
M2e-Based Universal Influenza A Vaccines


Abstract
:1. Introduction
2. Biological Function of M2

3. M2-Specific Immune Responses Following Infection
4. T Cell Epitopes in M2e

5. M2e-Based Vaccines

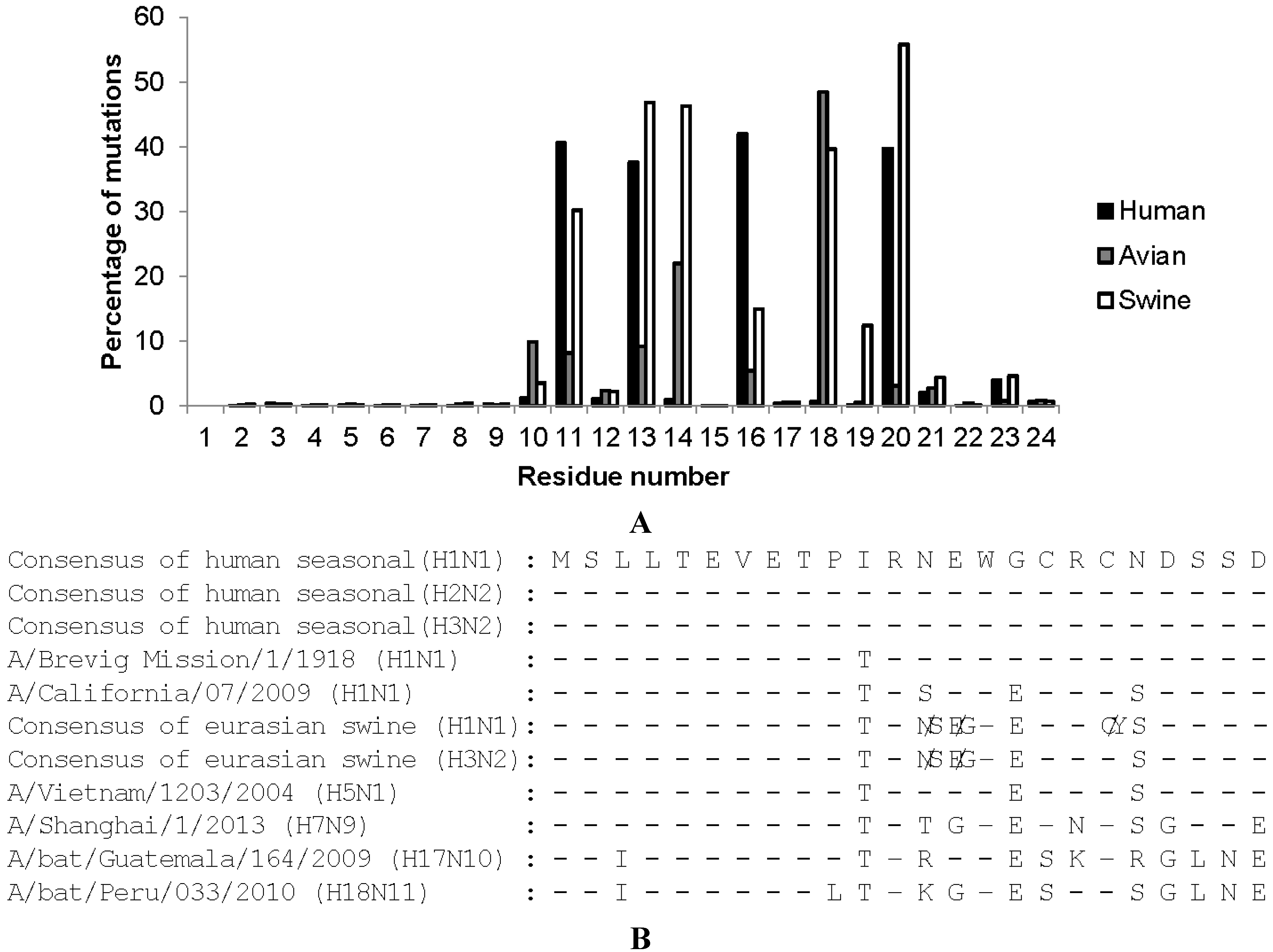{kind=link}
{kind=link}
{kind=link}
| Overview of M2e Based Vaccines | |||||
|---|---|---|---|---|---|
| Vaccine Type | Carriers | Copy Numbers | Antigen Type | Immunogenicity Readout in Animal Models (Administration Routes) | Reference |
| VLPs | HBc | 1, 2, 3 | human | Mice (intranasal, intraperitoneal) Pigs (intramuscular), Human | [32,111,114,115,116] |
| HA(TM) | 5 | human, swine, avian | Mice (intramuscular) | [69] | |
| MaMV | 3 | canine | mice (subcutaneous), dogs (intramuscular) | [70] | |
| Tobacco mosaic virus coat protein | 1 | human | Mice | [71] | |
| Papaya mosaic virus | 1 | human | Mice (subcutaneous) | [73] | |
| Woodchuck hepatitis VLP vectored in Salmonella Typhimurium | 1 | avian-like | Mice (oral) | [75] | |
| T7 | 1 | human | Mice (subcutaneous) | [76] | |
| Q-β | 1 | human | Mice (intranasal, subcutaneous) | [77] | |
| DNA | Complete NP | 1 | swine | Pigs (intradermal) | [111] |
| VP22, tegument protein of bovine herpesvirus-1 | 1 | human | Mice (intramuscular) | [93] | |
| HA, NP (147-155) | 1 | human | Mice (gene gun) | [94] | |
| HA | 1,2 | human, avian | Mice (gene gun, intramuscular) | [94,95] | |
| peptide | - | 1 | human | Mice (subcutaneous) | [65] |
| Multiple antigen peptide | 1, 4 | human, avian | Mice (intranasal, subcutaneous) | [56,80,81,82,83] | |
| protein | Influenza NP | 8 | - | Mice | [117] |
| CTA1-DD | 1, 3 | human | Mice (intranasal) | [64] | |
| tGCN4 | tetramer | human | Mice (intraperitoneal, intranasal) | [78] | |
| rotavirus fragment NSP4 | tetramer | human | Mice (subcutaneous) | [79] | |
| KLH | 1 | human, avian | Mice (subcutaneous, intramuscular), Ferrets (intramuscular), Rabbit | [66,85] | |
| OMPC | 1 | human | Ferrets (intramuscular), Rhesus Monkey (intramuscular) | [85,86] | |
| RSV NP | 1, 3 | human | Mice (intranasal, subcutaneous) | [91] | |
| BLS | 1, 4 | human | Mice (intranasal, subcutaneous, intramuscular) | [90] | |
| glutathione S-transferase | 1, 4, 8 | human | Rabbit (subcutaneous) | [118] | |
| flagelin | 4 | human | Human (intramuscular), Mice (subcutaneous, intranasal) | [87] | |
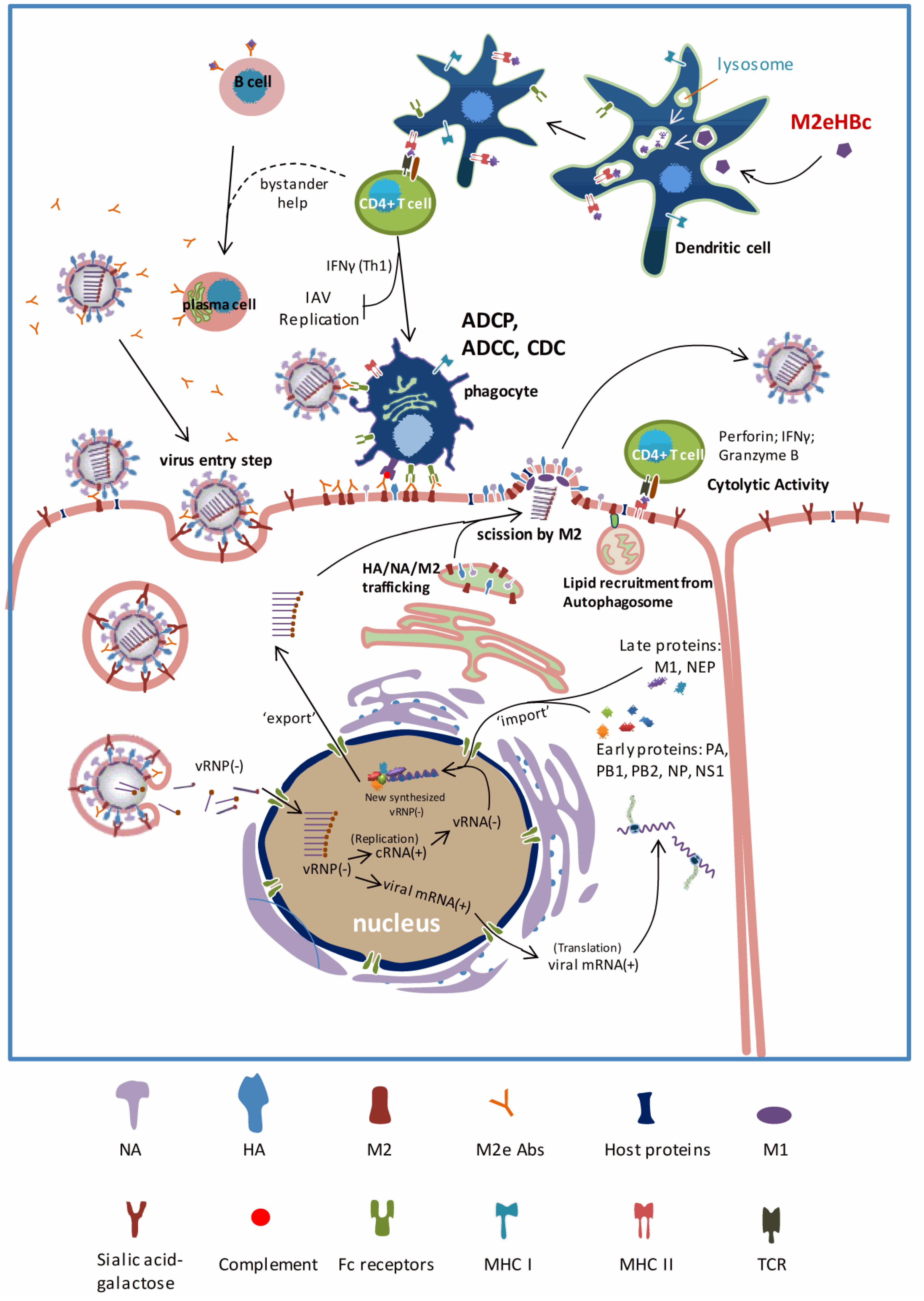
6. Mechanisms of Protection by M2e-Based Vaccines: Universal Protection and Beyond
7. Clinical Development of M2e-Based Vaccines
8. Conclusions and Future Perspectives

Acknowledgments
Author Contributions
Conflicts of Interest
References
- CDC. Estimates of deaths associated with seasonal influenza—United States, 1976–2007. In MMWR Morb. Mortal. Wkly. Rep.; 2010; 59, pp. 1057–1062. [Google Scholar]
- Barker, W.H.; Mullooly, J.P. Pneumonia and influenza deaths during epidemics: Implications for prevention. Arch. Intern. Med. 1982, 142, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Nair, H.; Brooks, W.A.; Katz, M.; Roca, A.; Berkley, J.A.; Madhi, S.A.; Simmerman, J.M.; Gordon, A.; Sato, M.; Howie, S.; et al. Global burden of respiratory infections due to seasonal influenza in young children: A systematic review and meta-analysis. Lancet 2011, 378, 1917–1930. [Google Scholar] [CrossRef] [PubMed]
- Patterson, K.D.; Pyle, G.F. The geography and mortality of the 1918 influenza pandemic. Bull. Hist. Med. 1991, 65, 4–21. [Google Scholar] [PubMed]
- Johnson, N.P.; Mueller, J. Updating the accounts: Global mortality of the 1918–1920 “Spanish” influenza pandemic. Bull. Hist. Med. 2002, 76, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Simonsen, L.; Clarke, M.J.; Williamson, G.D.; Stroup, D.F.; Arden, N.H.; Schonberger, L.B. The impact of influenza epidemics on mortality: Introducing a severity index. Am. J. Public Health 1997, 87, 1944–1950. [Google Scholar] [CrossRef] [PubMed]
- Dawood, F.S.; Iuliano, A.D.; Reed, C.; Meltzer, M.I.; Shay, D.K.; Cheng, P.Y.; Bandaranayake, D.; Breiman, R.F.; Brooks, W.A.; Buchy, P.; et al. Estimated global mortality associated with the first 12 months of 2009 pandemic influenza A H1N1 virus circulation: A modelling study. Lancet Infect. Dis. 2012, 12, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Jiang, D.; Wang, H.; Zhuang, D.; Ma, J.; Fu, J.; Qu, J.; Sun, Y.; Yu, S.; Meng, Y.; et al. Calculating the burden of disease of avian-origin H7N9 infections in China. Br. Med. J. Open 2014, 4, e004189. [Google Scholar]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar] [PubMed]
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco, S.; Gomez, J.; et al. New world bats harbor diverse influenza A viruses. PLOS Pathog. 2013, 9, e1003657. [Google Scholar] [CrossRef] [PubMed]
- Chaipan, C.; Kobasa, D.; Bertram, S.; Glowacka, I.; Steffen, I.; Tsegaye, T.S.; Takeda, M.; Bugge, T.H.; Kim, S.; Park, Y.; et al. Proteolytic activation of the 1918 influenza virus hemagglutinin. J. Virol. 2009, 83, 3200–3211. [Google Scholar] [CrossRef] [PubMed]
- Matrosovich, M.N.; Matrosovich, T.Y.; Gray, T.; Roberts, N.A.; Klenk, H.D. Human and avian influenza viruses target different cell types in cultures of human airway epithelium. Proc. Natl. Acad. Sci. USA 2004, 101, 4620–4624. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, B.; Blazejewska, P.; Hessmann, M.; Bruder, D.; Geffers, R.; Mauel, S.; Gruber, A.D.; Schughart, K. Host genetic background strongly influences the response to influenza a virus infections. PLOS ONE 2009, 4, e4857. [Google Scholar] [CrossRef] [PubMed]
- Gouarin, S.; Vabret, A.; Dina, J.; Petitjean, J.; Brouard, J.; Cuvillon-Nimal, D.; Freymuth, F. Study of influenza C virus infection in France. J. Med. Virol. 2008, 80, 1441–1446. [Google Scholar] [CrossRef] [PubMed]
- Kauppila, J.; Ronkko, E.; Juvonen, R.; Saukkoriipi, A.; Saikku, P.; Bloigu, A.; Vainio, O.; Ziegler, T. Influenza C virus infection in military recruits—Symptoms and clinical manifestation. J. Med. Virol. 2014, 86, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, Y.; Katsushima, N.; Nagai, Y.; Shoji, M.; Itagaki, T.; Sakamoto, M.; Kitaoka, S.; Mizuta, K.; Nishimura, H. Clinical features of influenza C virus infection in children. J. Infect. Dis. 2006, 193, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Kendal, A.P. Cold-adapted live attenuated influenza vaccines developed in Russia: Can they contribute to meeting the needs for influenza control in other countries? Eur. J. Epidemiol. 1997, 13, 591–609. [Google Scholar] [CrossRef] [PubMed]
- Isakova-Sivak, I.; Chen, L.M.; Matsuoka, Y.; Voeten, J.T.; Kiseleva, I.; Heldens, J.G.; den Bosch, H.; Klimov, A.; Rudenko, L.; Cox, N.J.; et al. Genetic bases of the temperature-sensitive phenotype of a master donor virus used in live attenuated influenza vaccines: A/Leningrad/134/17/57 (H2N2). Virology 2011, 412, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Durviaux, S.; Treanor, J.; Beran, J.; Duval, X.; Esen, M.; Feldman, G.; Frey, S.E.; Launay, O.; Leroux-Roels, G.; McElhaney, J.E.; et al. Genetic and antigenic typing of seasonal influenza virus breakthrough cases from a 2008–2009 vaccine efficacy trial. Clin. Vaccine Immunol. 2014, 21, 271–279. [Google Scholar] [CrossRef] [PubMed]
- El Moussi, A.; Pozo, F.; Ben Hadj Kacem, M.A.; Ledesma, J.; Cuevas, M.T.; Casas, I.; Slim, A. Virological Surveillance of Influenza Viruses during the 2008–09, 2009–10 and 2010–11 Seasons in Tunisia. PLOS ONE 2013, 8, e74064. [Google Scholar]
- Heikkinen, T.; Ikonen, N.; Ziegler, T. Impact of influenza B lineage-level mismatch between trivalent seasonal influenza vaccines and circulating viruses, 1999–2012. Clin. Infect. Dis. 2014. [Google Scholar] [CrossRef]
- McKeage, K. Inactivated quadrivalent split-virus seasonal influenza vaccine (Fluarix(R) quadrivalent): A review of its use in the prevention of disease caused by influenza A and B. Drugs 2013, 73, 1587–1594. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, D.P.; Robertson, C.A.; Landolfi, V.A.; Bhaumik, A.; Senders, S.D.; Decker, M.D. Safety and immunogenicity of an inactivated quadrivalent influenza vaccine in children 6 months through 8 years of age. Pediatr. Infect. Dis. J. 2014, 33, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Ekiert, D.C.; Krause, J.C.; Hai, R.; Crowe, J.E., Jr.; Wilson, I.A. Structural basis of preexisting immunity to the 2009 H1N1 pandemic influenza virus. Science 2010, 328, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Kirchenbaum, G.A.; Ross, T.M. Eliciting broadly protective antibody responses against influenza. Curr. Opin. Immunol. 2014, 28, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Vasin, A.V.; Temkina, O.A.; Egorov, V.V.; Klotchenko, S.A.; Plotnikova, M.A.; Kiselev, O.I. Molecular mechanisms enhancing the proteome of influenza A viruses: An overview of recently discovered proteins. Virus Res. 2014, 185, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Lamb, R.A.; Choppin, P.W. Identification of a second protein (M2) encoded by RNA segment 7 of influenza virus. Virology 1981, 112, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Lamb, R.A.; Lai, C.J.; Choppin, P.W. Sequences of mRNAs derived from genome RNA segment 7 of influenza virus: Colinear and interrupted mRNAs code for overlapping proteins. Proc. Natl. Acad. Sci. USA 1981, 78, 4170–4174. [Google Scholar] [CrossRef] [PubMed]
- Leiding, T.; Wang, J.; Martinsson, J.; DeGrado, W.F.; Arskold, S.P. Proton and cation transport activity of the M2 proton channel from influenza A virus. Proc. Natl. Acad. Sci. USA 2010, 107, 15409–15414. [Google Scholar] [CrossRef] [PubMed]
- Stauffer, S.; Feng, Y.; Nebioglu, F.; Heilig, R.; Picotti, P.; Helenius, A. Stepwise priming by acidic pH and a high K+ concentration is required for efficient uncoating of influenza A virus cores after penetration. J. Virol. 2014, 88, 13029–13046. [Google Scholar] [CrossRef] [PubMed]
- Helenius, A. Unpacking the incoming influenza virus. Cell 1992, 69, 577–578. [Google Scholar] [CrossRef] [PubMed]
- Neirynck, S.; Deroo, T.; Saelens, X.; Vanlandschoot, P.; Jou, W.M.; Fiers, W. A universal influenza A vaccine based on the extracellular domain of the M2 protein. Nat. Med. 1999, 5, 1157–1163. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhou, L.; Chen, Y.H. Immunization with high epitope density of M2e derived from 2009 pandemic H1N1 elicits protective immunity in mice. Vaccine 2012, 30, 3463–3469. [Google Scholar] [CrossRef] [PubMed]
- Black, R.A.; Rota, P.A.; Gorodkova, N.; Klenk, H.D.; Kendal, A.P. Antibody response to the M2 protein of influenza A virus expressed in insect cells. J. Gen. Virol. 1993, 74, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Wise, H.M.; Hutchinson, E.C.; Jagger, B.W.; Stuart, A.D.; Kang, Z.H.; Robb, N.; Schwartzman, L.M.; Kash, J.C.; Fodor, E.; Firth, A.E.; et al. Identification of a novel splice variant form of the influenza A virus M2 ion channel with an antigenically distinct ectodomain. PLOS Pathog. 2012, 8, e1002998. [Google Scholar] [CrossRef] [PubMed]
- Stouffer, A.L.; Acharya, R.; Salom, D.; Levine, A.S.; di Costanzo, L.; Soto, C.S.; Tereshko, V.; Nanda, V.; Stayrook, S.; DeGrado, W.F. Structural basis for the function and inhibition of an influenza virus proton channel. Nature 2008, 451, 596–599. [Google Scholar] [CrossRef] [PubMed]
- Khurana, E.; dal Peraro, M.; DeVane, R.; Vemparala, S.; DeGrado, W.F.; Klein, M.L. Molecular dynamics calculations suggest a conduction mechanism for the M2 proton channel from influenza A virus. Proc. Natl. Acad. Sci. USA 2009, 106, 1069–1074. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Yi, M.; Dong, H.; Qin, H.; Peterson, E.; Busath, D.D.; Zhou, H.X.; Cross, T.A. Insight into the mechanism of the influenza A proton channel from a structure in a lipid bilayer. Science 2010, 330, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Pielak, R.M.; Schnell, J.R.; Chou, J.J. Mechanism of drug inhibition and drug resistance of influenza A M2 channel. Proc. Natl. Acad. Sci. USA 2009, 106, 7379–7384. [Google Scholar] [CrossRef] [PubMed]
- Schnell, J.R.; Chou, J.J. Structure and mechanism of the M2 proton channel of influenza A virus. Nature 2008, 451, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Pielak, R.M.; Oxenoid, K.; Chou, J.J. Structural investigation of rimantadine inhibition of the AM2-BM2 chimera channel of influenza viruses. Structure 2011, 19, 1655–1663. [Google Scholar] [CrossRef] [PubMed]
- McCown, M.F.; Pekosz, A. The influenza A virus M2 cytoplasmic tail is required for infectious virus production and efficient genome packaging. J. Virol. 2005, 79, 3595–3605. [Google Scholar] [CrossRef] [PubMed]
- McCown, M.F.; Pekosz, A. Distinct domains of the influenza a virus M2 protein cytoplasmic tail mediate binding to the M1 protein and facilitate infectious virus production. J. Virol. 2006, 80, 8178–8189. [Google Scholar] [CrossRef] [PubMed]
- Rossman, J.S.; Jing, X.; Leser, G.P.; Balannik, V.; Pinto, L.H.; Lamb, R.A. Influenza virus m2 ion channel protein is necessary for filamentous virion formation. J. Virol. 2010, 84, 5078–5088. [Google Scholar] [CrossRef] [PubMed]
- Rossman, J.S.; Jing, X.; Leser, G.P.; Lamb, R.A. Influenza virus M2 protein mediates ESCRT-independent membrane scission. Cell 2010, 142, 902–913. [Google Scholar] [CrossRef] [PubMed]
- Ciampor, F.; Bayley, P.M.; Nermut, M.V.; Hirst, E.M.; Sugrue, R.J.; Hay, A.J. Evidence that the amantadine-induced, M2-mediated conversion of influenza A virus hemagglutinin to the low pH conformation occurs in an acidic trans Golgi compartment. Virology 1992, 188, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Betakova, T.; Ciampor, F.; Hay, A.J. Influence of residue 44 on the activity of the M2 proton channel of influenza A virus. J. Gen. Virol. 2005, 86, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Grambas, S.; Hay, A.J. Maturation of influenza A virus hemagglutinin—Estimates of the pH encountered during transport and its regulation by the M2 protein. Virology 1992, 190, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Ichinohe, T.; Pang, I.K.; Iwasaki, A. Influenza virus activates inflammasomes via its intracellular M2 ion channel. Nat. Immunol. 2010, 11, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Deretic, V.; Levine, B. Autophagy, immunity, and microbial adaptations. Cell Host Microbe 2009, 5, 527–549. [Google Scholar] [CrossRef] [PubMed]
- Gannage, M.; Dormann, D.; Albrecht, R.; Dengjel, J.; Torossi, T.; Ramer, P.C.; Lee, M.; Strowig, T.; Arrey, F.; Conenello, G.; et al. Matrix protein 2 of influenza A virus blocks autophagosome fusion with lysosomes. Cell Host Microbe 2009, 6, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Beale, R.; Wise, H.; Stuart, A.; Ravenhill, B.J.; Digard, P.; Randow, F. A LC3-interacting motif in the influenza A virus M2 protein is required to subvert autophagy and maintain virion stability. Cell Host Microbe 2014, 15, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhang, M.; Mozdzanowska, K.; Zharikova, D.; Hoff, H.; Wunner, W.; Couch, R.B.; Gerhard, W. Influenza A virus infection engenders a poor antibody response against the ectodomain of matrix protein 2. Virol. J. 2006. [Google Scholar] [CrossRef]
- Hutchinson, E.C.; Charles, P.D.; Hester, S.S.; Thomas, B.; Trudgian, D.; Martinez-Alonso, M.; Fodor, E. Conserved and host-specific features of influenza virion architecture. Nat. Commun. 2014. [Google Scholar] [CrossRef]
- Heinen, P.P.; de Boer-Luijtze, E.A.; Bianchi, A.T. Respiratory and systemic humoral and cellular immune responses of pigs to a heterosubtypic influenza A virus infection. J. Gen. Virol. 2001, 82, 2697–2707. [Google Scholar] [PubMed]
- Wolf, A.I.; Mozdzanowska, K.; Williams, K.L.; Singer, D.; Richter, M.; Hoffmann, R.; Caton, A.J.; Otvos, L.; Erikson, J. Vaccination with M2e-based multiple antigenic peptides: Characterization of the B cell response and protection efficacy in inbred and outbred mice. PLOS ONE 2011, 6, e28445. [Google Scholar] [CrossRef] [PubMed]
- Hemmatzadeh, F.; Sumarningsih, S.; Tarigan, S.; Indriani, R.; Dharmayanti, N.L.; Ebrahimie, E.; Igniatovic, J. Recombinant M2e protein-based ELISA: A novel and inexpensive approach for differentiating avian influenza infected chickens from vaccinated ones. PLOS ONE 2013, 8, e56801. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.; Steensels, M.; van Borm, S.; Meulemans, G.; van den Berg, T. Development of an M2e-specific enzyme-linked immunosorbent assay for differentiating infected from vaccinated animals. Avian Dis. 2007, 51, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Reed, C.; Blair, P.J.; Katz, J.M.; Hancock, K. Serum antibody response to matrix protein 2 following natural infection with 2009 pandemic influenza A(H1N1) virus in humans. J. Infect. Dis. 2014, 209, 986–994. [Google Scholar] [CrossRef] [PubMed]
- Jameson, J.; Cruz, J.; Terajima, M.; Ennis, F.A. Human CD8+ and CD4+ T lymphocyte memory to influenza A viruses of swine and avian species. J. Immunol. 1999, 162, 7578–7583. [Google Scholar] [PubMed]
- Jameson, J.; Cruz, J.; Ennis, F.A. Human cytotoxic T-lymphocyte repertoire to influenza A viruses. J. Virol. 1998, 72, 8682–8689. [Google Scholar] [PubMed]
- Gianfrani, C.; Oseroff, C.; Sidney, J.; Chesnut, R.W.; Sette, A. Human memory CTL response specific for influenza A virus is broad and multispecific. Hum. Immunol. 2000, 61, 438–452. [Google Scholar] [CrossRef] [PubMed]
- Canaday, D.H.; Gehring, A.; Leonard, E.G.; Eilertson, B.; Schreiber, J.R.; Harding, C.V.; Boom, W.H. T-cell hybridomas from HLA-transgenic mice as tools for analysis of human antigen processing. J. Immunol. Methods 2003, 281, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Eliasson, D.G.; el Bakkouri, K.; Schon, K.; Ramne, A.; Festjens, E.; Lowenadler, B.; Fiers, W.; Saelens, X. CTA1-M2e-DD: A novel mucosal adjuvant targeted influenza vaccine. Vaccine 2008, 26, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Pejoski, D.; Zeng, W.; Rockman, S.; Brown, L.E.; Jackson, D.C. A lipopeptide based on the M2 and HA proteins of influenza A viruses induces protective antibody. Immunol. Cell Biol. 2010, 88, 605–611. [Google Scholar] [CrossRef] [PubMed]
- De Filette, M.; Ysenbaert, T.; Roose, K.; Schotsaert, M.; Roels, S.; Goossens, E.; Schepens, B.; Fiers, W.; Saelens, X. Antiserum against the conserved nine amino acid N-terminal peptide of influenza A virus matrix protein 2 is not immunoprotective. J. Gen. Virol. 2011, 92, 301–306. [Google Scholar]
- Misplon, J.A.; Lo, C.Y.; Gabbard, J.D.; Tompkins, S.M.; Epstein, S.L. Genetic control of immune responses to influenza A matrix 2 protein (M2). Vaccine 2010, 28, 5817–5827. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.N.; Kim, M.C.; Lee, Y.T.; Hwang, H.S.; Cho, M.K.; Lee, J.S.; Ko, E.J.; Kwon, Y.M.; Kang, S.M. AS04-adjuvanted virus-like particles containing multiple M2 extracellular domains of influenza virus confer improved protection. Vaccine 2014, 32, 4578–4585. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.C.; Song, J.M.; Eunju, O.; Kwon, Y.M.; Lee, Y.J.; Compans, R.W.; Kang, S.M. Virus-like particles containing multiple M2 extracellular domains confer improved cross-protection against various subtypes of influenza virus. Mol. Ther. 2013, 21, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, D.; Rivest, M.; Babin, C.; Lopez-Macias, C.; Savard, P. A novel M2e based flu vaccine formulation for dogs. PLOS ONE 2013, 8, e77084. [Google Scholar] [CrossRef] [PubMed]
- Petukhova, N.V.; Gasanova, T.V.; Stepanova, L.A.; Rusova, O.A.; Potapchuk, M.V.; Korotkov, A.V.; Skurat, E.V.; Tsybalova, L.M.; Kiselev, O.I.; Ivanov, P.A.; et al. Immunogenicity and protective efficacy of candidate universal influenza A nanovaccines produced in plants by Tobacco mosaic virus-based vectors. Curr. Pharm. Des. 2013, 19, 5587–5600. [Google Scholar] [CrossRef] [PubMed]
- Tyulkina, L.G.; Skurat, E.V.; Frolova, O.Y.; Komarova, T.V.; Karger, E.M.; Atabekov, I.G. New viral vector for superproduction of epitopes of vaccine proteins in plants. Acta Naturae 2011, 3, 73–82. [Google Scholar] [PubMed]
- Denis, J.; Acosta-Ramirez, E.; Zhao, Y.; Hamelin, M.E.; Koukavica, I.; Baz, M.; Abed, Y.; Savard, C.; Pare, C.; Lopez Macias, C.; et al. Development of a universal influenza A vaccine based on the M2e peptide fused to the papaya mosaic virus (PapMV) vaccine platform. Vaccine 2008, 26, 3395–3403. [Google Scholar] [CrossRef] [PubMed]
- Matic, S.; Rinaldi, R.; Masenga, V.; Noris, E. Efficient production of chimeric human papillomavirus 16 L1 protein bearing the M2e influenza epitope in Nicotiana benthamiana plants. BMC Biotechnol. 2011. [Google Scholar] [CrossRef]
- Ameiss, K.; Ashraf, S.; Kong, W.; Pekosz, A.; Wu, W.H.; Milich, D.; Billaud, J.N.; Curtiss, R., 3rd. Delivery of woodchuck hepatitis virus-like particle presented influenza M2e by recombinant attenuated Salmonella displaying a delayed lysis phenotype. Vaccine 2010, 28, 6704–6713. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, H.; Pouyanfard, S.; Bandehpour, M.; Noroozbabaei, Z.; Kazemi, B.; Saelens, X.; Mokhtari-Azad, T. Immunization with M2e-displaying T7 bacteriophage nanoparticles protects against influenza A virus challenge. PLOS ONE 2012, 7, e45765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bessa, J.; Schmitz, N.; Hinton, H.J.; Schwarz, K.; Jegerlehner, A.; Bachmann, M.F. Efficient induction of mucosal and systemic immune responses by virus-like particles administered intranasally: Implications for vaccine design. Eur. J. Immunol. 2008, 38, 114–126. [Google Scholar] [CrossRef] [PubMed]
- De Filette, M.; Martens, W.; Roose, K.; Deroo, T.; Vervalle, F.; Bentahir, M.; Vandekerckhove, J.; Fiers, W.; Saelens, X. An influenza A vaccine based on tetrameric ectodomain of matrix protein 2. J. Biol. Chem. 2008, 283, 11382–11387. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A.M.; Hakansson, K.O.; Jensen, B.A.; Christensen, D.; Andersen, P.; Thomsen, A.R.; Christensen, J.P. Increased immunogenicity and protective efficacy of influenza M2e fused to a tetramerizing protein. PLOS ONE 2012, 7, e46395. [Google Scholar] [CrossRef] [PubMed]
- Mozdzanowska, K.; Feng, J.; Eid, M.; Kragol, G.; Cudic, M.; Otvos, L., Jr.; Gerhard, W. Induction of influenza type A virus-specific resistance by immunization of mice with a synthetic multiple antigenic peptide vaccine that contains ectodomains of matrix protein 2. Vaccine 2003, 21, 2616–2626. [Google Scholar] [CrossRef] [PubMed]
- Mozdzanowska, K.; Zharikova, D.; Cudic, M.; Otvos, L.; Gerhard, W. Roles of adjuvant and route of vaccination in antibody response and protection engendered by a synthetic matrix protein 2-based influenza A virus vaccine in the mouse. Virol. J. 2007. [Google Scholar] [CrossRef]
- Zhao, G.; Lin, Y.; Du, L.; Guan, J.; Sun, S.; Sui, H.; Kou, Z.; Chan, C.C.; Guo, Y.; Jiang, S.; et al. An M2e-based multiple antigenic peptide vaccine protects mice from lethal challenge with divergent H5N1 influenza viruses. Virol. J. 2010. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Sun, S.; Du, L.; Xiao, W.; Ru, Z.; Kou, Z.; Guo, Y.; Yu, H.; Jiang, S.; Lone, Y.; et al. An H5N1 M2e-based multiple antigenic peptide vaccine confers heterosubtypic protection from lethal infection with pandemic 2009 H1N1 virus. Virol. J. 2010. [Google Scholar] [CrossRef]
- Liu, W.; Li, H.; Chen, Y.H. N-terminus of M2 protein could induce antibodies with inhibitory activity against influenza virus replication. FEMS Immunol. Med. Microbiol. 2003, 35, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Liang, X.; Horton, M.S.; Perry, H.C.; Citron, M.P.; Heidecker, G.J.; Fu, T.M.; Joyce, J.; Przysiecki, C.T.; Keller, P.M.; et al. Preclinical study of influenza virus A M2 peptide conjugate vaccines in mice, ferrets, and rhesus monkeys. Vaccine 2004, 22, 2993–3003. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.M.; Grimm, K.M.; Citron, M.P.; Freed, D.C.; Fan, J.; Keller, P.M.; Shiver, J.W.; Liang, X.; Joyce, J.G. Comparative immunogenicity evaluations of influenza A virus M2 peptide as recombinant virus like particle or conjugate vaccines in mice and monkeys. Vaccine 2009, 27, 1440–1447. [Google Scholar] [CrossRef] [PubMed]
- Turley, C.B.; Rupp, R.E.; Johnson, C.; Taylor, D.N.; Wolfson, J.; Tussey, L.; Kavita, U.; Stanberry, L.; Shaw, A. Safety and immunogenicity of a recombinant M2e-flagellin influenza vaccine (STF2.4xM2e) in healthy adults. Vaccine 2011, 29, 5145–5152. [Google Scholar] [CrossRef] [PubMed]
- Huleatt, J.W.; Nakaar, V.; Desai, P.; Huang, Y.; Hewitt, D.; Jacobs, A.; Tang, J.; McDonald, W.; Song, L.; Evans, R.K.; et al. Potent immunogenicity and efficacy of a universal influenza vaccine candidate comprising a recombinant fusion protein linking influenza M2e to the TLR5 ligand flagellin. Vaccine 2008, 26, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Talbot, H.K.; Rock, M.T.; Johnson, C.; Tussey, L.; Kavita, U.; Shanker, A.; Shaw, A.R.; Taylor, D.N. Immunopotentiation of trivalent influenza vaccine when given with VAX102, a recombinant influenza M2e vaccine fused to the TLR5 ligand flagellin. PLOS ONE 2010, 5, e14442. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, P.; Zylberman, V.; Ghersi, G.; Boado, L.; Palacios, C.; Goldbaum, F.; Mattion, N. Tandem repeats of the extracellular domain of Matrix 2 influenza protein exposed in Brucella lumazine synthase decameric carrier molecule induce protection in mice. Vaccine 2013, 31, 806–812. [Google Scholar] [CrossRef] [PubMed]
- Herve, P.L.; Raliou, M.; Bourdieu, C.; Dubuquoy, C.; Petit-Camurdan, A.; Bertho, N.; Eleouet, J.F.; Chevalier, C.; Riffault, S. A novel subnucleocapsid nanoplatform for mucosal vaccination against influenza virus that targets the ectodomain of matrix protein 2. J. Virol. 2014, 88, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Redding, L.; Weiner, D.B. DNA vaccines in veterinary use. Expert Rev. Vaccines 2009, 8, 1251–1276. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Lenz, S.D.; Thompson, D.H.; Pogranichniy, R.M. DNA-vaccine platform development against H1N1 subtype of swine influenza A viruses. Viral Immunol. 2012, 25, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yu, H.; Yang, F.R.; Huang, M.; Ma, J.H.; Tong, G.Z. Protective efficacy of a broadly cross-reactive swine influenza DNA vaccine encoding M2e, cytotoxic T lymphocyte epitope and consensus H3 hemagglutinin. Virol. J. 2012. [Google Scholar] [CrossRef]
- Park, K.S.; Seo, Y.B.; Lee, J.Y.; Im, S.J.; Seo, S.H.; Song, M.S.; Choi, Y.K.; Sung, Y.C. Complete protection against a H5N2 avian influenza virus by a DNA vaccine expressing a fusion protein of H1N1 HA and M2e. Vaccine 2011, 29, 5481–5487. [Google Scholar] [CrossRef] [PubMed]
- Lalor, P.A.; Webby, R.J.; Morrow, J.; Rusalov, D.; Kaslow, D.C.; Rolland, A.; Smith, L.R. Plasmid DNA-based vaccines protect mice and ferrets against lethal challenge with A/Vietnam/1203/04 (H5N1) influenza virus. J. Infect. Dis. 2008, 197, 1643–1652. [Google Scholar] [CrossRef] [PubMed]
- Okuda, K.; Ihata, A.; Watabe, S.; Okada, E.; Yamakawa, T.; Hamajima, K.; Yang, J.; Ishii, N.; Nakazawa, M.; Ohnari, K.; et al. Protective immunity against influenza A virus induced by immunization with DNA plasmid containing influenza M gene. Vaccine 2001, 19, 3681–3691. [Google Scholar] [CrossRef] [PubMed]
- Madan, V.; Castello, A.; Carrasco, L. Viroporins from RNA viruses induce caspase-dependent apoptosis. Cell Microbiol. 2008, 10, 437–451. [Google Scholar] [PubMed]
- Zhou, D.; Wu, T.L.; Emmer, K.L.; Kurupati, R.; Tuyishime, S.; Li, Y.; Giles-Davis, W.; Zhou, X.; Xiang, Z.; Liu, Q.; et al. Hexon-modified recombinant E1-deleted adenovirus vectors as dual specificity vaccine carriers for influenza virus. Mol. Ther. 2013, 21, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.H.; Park, H.J.; Han, G.Y.; Song, M.K.; Pereboev, A.; Hong, J.S.; Chang, J.; Byun, Y.H.; Seong, B.L.; Nguyen, H.H. Intranasal adenovirus-vectored vaccine for induction of long-lasting humoral immunity-mediated broad protection against influenza in mice. J. Virol. 2014, 88, 9693–9703. [Google Scholar] [CrossRef] [PubMed]
- Layton, S.L.; Kapczynski, D.R.; Higgins, S.; Higgins, J.; Wolfenden, A.D.; Liljebjelke, K.A.; Bottje, W.G.; Swayne, D.; Berghman, L.R.; Kwon, Y.M.; et al. Vaccination of chickens with recombinant Salmonella expressing M2e and CD154 epitopes increases protection and decreases viral shedding after low pathogenic avian influenza challenge. Poult. Sci. 2009, 88, 2244–2252. [Google Scholar] [CrossRef] [PubMed]
- Hessel, A.; Savidis-Dacho, H.; Coulibaly, S.; Portsmouth, D.; Kreil, T.R.; Crowe, B.A.; Schwendinger, M.G.; Pilz, A.; Barrett, P.N.; Falkner, F.G.; et al. MVA vectors expressing conserved influenza proteins protect mice against lethal challenge with H5N1, H9N2 and H7N1 viruses. PLOS ONE 2014, 9, e88340. [Google Scholar] [CrossRef] [PubMed]
- Okuno, Y.; Isegawa, Y.; Sasao, F.; Ueda, S. A common neutralizing epitope conserved between the hemagglutinins of influenza A virus H1 and H2 strains. J. Virol. 1993, 67, 2552–2558. [Google Scholar] [PubMed]
- DiLillo, D.J.; Tan, G.S.; Palese, P.; Ravetch, J.V. Broadly neutralizing hemagglutinin stalk-specific antibodies require FcgammaR interactions for protection against influenza virus in vivo. Nat. Med. 2014, 20, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Berendt, R.F. Simian model for the evaluation of immunity to influenza. Infect. Immun. 1974, 9, 101–105. [Google Scholar] [PubMed]
- Bodewes, R.; Rimmelzwaan, G.F.; Osterhaus, A.D. Animal models for the preclinical evaluation of candidate influenza vaccines. Expert Rev. Vaccines 2010, 9, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Van Riel, D.; Munster, V.J.; de Wit, E.; Rimmelzwaan, G.F.; Fouchier, R.A.; Osterhaus, A.D.; Kuiken, T. Human and avian influenza viruses target different cells in the lower respiratory tract of humans and other mammals. Am. J. Pathol. 2007, 171, 1215–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maher, J.A.; DeStefano, J. The ferret: An animal model to study influenza virus. Lab. Anim. (NY) 2004, 33, 50–53. [Google Scholar] [CrossRef]
- Ito, T.; Couceiro, J.N.; Kelm, S.; Baum, L.G.; Krauss, S.; Castrucci, M.R.; Donatelli, I.; Kida, H.; Paulson, J.C.; Webster, R.G.; et al. Molecular basis for the generation in pigs of influenza A viruses with pandemic potential. J. Virol. 1998, 72, 7367–7373. [Google Scholar] [PubMed]
- Ma, W.; Kahn, R.E.; Richt, J.A. The pig as a mixing vessel for influenza viruses: Human and veterinary implications. J. Mol. Genet. Med. 2008, 3, 158–166. [Google Scholar] [PubMed]
- Heinen, P.P.; Rijsewijk, F.A.; de Boer-Luijtze, E.A.; Bianchi, A.T. Vaccination of pigs with a DNA construct expressing an influenza virus M2-nucleoprotein fusion protein exacerbates disease after challenge with influenza A virus. J. Gen. Virol. 2002, 83, 1851–1859. [Google Scholar] [PubMed]
- Reese, K.A.; Lupfer, C.; Johnson, R.C.; Mitev, G.M.; Mullen, V.M.; Geller, B.L.; Pastey, M. A novel lactococcal vaccine expressing a peptide from the M2 antigen of H5N2 highly pathogenic avian Influenza A virus prolongs survival of vaccinated chickens. Vet. Med. Int. 2013. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, M.; Liu, C.; Du, J.; Shi, W.; Sun, E.; Li, H.; Li, J.; Zhang, Y. Vaccination with different M2e epitope densities confers partial protection against H5N1 influenza A virus challenge in chickens. Intervirology 2011, 54, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Zebedee, S.L.; Lamb, R.A. Influenza A virus M2 protein: Monoclonal antibody restriction of virus growth and detection of M2 in virions. J. Virol. 1988, 62, 2762–2772. [Google Scholar] [PubMed]
- Treanor, J.J.; Tierney, E.L.; Zebedee, S.L.; Lamb, R.A.; Murphy, B.R. Passively transferred monoclonal antibody to the M2 protein inhibits influenza A virus replication in mice. J. Virol. 1990, 64, 1375–1377. [Google Scholar] [PubMed]
- Zharikova, D.; Mozdzanowska, K.; Feng, J.; Zhang, M.; Gerhard, W. Influenza type A virus escape mutants emerge in vivo in the presence of antibodies to the ectodomain of matrix protein 2. J. Virol. 2005, 79, 6644–6654. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zou, P.; Chen, Y.H. Monoclonal antibodies recognizing EVETPIRN epitope of influenza A virus M2 protein could protect mice from lethal influenza A virus challenge. Immunol. Lett. 2004, 93, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Grandea, A.G., 3rd; Olsen, O.A.; Cox, T.C.; Renshaw, M.; Hammond, P.W.; Chan-Hui, P.Y.; Mitcham, J.L.; Cieplak, W.; Stewart, S.M.; Grantham, M.L.; et al. Human antibodies reveal a protective epitope that is highly conserved among human and nonhuman influenza A viruses. Proc. Natl. Acad. Sci. USA 2010, 107, 12658–12663. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.M.; Freed, D.C.; Horton, M.S.; Fan, J.; Citron, M.P.; Joyce, J.G.; Garsky, V.M.; Casimiro, D.R.; Zhao, Q.; Shiver, J.W.; et al. Characterizations of four monoclonal antibodies against M2 protein ectodomain of influenza A virus. Virology 2009, 385, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Ernst, W.A.; Kim, H.J.; Tumpey, T.M.; Jansen, A.D.; Tai, W.; Cramer, D.V.; Adler-Moore, J.P.; Fujii, G. Protection against H1, H5, H6 and H9 influenza A infection with liposomal matrix 2 epitope vaccines. Vaccine 2006, 24, 5158–5168. [Google Scholar] [CrossRef] [PubMed]
- Tompkins, S.M.; Zhao, Z.S.; Lo, C.Y.; Misplon, J.A.; Liu, T.; Ye, Z.; Hogan, R.J.; Wu, Z.; Benton, K.A.; Tumpey, T.M.; et al. Matrix protein 2 vaccination and protection against influenza viruses, including subtype H5N1. Emerg. Infect. Dis. 2007, 13, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Jegerlehner, A.; Schmitz, N.; Storni, T.; Bachmann, M.F. Influenza A vaccine based on the extracellular domain of M2: Weak protection mediated via antibody-dependent NK cell activity. J. Immunol. 2004, 172, 5598–5605. [Google Scholar] [CrossRef] [PubMed]
- El Bakkouri, K.; Descamps, F.; de Filette, M.; Smet, A.; Festjens, E.; Birkett, A.; van Rooijen, N.; Verbeek, S.; Fiers, W.; Saelens, X. Universal vaccine based on ectodomain of matrix protein 2 of influenza A: Fc receptors and alveolar macrophages mediate protection. J. Immunol. 2011, 186, 1022–1031. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Song, A.; Levin, J.; Dennis, D.; Zhang, N.J.; Yoshida, H.; Koriazova, L.; Madura, L.; Shapiro, L.; Matsumoto, A.; et al. Therapeutic potential of a fully human monoclonal antibody against influenza A virus M2 protein. Antiviral Res. 2008, 80, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Huang, J.H.; Yuan, X.Y.; Huang, W.S.; Chen, Y.H. Characterization of immunity induced by M2e of influenza virus. Vaccine 2007, 25, 8868–8873. [Google Scholar] [CrossRef] [PubMed]
- Markine-Goriaynoff, D.; Coutelier, J.P. Increased efficacy of the immunoglobulin G2a subclass in antibody-mediated protection against lactate dehydrogenase-elevating virus-induced polioencephalomyelitis revealed with switch mutants. J. Virol. 2002, 76, 432–435. [Google Scholar] [CrossRef] [PubMed]
- De Filette, M.; Ramne, A.; Birkett, A.; Lycke, N.; Lowenadler, B.; Min Jou, W.; Saelens, X.; Fiers, W. The universal influenza vaccine M2e-HBc administered intranasally in combination with the adjuvant CTA1-DD provides complete protection. Vaccine 2006, 24, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Guilliams, M.; Bruhns, P.; Saeys, Y.; Hammad, H.; Lambrecht, B.N. The function of Fcgamma receptors in dendritic cells and macrophages. Nat. Rev. Immunol. 2014, 14, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Purzel, J.; Schmitt, R.; Viertlboeck, B.C.; Gobel, T.W. Chicken IgY binds its receptor at the CH3/CH4 interface similarly as the human IgA: Fc alpha RI interaction. J. Immunol. 2009, 183, 4554–4559. [Google Scholar] [CrossRef] [PubMed]
- Viertlboeck, B.C.; Schweinsberg, S.; Hanczaruk, M.A.; Schmitt, R.; du Pasquier, L.; Herberg, F.W.; Gobel, T.W. The chicken leukocyte receptor complex encodes a primordial, activating, high-affinity IgY Fc receptor. Proc. Natl. Acad. Sci. USA 2007, 104, 11718–11723. [Google Scholar] [CrossRef] [PubMed]
- Magor, K.E.; Higgins, D.A.; Middleton, D.L.; Warr, G.W. One gene encodes the heavy chains for three different forms of IgY in the duck. J. Immunol. 1994, 153, 5549–5555. [Google Scholar] [PubMed]
- Ramos, E.L.; Mitcham, J.L.; Koller, T.D.; Bonavia, A.; Usner, D.W.; Balaratnam, G.; Fredlund, P.; Swiderek, K.M. Efficacy and safety of treatment with an anti-M2e monoclonal antibody in experimental human influenza. J. Infect. Dis. 2014. [Google Scholar] [CrossRef]
- Song, A.; Myojo, K.; Laudenslager, J.; Harada, D.; Miura, T.; Suzuki, K.; Kuni-Kamochi, R.; Soloff, R.; Ohgami, K.; Kanda, Y. Evaluation of a fully human monoclonal antibody against multiple influenza A viral strains in mice and a pandemic H1N1 strain in nonhuman primates. Antiviral Res. 2014, 111C, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Schotsaert, M.; Ysenbaert, T.; Neyt, K.; Ibanez, L.I.; Bogaert, P.; Schepens, B.; Lambrecht, B.N.; Fiers, W.; Saelens, X. Natural and long-lasting cellular immune responses against influenza in the M2e-immune host. Mucosal. Immunol. 2013, 6, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Bodewes, R.; Kreijtz, J.H.; Rimmelzwaan, G.F. Yearly influenza vaccinations: A double-edged sword? Lancet Infect. Dis. 2009, 9, 784–788. [Google Scholar] [CrossRef] [PubMed]
- CDC. Prevention and control of seasonal influenza with vaccines. Recommendations of the Advisory Committee on Immunization Practices—United States, 2013–2014. In MMWR Recomm. Rep.; 2013; 62, pp. 1–43. [Google Scholar]
- Bodewes, R.; Kreijtz, J.H.; Baas, C.; Geelhoed-Mieras, M.M.; de Mutsert, G.; van Amerongen, G.; van den Brand, J.M.; Fouchier, R.A.; Osterhaus, A.D.; Rimmelzwaan, G.F. Vaccination against human influenza A/H3N2 virus prevents the induction of heterosubtypic immunity against lethal infection with avian influenza A/H5N1 virus. PLOS ONE 2009, 4, e5538. [Google Scholar] [CrossRef] [PubMed]
- Bodewes, R.; Kreijtz, J.H.; Geelhoed-Mieras, M.M.; van Amerongen, G.; Verburgh, R.J.; van Trierum, S.E.; Kuiken, T.; Fouchier, R.A.; Osterhaus, A.D.; Rimmelzwaan, G.F. Vaccination against seasonal influenza A/H3N2 virus reduces the induction of heterosubtypic immunity against influenza A/H5N1 virus infection in ferrets. J. Virol. 2011, 85, 2695–2702. [Google Scholar] [CrossRef] [PubMed]
- Bodewes, R.; Kreijtz, J.H.; Hillaire, M.L.; Geelhoed-Mieras, M.M.; Fouchier, R.A.; Osterhaus, A.D.; Rimmelzwaan, G.F. Vaccination with whole inactivated virus vaccine affects the induction of heterosubtypic immunity against influenza virus A/H5N1 and immunodominance of virus-specific CD8+ T-cell responses in mice. J. Gen. Virol. 2010, 91, 1743–1753. [Google Scholar] [CrossRef] [PubMed]
- Bodewes, R.; Fraaij, P.L.; Geelhoed-Mieras, M.M.; van Baalen, C.A.; Tiddens, H.A.; van Rossum, A.M.; van der Klis, F.R.; Fouchier, R.A.; Osterhaus, A.D.; Rimmelzwaan, G.F. Annual vaccination against influenza virus hampers development of virus-specific CD8+ T cell immunity in children. J. Virol. 2011, 85, 11995–12000. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Peng, Z.; Liu, Z.; Lu, Y.; Ding, J.; Chen, Y.H. High epitope density in a single recombinant protein molecule of the extracellular domain of influenza A virus M2 protein significantly enhances protective immunity. Vaccine 2004, 23, 366–371. [Google Scholar] [CrossRef] [PubMed]
- De Filette, M.; Fiers, W.; Martens, W.; Birkett, A.; Ramne, A.; Lowenadler, B.; Lycke, N.; Jou, W.M.; Saelens, X. Improved design and intranasal delivery of an M2e-based human influenza A vaccine. Vaccine 2006, 24, 6597–6601. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Chen, Y.H. High epitope density in a single protein molecule significantly enhances antigenicity as well as immunogenicity: A novel strategy for modern vaccine development and a preliminary investigation about B cell discrimination of monomeric proteins. Eur. J. Immunol. 2005, 35, 505–514. [Google Scholar] [CrossRef] [PubMed]
- De Filette, M.; Min Jou, W.; Birkett, A.; Lyons, K.; Schultz, B.; Tonkyro, A.; Resch, S.; Fiers, W. Universal influenza A vaccine: Optimization of M2-based constructs. Virology 2005, 337, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Sendi, P.; Locher, R.; Bucheli, B.; Battegay, M. Intranasal influenza vaccine in a working population. Clin. Infect. Dis. 2004, 38, 974–980. [Google Scholar] [CrossRef] [PubMed]
- Dynavax reports new phase 1a and phase 1b data for universal flu vaccine candidate. Available online: http://investors.dynavax.com/releasedetail.cfm?ReleaseID=551606&refdoi=10.1186/1471–2458–13-s3-s14 (accessed on 22 February 2011).
- Fiers, W.; de Filette, M.; el Bakkouri, K.; Schepens, B.; Roose, K.; Schotsaert, M.; Birkett, A.; Saelens, X. M2e-based universal influenza A vaccine. Vaccine 2009, 27, 6280–6283. [Google Scholar] [CrossRef] [PubMed]
- Safety study of recombinant M2e influenza-A vaccine in healthy adults (FLU-A). Available online: https://clinicaltrials.gov/ct2/show/NCT00819013?term=Acam-flu-A&rank=1 (accessed on 10 October 2008).
- Smith, L.R.; Wloch, M.K.; Ye, M.; Reyes, L.R.; Boutsaboualoy, S.; Dunne, C.E.; Chaplin, J.A.; Rusalov, D.; Rolland, A.P.; Fisher, C.L.; et al. Phase 1 clinical trials of the safety and immunogenicity of adjuvanted plasmid DNA vaccines encoding influenza A virus H5 hemagglutinin. Vaccine 2010, 28, 2565–2572. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, L.; Cho, K.J.; Fiers, W.; Saelens, X. M2e-Based Universal Influenza A Vaccines. Vaccines 2015, 3, 105-136. https://doi.org/10.3390/vaccines3010105
Deng L, Cho KJ, Fiers W, Saelens X. M2e-Based Universal Influenza A Vaccines. Vaccines. 2015; 3(1):105-136. https://doi.org/10.3390/vaccines3010105
Chicago/Turabian StyleDeng, Lei, Ki Joon Cho, Walter Fiers, and Xavier Saelens. 2015. "M2e-Based Universal Influenza A Vaccines" Vaccines 3, no. 1: 105-136. https://doi.org/10.3390/vaccines3010105