Proteomic Analysis of Antigen 60 Complex of M. bovis Bacillus Calmette-Guérin Reveals Presence of Extracellular Vesicle Proteins and Predicted Functional Interactions

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Trypsin Digestion
2.2. Mass Spectrometry (MS)
2.3. Seroimmunological Analysis
2.3.1. Serum Sample Population
2.3.2. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE) and Western Blot (WB)
2.4. Data Analysis
3. Results
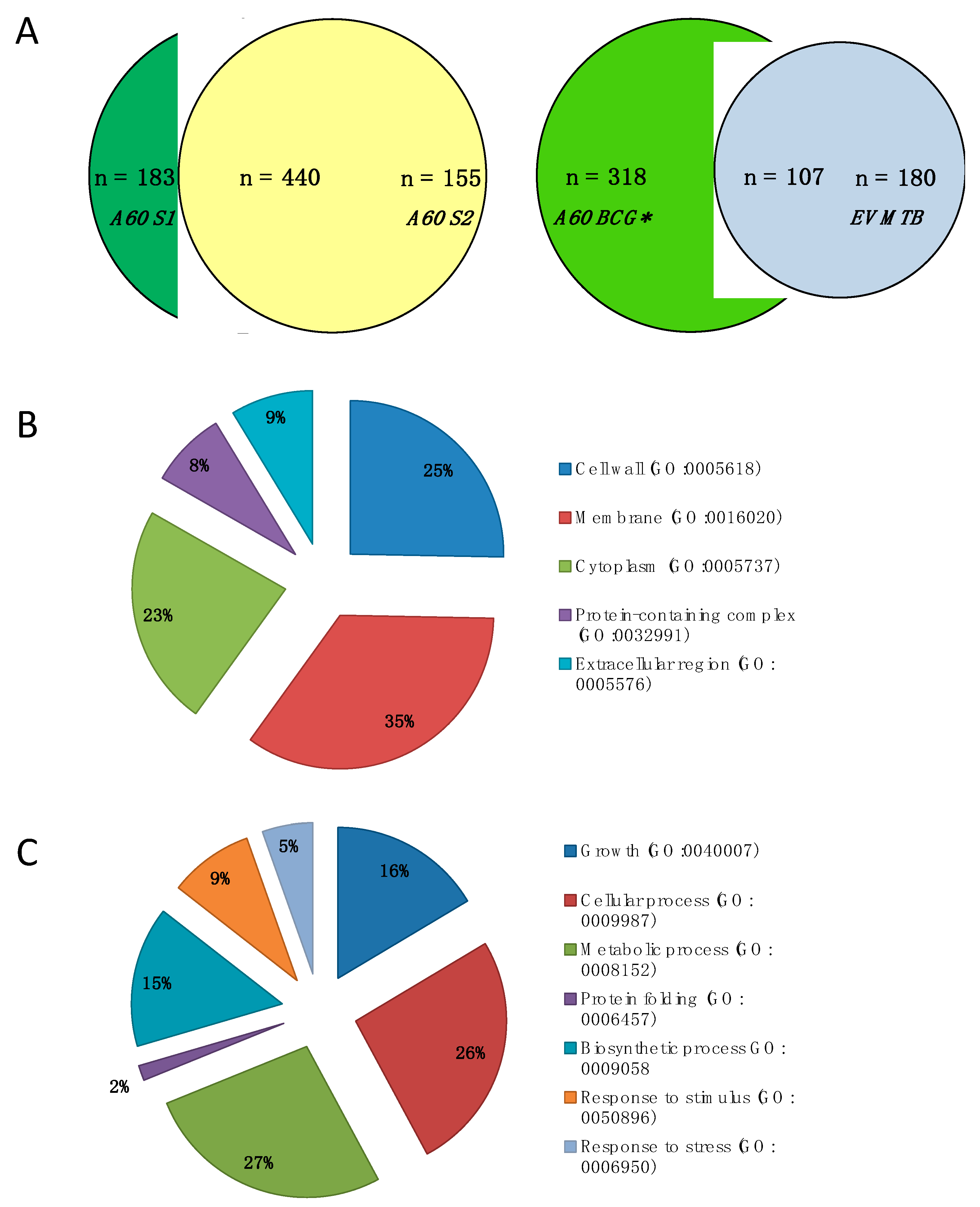
3.1. Protein Identification and Annotation
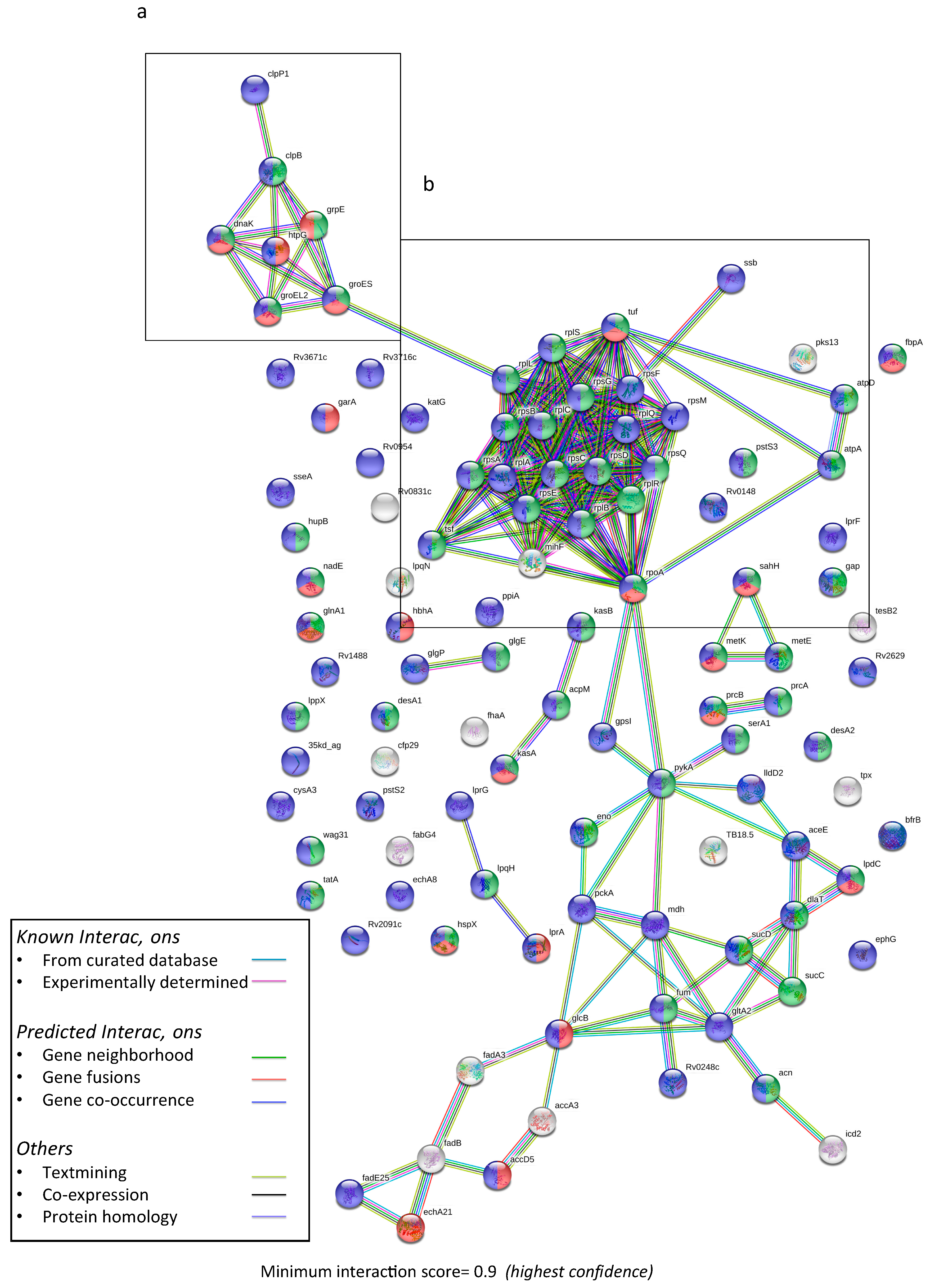
3.2. Predictive Protein–Protein Interaction
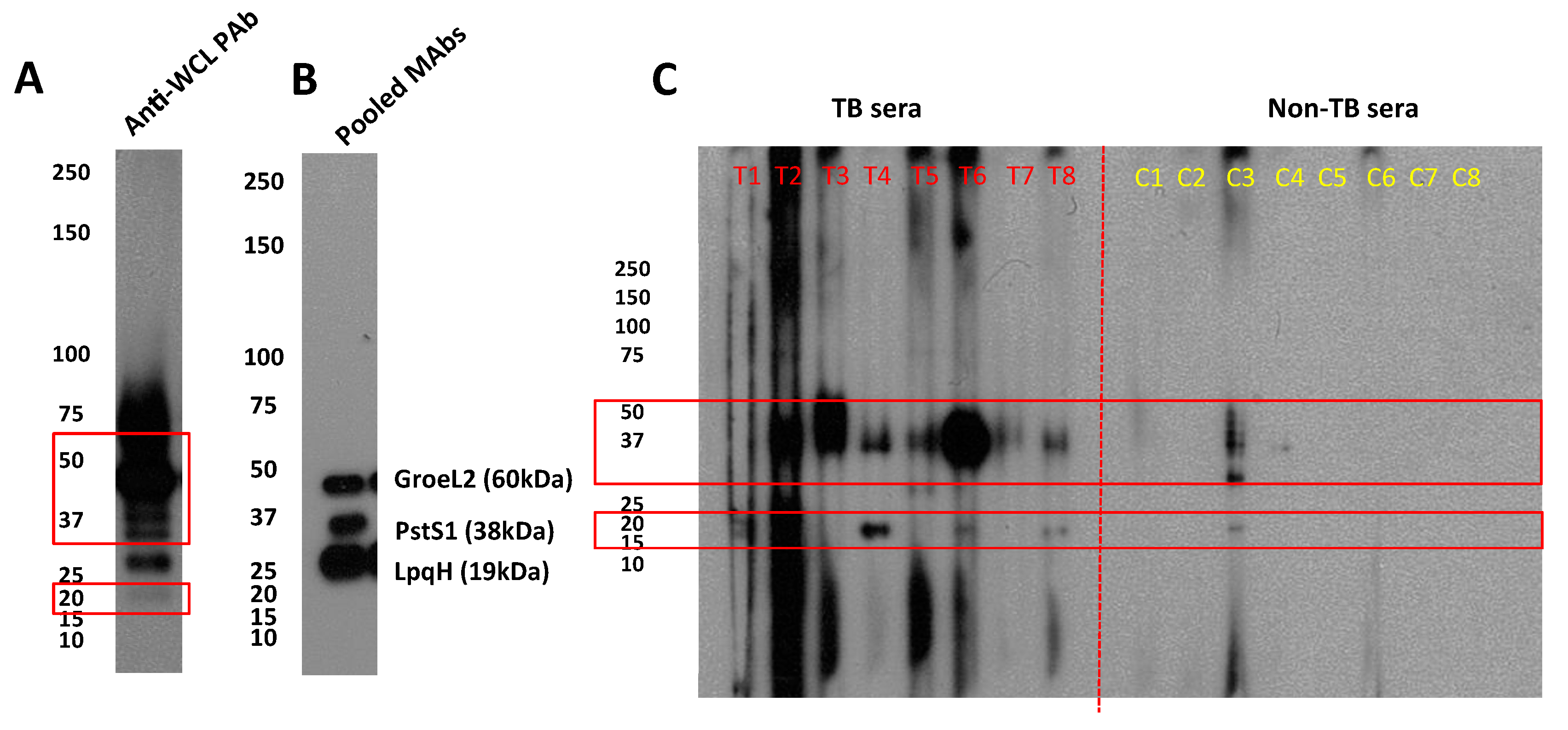
3.3. Antigenicity of A60-BCG
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Global Tuberculosis Report 2018; World Health Organization: Geneva, Switzerland, 2018.
- Riojas, M.A.; McGough, K.J.; Rider-Riojas, C.J.; Rastogi, N.; Hazbon, M.H. Phylogenomic analysis of the species of the mycobacterium tuberculosis complex demonstrates that Mycobacterium africanum, Mycobacterium bovis, Mycobacterium caprae, Mycobacterium microti and Mycobacterium pinnipedii are later heterotypic synonyms of Mycobacterium tuberculosis. Int. J. Syst. Evol. Microbiol. 2018, 68, 324–332. [Google Scholar] [PubMed]
- Brewer, T.F.; Colditz, G.A. Relationship between Bacille Calmette-Guerin (BCG) strains and the efficacy of BCG vaccine in the prevention of tuberculosis. Clin. Infect. Dis. 1995, 20, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Dockrell, H.M.; Smith, S.G. What have we learnt about BCG vaccination in the last 20 years? Front. Immunol. 2017, 8, 1134. [Google Scholar] [CrossRef] [PubMed]
- Mohd Hanafiah, K.; Liu, J.J.; Lieschke, K.; Barnes, N.C.; Garcia, M.L.; Anderson, D.A. Serological biomarker screening and host factor analysis elucidating immune response heterogeneity in active pulmonary tuberculosis. Trop. Biomed. 2017, 34, 556–569. [Google Scholar]
- Sridhar, S.; Pollock, K.; Lalvani, A. Redefining latent tuberculosis. Future Microbiol. 2011, 6, 1021–1035. [Google Scholar] [CrossRef] [PubMed]
- Mohd Hanafiah, K.G.; Garcia, M.L.; Anderson, D.A. An observational case-control study to determine human immunodeficiency virus and host factor influence on biomarker distribution and serodiagnostic potential in adult pulmonary tuberculosis. Trop. Med. Infect. Dis. 2019, 4, 57. [Google Scholar] [CrossRef] [PubMed]
- Kik, S.V.; Denkinger, C.M.; Casenghi, M.; Vadnais, C.; Pai, M. Tuberculosis diagnostics: Which target product profiles should be prioritised? Eur. Res. J. 2014, 44, 537–540. [Google Scholar] [CrossRef]
- Sotgiu, G.; Centis, R.; D’Ambrosio, L.; Migliori, G.B. Tuberculosis treatment and drug regimens. Cold Spring Harb. Perspect. Med. 2015, 5, a017822. [Google Scholar] [CrossRef]
- Khan, M.S.; Fletcher, H. London School of Hygiene and Tropical Medicine TB Centre Steering Committee; Coker, R. Investments in tuberculosis research—What are the gaps? BMC Med. 2016, 14, 123. [Google Scholar]
- Schrager, L.K.; Harris, R.C.; Vekemans, J. Research and development of new tuberculosis vaccines: A review. F1000Res 2018, 7, 1732. [Google Scholar] [CrossRef]
- Ernst, J.D. Mechanisms of m. Tuberculosis immune evasion as challenges to TB vaccine design. Cell Host Microbe 2018, 24, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Closs, O.; Harboe, M.; Axelsen, N.H.; Bunch-Christensen, K.; Magnusson, M. The antigens of Mycobacterium bovis, strain BCG, studied by crossed immunoelectrophoresis: A reference system. Scand. J. Immunol. 1980, 12, 249–263. [Google Scholar] [CrossRef] [PubMed]
- Harboe, M.; Mshana, R.N.; Closs, O.; Kronvall, G.; Axelsen, N.H. Cross-reactions between mycobacteria. Ii. Crossed immunoelectrophoretic analysis of soluble antigens of BCG and comparison with other mycobacteria. Scand. J. Immunol. 1979, 9, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Cocito, C.; Vanlinden, F. Composition and immunoreactivity of the A60 complex and other cell fractions from Mycobacterium bovis bcg. Scand. J. Immunol. 1995, 41, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Goudarzi, H.; Kazemi, B.; Falah, F.; Eslami, G.; Ghazi, M.; Doustar, F. Purification and molecular analysis of BCG antigen 60. Med. J. Islam. Repub. Iran 2003, 17, 147–152. [Google Scholar]
- Maes, R. Comparison of a60 and three glycolipid antigens in an ELISA test for tuberculosis. Clin. Microbiol. Infect. 1997, 3, 595–596. [Google Scholar] [CrossRef]
- Steingart, K.R.; Henry, M.; Laal, S.; Hopewell, P.C.; Ramsay, A.; Menzies, D.; Cunningham, J.; Weldingh, K.; Pai, M. Commercial serological antibody detection tests for the diagnosis of pulmonary tuberculosis: A systematic review. PLoS Med. 2007, 4, 1041–1060. [Google Scholar] [CrossRef]
- Abebe, F.; Holm-Hansen, C.; Wiker, H.G.; Bjune, G. Progress in serodiagnosis of mycobacterium tuberculosis infection. Scand. J. Immunol. 2007, 66, 176–191. [Google Scholar] [CrossRef]
- Hubbard, R.D.; Flory, C.M.; Collins, F.M.; Cocito, C. Immunization of mice with the antigen A60 of Mycobacterium bovis BCG. Clin. Exp. Immunol. 1992, 88, 129–131. [Google Scholar] [CrossRef]
- Coetsier, C.; Baelden, M.C.; Coene, M.; Cocito, C. Immunological analysis of the components of the antigen complex A60 of Mycobacterium bovis BCG. Clin. Diagn. Lab. Immunol. 1994, 1, 139–144. [Google Scholar]
- Cocito, C.; Vanlinden, F. Preparation and properties of antigen 60 from Mycobacterium bovis BCG. Clin. Exp. Immunol. 1986, 66, 262–272. [Google Scholar] [PubMed]
- Fabre, I.; L’Homme, O.; Bruneteau, M.; Michel, G.; Cocito, C. Chemical composition of antigen 60 from Mycobacterium bovis BCG. Scand. J. Immunol. 1986, 24, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Harboe, M. Antigens of PPD, old tuberculin, and autoclaved Mycobacterium bovis BCG studied by crossed immunoelectrophoresis. Am. Rev. Respir. Dis. 1981, 124, 80–87. [Google Scholar] [PubMed]
- Cocito, C.; Baelden, M.C.; Benoit, C. Immunological properties of Antigen 60 of BCG. Induction of humoral and cellular immune reactions. Scand. J. Immunol. 1987, 25, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Rodriguez, G.M. Mycobacterial extracellular vesicles and host pathogen interactions. Pathog. Dis. 2018, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Athman, J.J.; Sande, O.J.; Groft, S.G.; Reba, S.M.; Nagy, N.; Wearsch, P.A.; Richardson, E.T.; Rojas, R.; Boom, W.H.; Shukla, S.; et al. Mycobacterium tuberculosis membrane vesicles inhibit t cell activation. J. Immunol. 2017, 198, 2028–2037. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.; Wolf, J.M.; Prados-Rosales, R.; Casadevall, A. Through the wall: Extracellular vesicles in gram-positive bacteria, mycobacteria and fungi. Nat. Rev. Microbiol. 2015, 13, 620–630. [Google Scholar] [CrossRef]
- Prados-Rosales, R.; Baena, A.; Martinez, L.R.; Luque-Garcia, J.; Kalscheuer, R.; Veeraraghavan, U.; Camara, C.; Nosanchuk, J.D.; Besra, G.S.; Chen, B.; et al. Mycobacteria release active membrane vesicles that modulate immune responses in a TLR2-dependent manner in mice. J. Clin. Investig. 2011, 121, 1471–1483. [Google Scholar] [CrossRef]
- Drage, M.G.; Pecora, N.D.; Hise, A.G.; Febbraio, M.; Silverstein, R.L.; Golenbock, D.T.; Boom, W.H.; Harding, C.V. TLR2 and its co-receptors determine responses of macrophages and dendritic cells to lipoproteins of mycobacterium tuberculosis. Cell. Immunol. 2009, 258, 29–37. [Google Scholar] [CrossRef]
- Palma, C.; Spallek, R.; Piccaro, G.; Pardini, M.; Jonas, F.; Oehlmann, W.; Singh, M.; Cassone, A. Tuberculosis phosphate-binding lipoproteins PstS1 and PstS3 induce TH1 and TH17 responses that are not associated with protection against m. Tuberculosis infection. Clin. Dev. Immunol. 2011, 2011, 690328. [Google Scholar] [CrossRef]
- Athman, J.J.; Wang, Y.; McDonald, D.J.; Boom, W.H.; Harding, C.V.; Wearsch, P.A. Bacterial membrane vesicles mediate the release of Mycobacterium tuberculosis lipoglycans and lipoproteins from infected macrophages. J. Immunol. 2015, 195, 1044–1053. [Google Scholar] [CrossRef] [PubMed]
- Stutz, M.D.; Clark, M.P.; Doerflinger, M.; Pellegrini, M. Mycobacterium tuberculosis: Rewiring host cell signaling to promote infection. J. Leukoc. Biol. 2018, 103, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Maes, R.F. Isolation of Mycobacterial a 60 Antigen for Diagnostic Purposes. U.S. Patent 4,777,130, 11 October 1988. Available online: https://patents.google.com/patent/US4777130A/en (accessed on 2 August 2019).
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. String v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- Bioinformatics and Evolutionary Genomics—Calculate and Draw Custom Venn Diagrams. Available online: http://bioinformatics.psb.ugent.be/webtools/Venn/ (accessed on 10 June 2018).
- Lee, J.; Kim, S.H.; Choi, D.S.; Lee, J.S.; Kim, D.K.; Go, G.; Park, S.M.; Kim, S.H.; Shin, J.H.; Chang, C.L.; et al. Proteomic analysis of extracellular vesicles derived from mycobacterium tuberculosis. Proteomics 2015, 15, 3331–3337. [Google Scholar] [CrossRef] [PubMed]
- Roperto, S.; Varano, M.; Russo, V.; Luca, R.; Cagiola, M.; Gaspari, M.; Ceccarelli, D.M.; Cuda, G.; Roperto, F. Proteomic analysis of protein purified derivative of mycobacterium bovis. J. Transl. Med. 2017, 15, 68. [Google Scholar] [CrossRef] [PubMed]
- Prasad, T.S.; Verma, R.; Kumar, S.; Nirujogi, R.S.; Sathe, G.J.; Madugundu, A.K.; Sharma, J.; Puttamallesh, V.N.; Ganjiwale, A.; Myneedu, V.P.; et al. Proteomic analysis of purified protein derivative of mycobacterium tuberculosis. Clin. Proteom. 2013, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Brosch, R.; Gordon, S.V.; Garnier, T.; Eiglmeier, K.; Frigui, W.; Valenti, P.; Dos Santos, S.; Duthoy, S.; Lacroix, C.; Garcia-Pelayo, C.; et al. Genome plasticity of bcg and impact on vaccine efficacy. Proc. Natl. Acad. Sci. USA 2007, 104, 5596–5601. [Google Scholar] [CrossRef]
- Yang, H.; Kruh-Garcia, N.A.; Dobos, K.M. Purified protein derivatives of tuberculin--past, present, and future. FEMS Immunol. Med. Microbiol. 2012, 66, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Gcebe, N.; Michel, A.; Gey van Pittius, N.C.; Rutten, V. Comparative genomics and proteomic analysis of four non-tuberculous mycobacterium species and Mycobacterium tuberculosis complex: Occurrence of shared immunogenic proteins. Front. Microbiol. 2016, 7, 795. [Google Scholar] [CrossRef]
- He, Z.; De Buck, J. Localization of proteins in the cell wall of Mycobacterium avium subsp. Paratuberculosis k10 by proteomic analysis. Proteome Sci. 2010, 8, 21. [Google Scholar] [CrossRef]
- Wolfe, L.M.; Mahaffey, S.B.; Kruh, N.A.; Dobos, K.M. Proteomic definition of the cell wall of Mycobacterium tuberculosis. J. Proteome Res. 2010, 9, 5816–5826. [Google Scholar] [CrossRef]
- Meneses, N.; Mendoza-Hernandez, G.; Encarnacion, S. The extracellular proteome of rhizobium etli CE3 in exponential and stationary growth phase. Proteome Sci. 2010, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Prados-Rosales, R.; Weinrick, B.C.; Pique, D.G.; Jacobs, W.R., Jr.; Casadevall, A.; Rodriguez, G.M. Role for mycobacterium tuberculosis membrane vesicles in iron acquisition. J. Bacteriol. 2014, 196, 1250–1256. [Google Scholar] [CrossRef]
- Parra, J.; Marcoux, J.; Poncin, I.; Canaan, S.; Herrmann, J.L.; Nigou, J.; Burlet-Schiltz, O.; Riviere, M. Scrutiny of Mycobacterium tuberculosis 19 kda antigen proteoforms provides new insights in the lipoglycoprotein biogenesis paradigm. Sci. Rep. 2017, 7, 43682. [Google Scholar] [CrossRef]
- Kim, W.S.; Jung, I.D.; Kim, J.S.; Kim, H.M.; Kwon, K.W.; Park, Y.M.; Shin, S.J. Mycobacterium tuberculosis GrpE, a heat-shock stress responsive chaperone, promotes Th1-biased T cell immune response via Tlr4-mediated activation of dendritic cells. Front. Cell. Infect. Microbiol. 2018, 8, 95. [Google Scholar] [CrossRef]
- Kim, W.S.; Kim, J.S.; Kim, H.M.; Kwon, K.W.; Eum, S.Y.; Shin, S.J. Comparison of immunogenicity and vaccine efficacy between heat-shock proteins, Hsp70 and GrpE, in the DnaK operon of Mycobacterium tuberculosis. Sci. Rep. 2018, 8, 14411. [Google Scholar] [CrossRef]
- Jeng, W.; Lee, S.; Sung, N.; Lee, J.; Tsai, F.T. Molecular chaperones: Guardians of the proteome in normal and disease states. F1000Res 2015. [Google Scholar] [CrossRef] [PubMed]
- Leibly, D.J.; Nguyen, T.N.; Kao, L.T.; Hewitt, S.N.; Barrett, L.K.; Van Voorhis, W.C. Stabilizing additives added during cell lysis aid in the solubilization of recombinant proteins. PLoS ONE 2012, 7, e52482. [Google Scholar] [CrossRef]
- Cho, Y.S.; Jang, Y.B.; Lee, S.E.; Cho, J.Y.; Ahn, J.M.; Hwang, I.; Heo, E.; Nam, H.M.; Cho, D.; Her, M.; et al. Short communication: Proteomic characterization of tuberculin purified protein derivative from Mycobacterium bovis. Res. Vet. Sci. 2015, 101, 117–119. [Google Scholar] [CrossRef]
- Jamieson, F.B.; Guthrie, J.L.; Neemuchwala, A.; Lastovetska, O.; Melano, R.G.; Mehaffy, C. Profiling of Rpob mutations and MICs for rifampin and rifabutin in Mycobacterium tuberculosis. J. Clin. Microbiol. 2014, 52, 2157–2162. [Google Scholar] [CrossRef]
- Singh, A.; Somvanshi, P.; Grover, A. Pyrazinamide drug resistance in rpsa mutant (438a) of mycobacterium tuberculosis: Dynamics of essential motions and free-energy landscape analysis. J. Cell. Biochem. 2018. [Google Scholar] [CrossRef]
- White, D.W.; Elliott, S.R.; Odean, E.; Bemis, L.T.; Tischler, A.D. Mycobacterium tuberculosis pst/senx3-regx3 regulates membrane vesicle production independently of esx-5 activity. MBio 2018, 9. [Google Scholar] [CrossRef]
- Martinot, A.J.; Farrow, M.; Bai, L.; Layre, E.; Cheng, T.Y.; Tsai, J.H.; Iqbal, J.; Annand, J.W.; Sullivan, Z.A.; Hussain, M.M.; et al. Mycobacterial metabolic syndrome: Lprg and rv1410 regulate triacylglyceride levels, growth rate and virulence in Mycobacterium tuberculosis. PLoS Pathog. 2016, 12, e1005351. [Google Scholar] [CrossRef]
- Doddam, S.N.; Peddireddy, V.; Ahmed, N. Mycobacterium tuberculosis Dosr regulon gene rv2004c encodes a novel antigen with pro-inflammatory functions and potential diagnostic application for detection of latent tuberculosis. Front. Immunol. 2017, 8, 712. [Google Scholar] [CrossRef]
- Collins, M.E.; Patki, A.; Wall, S.; Nolan, A.; Goodger, J.; Woodward, M.J.; Dale, J.W. Cloning and characterization of the gene for the ‘19 kda’ antigen of mycobacterium bovis. J. Gen. Microbiol. 1990, 136, 1429–1436. [Google Scholar] [CrossRef]
- Gopinath, K.; Singh, S. Non-tuberculous mycobacteria in tb-endemic countries: Are we neglecting the danger? PLoS Negl. Trop. Dis. 2010, 4, e615. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| No. | Protein Names | Gene Names | Length | Mass (Da) | Biological Process | Cellular Component | Peptide/Proteinin | |
|---|---|---|---|---|---|---|---|---|
| Sig. Matches | Cover (%) | |||||||
| 1 | Chaperone protein DnaK | dnaK BCG_0389 | 625 | 66790 | cellular response to stress, protein folding | bacterial extracellular vesicle | 58 | 43.75 |
| 2 | Probable fatty acid synthase (Fas) | fas BCG_2545c | 3069 | 326790 | metabolic process | cell wall, cytoplasm, plasma membrane | 55 | 17.25 |
| 3 | Chaperone protein ClpB | clpB BCG_0422c | 848 | 92512 | cellular response to stress, protein folding; metabolic process | cell wall, cytoplasm, plasma membrane | 50.5 | 36.65 |
| 4 | DNA-directed RNA polymerase subunit beta (RpoC) | rpoC BCG_0717 | 1316 | 147303 | transcription | cell wall, cytoplasm, plasma membrane | 46.5 | 33.35 |
| 5 | Polyketide synthase (Pks13) | pks13 BCG_3862c | 1733 | 186629 | biosynthetic process | NA | 46 | 21.7 |
| 6 | Probable multifunctional mycocerosic acid synthase (Mas) | mas BCG_2962c | 2111 | 225568 | NA | NA | 45.5 | 49.1 |
| 7 | Cell wall synthesis protein Wag31 | wag31 BCG_2162c | 260 | 28260 | cell wall synthesis | cell wall, plasma membrane | 41 | 54.05 |
| 8 | Probable succinate dehydrogenase | Rv0248c | 646 | 71092 | anaerobic respiration | cell wall, plasma membrane | 39 | 34.9 |
| 9 | 35 kDa protein | 35kd_ag BCG_2760c | 270 | 29240 | NA | cell wall, cytoplasm, plasma membrane | 37.5 | 75.95 |
| 10 | ATP synthase subunit alpha (AtpA) | atpA BCG_1368 | 549 | 59480 | ATP production | cell wall, plasma membrane | 37.5 | 23.7 |
| 11 | Alpha-crystallin (HspX) | hspX BCG_2050c | 144 | 16217 | cellular response to stress, protein folding | cell wall, cytoplasm, plasma membrane | 37 | 67.7 |
| 12 | 30S ribosomal protein S1 (RpsA) | rpsA BCG_1668 | 481 | 53199 | translation | ribosome, cell wall, plasma membrane | 36 | 38.65 |
| 13 | ATP synthase subunit beta (AtpD) | atpD BCG_1370 | 486 | 53175 | ATP production | cell wall, cytoplasm, plasma membrane | 34 | 41.65 |
| 14 | Elongation factor Tu (Tuf) | tuf BCG_0734 | 396 | 43566 | response to stress, protein synthesis | cell wall, cytoplasm, plasma membrane | 33 | 40.75 |
| 15 | 60 kDa chaperonin 2 (GroEL2) | groL2 groEL2 BCG_0479 | 540 | 56692 | cellular response to stress, protein folding | cell wall, cytoplasm, plasma membrane | 31 | 34.55 |
| 16 | DNA topoisomerase 1 (TopA) | topA BCG_3704c | 934 | 102307 | DNA replication | cell wall, cytoplasm, plasma membrane | 27.5 | 30.45 |
| 17 | 29 kDa antigen (CFP29) | cfp29 BCG_0850c | 265 | 28870 | defense response | NA | 26 | 41.1 |
| 18 | 60 kDa chaperonin 1 (GroEL1) | groL1 groEL1 BCG_3487c | 539 | 55843 | cellular response to stress, protein folding | cell wall, cytoplasm, plasma membrane, nucleoid | 23.5 | 32.5 |
| 19 | Protein GrpE | grpE BCG_0390 | 235 | 24544 | cellular response to starvation protein folding | cell wall, cytoplasm | 22 | 28.95 |
| 20 | DNA-directed RNA polymerase subunit beta (RpoB) | rpoB BCG_0716 | 1178 | 130354 | transcription; response to antibiotic | cell wall, cytoplasm, plasma membrane | 21.5 | 34.95 |
| 21 | Chaperone protein DnaJ1 | dnaJ1 BCG_0391 | 395 | 41746 | cellular response to stress, protein folding | cell wall, cytoplasm, plasma membrane | 21 | 23.95 |
| 22 | Probable 3-oxoacyl-[acyl-carrier protein] reductase (FabG4) | fabG4 BCG_0280c | 454 | 46916 | NA | NA | 20.5 | 47.05 |
| 23 | Heparin-binding hemagglutinin (HbHA) | hbhA BCG_0516 | 199 | 21522 | pathogenesis | cell wall, cytoplasm, plasma membrane | 16 | 37.45 |
| 24 | Phosphate-binding protein PstS3 | pstS3 BCG_0980 | 370 | 38215 | ligand transport; pathogenesis | Plasma membrane, secreted | 10 | 21.75 |
| 25 | Lipoarabinomannan carrier protein (LprG) | lprG BCG_1472c | 236 | 24647 | ligand transport; pathogenesis | bacterial extracellular vesicle | 8.5 | 21.6 |
| 26 | Glyceraldehyde-3-phosphate dehydrogenase (Gap) | gap BCG_1497 | 339 | 36105 | metabolic process | cell wall, cytoplasm, plasma membrane | 5 | 14.8 |
| 27 | Lipoprotein LpqH | lpqH BCG_3822 | 159 | 15309 | ligand transport; pathogenesis | bacterial extracellular vesicle | 5 | 14.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohd Hanafiah, K.; Arifin, N.; Sanders, P.R.; Othman, N.; Garcia, M.L.; Anderson, D.A. Proteomic Analysis of Antigen 60 Complex of M. bovis Bacillus Calmette-Guérin Reveals Presence of Extracellular Vesicle Proteins and Predicted Functional Interactions. Vaccines 2019, 7, 80. https://doi.org/10.3390/vaccines7030080
Mohd Hanafiah K, Arifin N, Sanders PR, Othman N, Garcia ML, Anderson DA. Proteomic Analysis of Antigen 60 Complex of M. bovis Bacillus Calmette-Guérin Reveals Presence of Extracellular Vesicle Proteins and Predicted Functional Interactions. Vaccines. 2019; 7(3):80. https://doi.org/10.3390/vaccines7030080
Chicago/Turabian StyleMohd Hanafiah, Khayriyyah, Norsyahida Arifin, Paul R. Sanders, Nurulhasanah Othman, Mary L. Garcia, and David A. Anderson. 2019. "Proteomic Analysis of Antigen 60 Complex of M. bovis Bacillus Calmette-Guérin Reveals Presence of Extracellular Vesicle Proteins and Predicted Functional Interactions" Vaccines 7, no. 3: 80. https://doi.org/10.3390/vaccines7030080
APA StyleMohd Hanafiah, K., Arifin, N., Sanders, P. R., Othman, N., Garcia, M. L., & Anderson, D. A. (2019). Proteomic Analysis of Antigen 60 Complex of M. bovis Bacillus Calmette-Guérin Reveals Presence of Extracellular Vesicle Proteins and Predicted Functional Interactions. Vaccines, 7(3), 80. https://doi.org/10.3390/vaccines7030080