also received baits a second time at 240 days post initial vaccination (dpv). Two control groups received baits containing the empty vector (RCN-TK-) as a placebo, designated groups 1a and 1b. The bait formulation was supplied by a commercial source (Food Source Lures, Alabaster, AL, USA) and peanut butter was added to increase palatability. To increase vaccine or placebo intake by PDs, fresh vegetables were withheld for 48 h prior to vaccination and food pellets were withheld for 12–18 h prior to vaccination. Animals were individually placed in pet carriers with either one vaccine-laden bait or one placebo bait. After 2–4 h, animals were released back into their animal rooms, and bait consumption was recorded. This process was performed again the next day, so that all animals were offered two baits. Group 4 animals, that received vaccine baits a second time, were treated similarly. Animals that did not eat any baits were removed from the study (one animal).
Blood samples were drawn from prairie dogs at approximately 30 dpv (n = 70), 180 dpv (n = 41) and 270 dpv (n = 41). At 30 dpv, one vaccine-treated (group 2) and one placebo group (group 1a) were challenged with Y. pestis. On 270 dpv, the remaining three groups were challenged: one vaccine-treated group (group 3), one vaccine treated group that consumed baits a second time 240 dpv (group 4), and the remaining placebo group (1b). PDs were challenged with Y. pestis (CO92) at a dosage of 3500 mouse LD50s via subcutaneous injection in the right hip region. They were monitored 2–3 times daily for 28 days for signs of illness or death. Carcasses were removed immediately upon discovery and stored at −20 °C until processed. Animals with obvious clinical signs (labored breathing, disinclination to move) were humanely euthanized, as were all survivors at the end of the 28-day period. Tissue samples for bacteriology were aseptically collected and stored at −20 °C, and plague was confirmed as the cause of death in selected animals.
2.6. Serology
Blood samples were collected from the medial saphenous vein of mice and PDs. Sera were collected and stored at −20 °C. Antibody titers (total IgG, IgG1, and IgG2c for mice and IgG only for PDs) to
Y. pestis F1 and V were determined by ELISA [
5,
11]. Serum samples were serially diluted 4-fold from 1:160 to 1:163,840 and tested in duplicate. Titers ≤1:160 were considered negative and titers <1:160 were designated as 1:40 for analyses. The highest dilution that was positive (exceeded the mean of four negative control samples by three standard deviations) was considered the endpoint and its reciprocal value recorded as the titer.
2.7. Statistical Analyses
All analyses were performed with SAS statistical software (SAS Institute Inc., Cary NC, USA). The effects of treatments and antibody titers on survival rates were evaluated with the Cox proportional hazards model, and PD survival curves were generated using Kaplan Meier analysis. Antibody titers were log-transformed prior to analysis. Pre-challenge geometric anti-F1 and anti-V titers and interactions between the two were analyzed in relation to time to death using the Cox proportional hazards model for the first and third mouse experiments. Data from all mouse experiments were combined and treatment groups were compared using analysis of variance. Likewise, antibody titers of PDs consuming vaccine baits were combined and compared at each time point using analysis of variance.
4. Discussion
Like other orthopoxviruses, RCN has proven to be an efficient vector for vaccine antigens, capable of delivering several antigens simultaneously. In this paper, we describe the construction of an RCN vectored vaccine expressing two protective
Y. pestis antigens (designated RCN-F1/V307) and compare the efficacy of this dual antigen vaccine construct to the simultaneous administration of two single antigen vaccines (RCN-F1 + RCN-V307). The RCN-F1/V307 was found to be highly stable following
in vitro serial passage in Vero cells and expressed both F1 and V307 antigens. In mice, RCN-F1/V307 was as effective at preventing plague as the simultaneous administration of RCN-F1 + RCN-V307. Although the dual antigen construct was a little slower in inducing protection than the combination vaccine, by 56 dpb, mice vaccinated with RCN-F1/V307 had similar survival rates (67%) as those vaccinated with RCN-F1 (50%) or RCN-F1 + RCN-V307 (33%). At least 50% of mice vaccinated with RCN-F1/V307 survived
Y. pestis challenge doses as high as 8 × 10
5 mouse LD
50s. This protective efficacy is similar to our previous findings in mice vaccinated simultaneously with the two single antigen constructs [
9].
Even more importantly, our findings demonstrated that RCN-F1/V307 can induce significant protection in orally vaccinated PDs, even after a single administration of the vaccine. Protective efficacy seems to improve with time (up to 60% at approximately 9 months post vaccination), although additional work is required to establish the kinetics of protection at both shorter (1–9 months) and longer times (>9 months) post vaccination. A booster vaccination via consumption of baits a second time provided even greater protection (85%) in PDs. This level of protection is very similar to results achieved in PDs (94%) that consumed baits with the combination vaccine (RCN/F1 + RCN/V307), with a 178 day interval between initial and booster vaccinations [
6]. These results will help inform the design and evaluation of future field studies in wild PD colonies.
We further demonstrated that RCN-F1/V307 protects mice against an F1 negative strain of Y. pestis. Mice vaccinated with RCN-F1 alone succumbed to challenge with F1 negative Y. pestis, whereas, approximately half the mice that received either RCN-F1/V307 or simultaneous injections of RCN-F1 and RCN-V307 survived the same challenge. Both these groups of mice had high anti-V antibody titers. Although not tested yet, we hypothesize that orally vaccinated PDs would also be protected against an F1 negative Y. pestis strain.
Previous work showed that a truncated version of the
lcrV gene (V307) was as effective as the entire V gene at eliciting a protective immune response in combination with F1 [
9]. This V307 antigen presumably avoids some of the immunomodulatory properties associated with full V protein [
13], including suppression of gamma interferon and tumor necrosis factor alpha
in vivo [
14,
15]. However, vaccination with RCN-V307 alone was not very effective and resulted in the poorest survival rate upon plague challenge of all the groups. Administration of RCN-F1/V307 or the combination of RCN-F1 and RCN-V307 resulted in the highest rates of protection against plague in mice. This result supports our previous findings in mice that simultaneous vaccination with two antigens provides increased protection against plague challenges [
6,
9].
Interestingly, vaccination with the dual antigen vaccine resulted in different antibody responses compared to the combination vaccine. In mice, administration of RCN-F1/V307 elicited significantly higher antibody titers to V compared to the combination vaccine but lower titers to F1.
In vitro results also demonstrated higher levels of V307 expression in the RCN-F1/V307 construct, which suggests that simultaneous expression of
Y. pestis antigens may result in F1 interference. We have previously observed this phenomenon in the design of rRCN vaccines against plague and other pathogens [
16]. It is possible that this viral interference can be overcome by using different viral promoters or stronger translation enhancer sequences. Despite the interference, in this study, RCN-F1/V307 expressed sufficient antigen levels to confer protection against both F1 positive and F1 negative strains.
Similar serologic results were obtained with PDs that consumed baits containing RCN-F1/V307, although titers were much lower relative to mice, likely due to the different routes of vaccine administration (oral
versus i.m.). Unfortunately, laboratory mice are not susceptible to infection by RCN via the oral route [
17] for better comparison. Anti-V titers were slightly higher in PDs than anti-F1 titers, and the levels of both were very similar to those reported in PDs that consumed baits with the combination vaccine [
6]. Protection of PDs against plague in the absence of high IgG titers suggests other mechanisms of immunity may be playing a role in protection. Other poxvirus recombinants (vaccinia expressing the luciferase reporter gene, HIV env protein, or β-galactosidase) delivered orally to mice resulted in significant mucosal IgA responses and both local and systemic cellular immune responses [
18]. Future studies will determine the role of mucosal and cellular immune responses in mice and PDs.
5. Conclusions
An ideal plague vaccine for wildlife would be safe, highly efficacious, stable, deliverable orally, easy to distribute to large numbers of animals in the field, and of relatively low cost to produce. Our dual antigen vaccine construct, as described here, meets all those requirements. Our data confirms that RCN-F1/V307 is stable, efficacious in protecting PDs in laboratory challenge studies, and can be delivered orally via baits. Consumption of baits a single time can be protective, but a second consumption provides even higher rates of protection, suggesting that booster vaccinations may be necessary to achieve full protection in the field. In this and previous studies, we exposed PDs to Y. pestis via the subcutaneous route to achieve a standardized challenge inoculum, but in the field, plague exposure occurs via multiple flea bites and possibly via other means. Because different routes of exposure could affect the outcome of vaccination, field studies will be needed to determine if SPV is efficacious in preventing plague in nature.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
