Food Additives as Novel Influenza Vaccine Adjuvants

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Influenza Vaccines and Compounds Used For The Screen
2.3. Immunization and Protection
2.4. Measurement of Virus-Specific Antibody Titers
2.5. Statistics
2.6. Ethics
3. Results
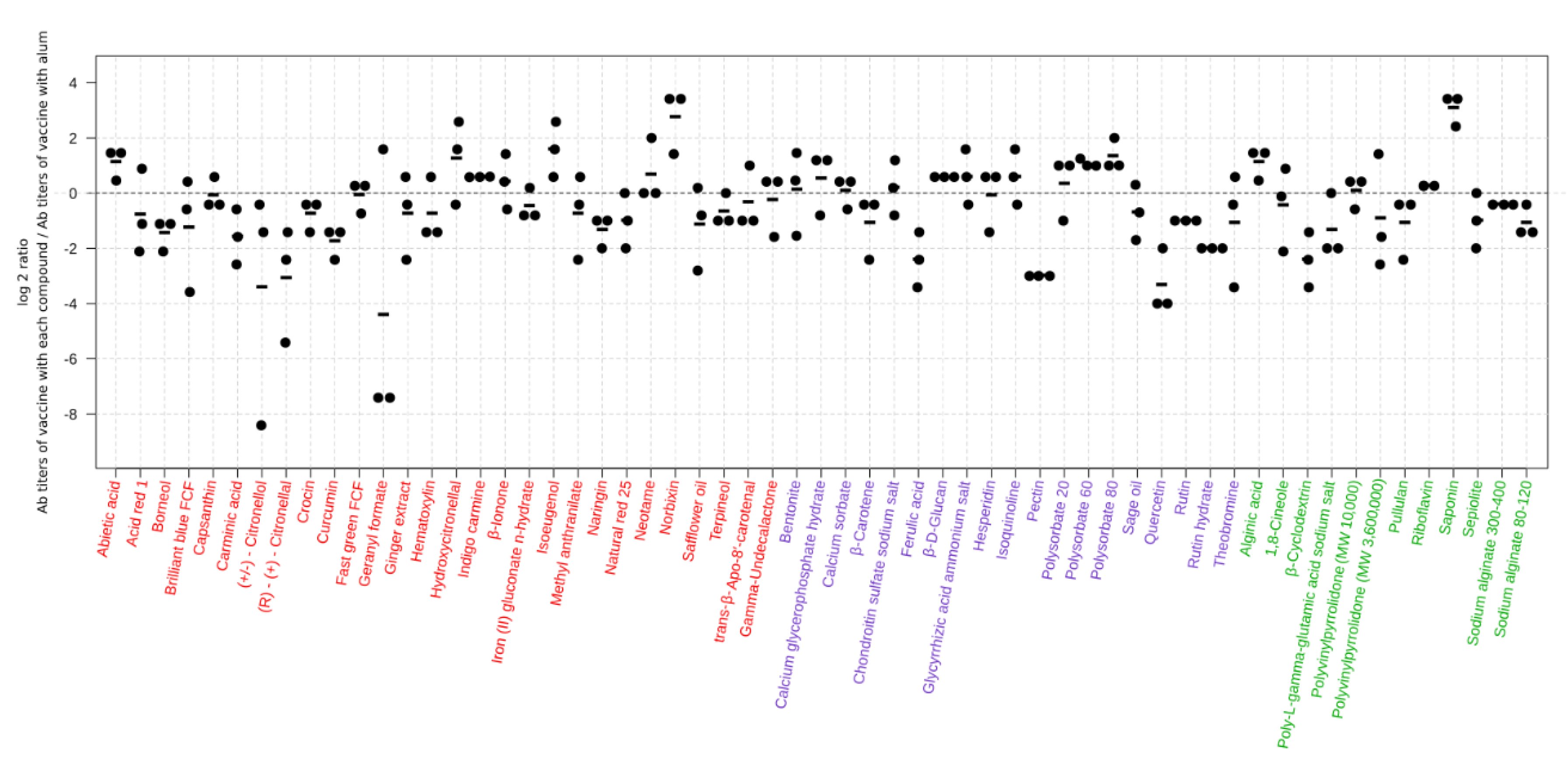
3.1. Identification of 59 Compounds that Enhance the Humoral Responses to an Influenza Vaccine in Mice
3.2. Identification of 41 Compounds that Enhance the Protective Efficacy of Influenza Vaccine against Lethal Virus Challenge in Mice
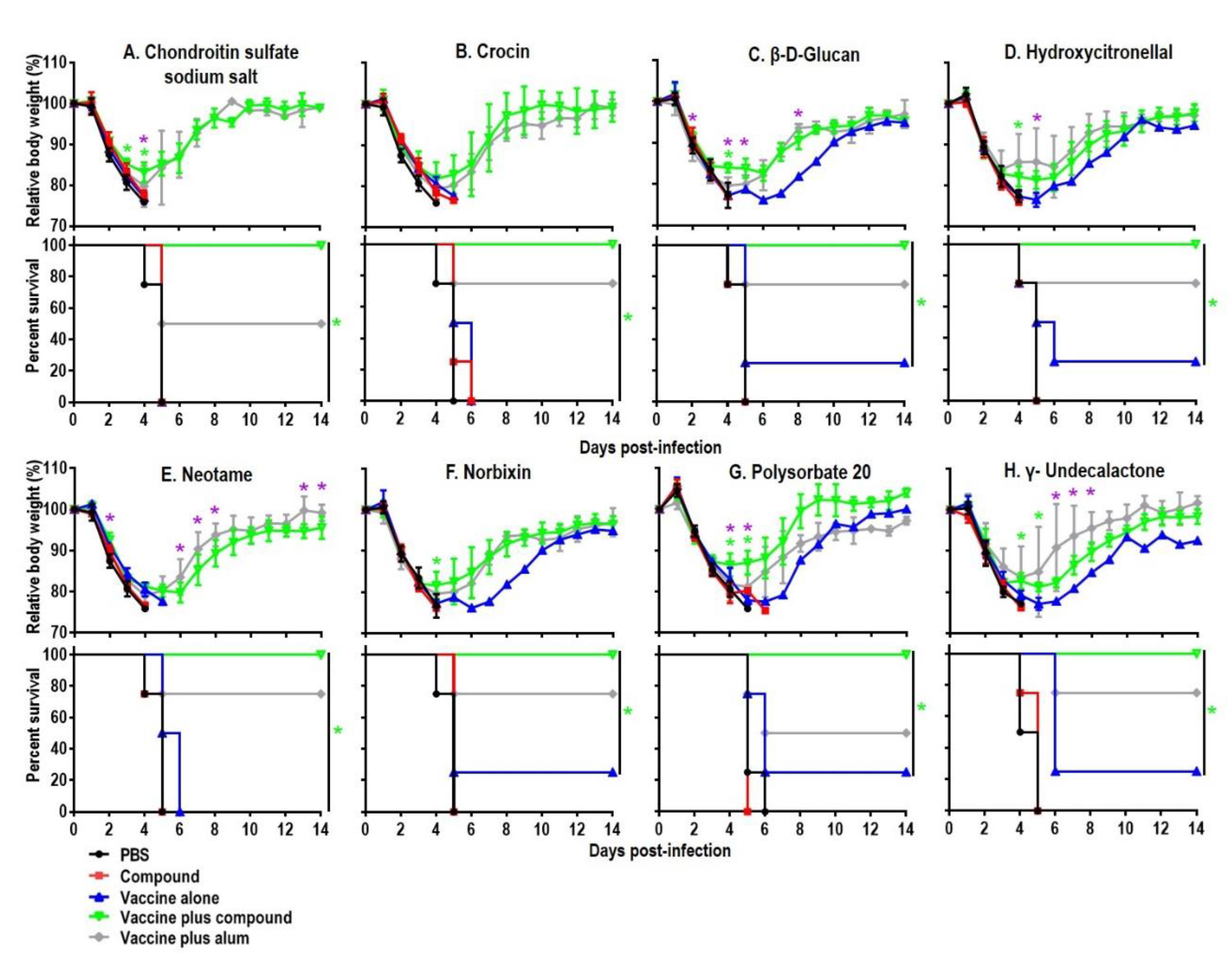
3.3. Effect of Immunizing Mice with the HA Vaccine Together with the Top Five Compounds on the Replication of the Challenge Virus in the Respiratory Tract
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Up to 650 000 People Die of Respiratory Diseases Linked to Seasonal Flu Each Year. Available online: http://www.who.int/mediacentre/news/statements/2017/flu/en/ (accessed on 1 July 2019).
- Li, C.; Bu, Z.; Chen, H. Avian influenza vaccines against H5N1 ‘bird flu’. Trends Biotechnol. 2014, 32, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Kreijtz, J.H.; Osterhaus, A.D.; Rimmelzwaan, G.F. Vaccination strategies and vaccine formulations for epidemic and pandemic influenza control. Hum. Vaccines 2009, 5, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Deng, G.; Ma, S.; Zeng, X.; Yin, X.; Li, M.; Zhang, B.; Cui, P.; Chen, Y.; Yang, H.; et al. Rapid Evolution of H7N9 Highly Pathogenic Viruses that Emerged in China in 2017. Cell Host Microbe 2018. [Google Scholar] [CrossRef] [PubMed]
- Hatta, M.; Gao, P.; Halfmann, P.; Kawaoka, Y. Molecular basis for high virulence of Hong Kong H5N1 influenza A viruses. Science 2001, 293, 1840–1842. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.S.; Webby, R.J. Traditional and new influenza vaccines. Clin. Microbiol. Rev. 2013, 26, 476–492. [Google Scholar] [CrossRef] [PubMed]
- WHO. Influenza (Seasonal). Available online: https://www.who.int/news-room/fact-sheets/detail/influenza-(seasonal) (accessed on 1 July 2019).
- Dominguez, A.; Godoy, P.; Torner, N. The Effectiveness of Influenza Vaccination in Different Groups. Expert Rev. Vaccines 2016, 15, 751–764. [Google Scholar] [CrossRef] [PubMed]
- Osterholm, M.T.; Kelley, N.S.; Sommer, A.; Belongia, E.A. Efficacy and effectiveness of influenza vaccines: A systematic review and meta-analysis. Lancet Infect. Dis. 2012, 12, 36–44. [Google Scholar] [CrossRef]
- Reed, S.G.; Orr, M.T.; Fox, C.B. Key roles of adjuvants in modern vaccines. Nat. Med. 2013, 19, 1597–1608. [Google Scholar] [CrossRef]
- Tregoning, J.S.; Russell, R.F.; Kinnear, E. Adjuvanted influenza vaccines. Hum. Vaccines Immunother. 2018, 14, 550–564. [Google Scholar] [CrossRef]
- Tetsutani, K.; Ishii, K.J. Adjuvants in influenza vaccines. Vaccine 2012, 30, 7658–7661. [Google Scholar] [CrossRef]
- Manzoli, L.; De Vito, C.; Salanti, G.; D’Addario, M.; Villari, P.; Ioannidis, J.P. Meta-analysis of the immunogenicity and tolerability of pandemic influenza A 2009 (H1N1) vaccines. PLoS ONE 2011, 6, e24384. [Google Scholar] [CrossRef] [PubMed]
- Szakacs, A.; Darin, N.; Hallbook, T. Increased childhood incidence of narcolepsy in western Sweden after H1N1 influenza vaccination. Neurology 2013, 80, 1315–1321. [Google Scholar] [CrossRef] [PubMed]
- Nohynek, H.; Jokinen, J.; Partinen, M.; Vaarala, O.; Kirjavainen, T.; Sundman, J.; Himanen, S.L.; Hublin, C.; Julkunen, I.; Olsen, P.; et al. AS03 adjuvanted AH1N1 vaccine associated with an abrupt increase in the incidence of childhood narcolepsy in Finland. PLoS ONE 2012, 7, e33536. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.X.; Xie, Y.; Ye, Y.P. Advances in saponin-based adjuvants. Vaccine 2009, 27, 1787–1796. [Google Scholar] [CrossRef] [PubMed]
- Ng, H.I.; Fernando, G.J.; Depelsenaire, A.C.; Kendall, M.A. Potent response of QS-21 as a vaccine adjuvant in the skin when delivered with the Nanopatch, resulted in adjuvant dose sparing. Sci. Rep. 2016, 6, 29368. [Google Scholar] [CrossRef]
- Morelli, A.B.; Becher, D.; Koernig, S.; Silva, A.; Drane, D.; Maraskovsky, E. ISCOMATRIX: A novel adjuvant for use in prophylactic and therapeutic vaccines against infectious diseases. J. Med. Microbiol. 2012, 61, 935–943. [Google Scholar] [CrossRef]
- Liu, H.; Patil, H.P.; de Vries-Idema, J.; Wilschut, J.; Huckriede, A. Enhancement of the immunogenicity and protective efficacy of a mucosal influenza subunit vaccine by the saponin adjuvant GPI-0100. PLoS ONE 2012, 7, e52135. [Google Scholar] [CrossRef]
- Cox, R.J.; Pedersen, G.; Madhun, A.S.; Svindland, S.; Saevik, M.; Breakwell, L.; Hoschler, K.; Willemsen, M.; Campitelli, L.; Nostbakken, J.K.; et al. Evaluation of a virosomal H5N1 vaccine formulated with Matrix M adjuvant in a phase I clinical trial. Vaccine 2011, 29, 8049–8059. [Google Scholar] [CrossRef]
- Sakabe, S.; Ozawa, M.; Takano, R.; Iwastuki-Horimoto, K.; Kawaoka, Y. Mutations in PA, NP, and HA of a pandemic (H1N1) 2009 influenza virus contribute to its adaptation to mice. Virus Res. 2011, 158, 124–129. [Google Scholar] [CrossRef] [Green Version]
- Gaush, C.R.; Smith, T.F. Replication and plaque assay of influenza virus in an established line of canine kidney cells. Appl. Microbiol. 1968, 16, 588–594. [Google Scholar]
- Uraki, R.; Das, S.C.; Hatta, M.; Kiso, M.; Iwatsuki-Horimoto, K.; Ozawa, M.; Coban, C.; Ishii, K.J.; Kawaoka, Y. Hemozoin as a novel adjuvant for inactivated whole virion influenza vaccine. Vaccine 2014, 32, 5295–5300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, H.P.; Yamashita, M.; Lopes, T.J.D.; Watanabe, T.; Kawaoka, Y. Injectable Excipients as Novel Influenza Vaccine Adjuvants. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Even-Or, O.; Samira, S.; Rochlin, E.; Balasingam, S.; Mann, A.J.; Lambkin-Williams, R.; Spira, J.; Goldwaser, I.; Ellis, R.; Barenholz, Y. Immunogenicity, protective efficacy and mechanism of novel CCS adjuvanted influenza vaccine. Vaccine 2010, 28, 6527–6541. [Google Scholar] [CrossRef] [PubMed]
- The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 1 July 2019).
- Bates, D.; Machler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- Lenth, R.V. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 33. [Google Scholar] [CrossRef]
- Han, S.K.; Lee, D.; Lee, H.; Kim, D.; Son, H.G.; Yang, J.S.; Lee, S.V.; Kim, S. OASIS 2: online application for survival analysis 2 with features for the analysis of maximal lifespan and healthspan in aging research. Oncotarget 2016, 7, 56147–56152. [Google Scholar] [CrossRef] [PubMed]
- Kusakabe, T.; Ozasa, K.; Kobari, S.; Momota, M.; Kishishita, N.; Kobiyama, K.; Kuroda, E.; Ishii, K.J. Intranasal hydroxypropyl-beta-cyclodextrin-adjuvanted influenza vaccine protects against sub-heterologous virus infection. Vaccine 2016, 34, 3191–3198. [Google Scholar] [CrossRef] [PubMed]
- Onishi, M.; Ozasa, K.; Kobiyama, K.; Ohata, K.; Kitano, M.; Taniguchi, K.; Homma, T.; Kobayashi, M.; Sato, A.; Katakai, Y.; et al. Hydroxypropyl-beta-cyclodextrin spikes local inflammation that induces Th2 cell and T follicular helper cell responses to the coadministered antigen. J. Immunol. 2015, 194, 2673–2682. [Google Scholar] [CrossRef]
- Yang, J.; Shim, S.M.; Nguyen, T.Q.; Kim, E.H.; Kim, K.; Lim, Y.T.; Sung, M.H.; Webby, R.; Poo, H. Poly-gamma-glutamic acid/chitosan nanogel greatly enhances the efficacy and heterosubtypic cross-reactivity of H1N1 pandemic influenza vaccine. Sci. Rep. 2017, 7, 44839. [Google Scholar] [CrossRef]
- Okamoto, S.; Matsuura, M.; Akagi, T.; Akashi, M.; Tanimoto, T.; Ishikawa, T.; Takahashi, M.; Yamanishi, K.; Mori, Y. Poly(gamma-glutamic acid) nano-particles combined with mucosal influenza virus hemagglutinin vaccine protects against influenza virus infection in mice. Vaccine 2009, 27, 5896–5905. [Google Scholar] [CrossRef]
- Bichurina, M.A.; Rozaeva, N.R.; Ishkil’din, M.I.; Degtiarev, A.A. Polyvinylpyrrolidone as a stimulator of the immunogenic activity of inactivated influenza vaccine. Tr. Inst. Im. Pastera 1982, 59, 109–112. [Google Scholar] [PubMed]
- Nagatomo, D.; Taniai, M.; Ariyasu, H.; Taniguchi, M.; Aga, M.; Ariyasu, T.; Ohta, T.; Fukuda, S. Cholesteryl Pullulan Encapsulated TNF-alpha Nanoparticles Are an Effective Mucosal Vaccine Adjuvant against Influenza Virus. Biomed. Res. Int. 2015, 2015, 471468. [Google Scholar] [CrossRef] [PubMed]
- Quintilio, W.; de Freitas, F.A.; Rodriguez, D.; Kubrusly, F.S.; Yourtov, D.; Miyaki, C.; de Cerqueira Leite, L.C.; Raw, I. Vitamins as influenza vaccine adjuvant components. Arch. Virol. 2016, 161, 2787–2795. [Google Scholar] [CrossRef] [PubMed]
- Marty-Roix, R.; Vladimer, G.I.; Pouliot, K.; Weng, D.; Buglione-Corbett, R.; West, K.; MacMicking, J.D.; Chee, J.D.; Wang, S.; Lu, S.; et al. Identification of QS-21 as an Inflammasome-activating Molecular Component of Saponin Adjuvants. J. Biol. Chem. 2016, 291, 1123–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.; Na, Y.J.; Yang, B.G.; Choi, J.P.; Seo, Y.B.; Hong, C.P.; Yun, C.H.; Kim, D.H.; Sohn, E.J.; Kim, J.H.; et al. Oral immunization of haemaggulutinin H5 expressed in plant endoplasmic reticulum with adjuvant saponin protects mice against highly pathogenic avian influenza A virus infection. Plant. Biotechnol. J. 2015, 13, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Bungener, L.; ter Veer, W.; Coller, B.A.; Wilschut, J.; Huckriede, A. Preclinical evaluation of the saponin derivative GPI-0100 as an immunostimulating and dose-sparing adjuvant for pandemic influenza vaccines. Vaccine 2011, 29, 2037–2043. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Hitzky, E.; Darder, M.; Aranda, P.; del Burgo, M.A.M.; del Real, G. Bionanocomposites as New Carriers for Influenza Vaccines. Adv. Mater. 2009, 21, 4167–4171. [Google Scholar] [CrossRef]
- Shechmeister, I.L.; Aeschliman, T.; Kammlade, W.G., Jr. Use of sodium alginate adjuvant in immunization against equine influenza. Am. J. Vet. Res. 1967, 28, 1373–1378. [Google Scholar]
- Morel, S.; Didierlaurent, A.; Bourguignon, P.; Delhaye, S.; Baras, B.; Jacob, V.; Planty, C.; Elouahabi, A.; Harvengt, P.; Carlsen, H.; et al. Adjuvant System AS03 containing alpha-tocopherol modulates innate immune response and leads to improved adaptive immunity. Vaccine 2011, 29, 2461–2473. [Google Scholar] [CrossRef]
- Didierlaurent, A.M.; Morel, S.; Lockman, L.; Giannini, S.L.; Bisteau, M.; Carlsen, H.; Kielland, A.; Vosters, O.; Vanderheyde, N.; Schiavetti, F.; et al. AS04, an aluminum salt- and TLR4 agonist-based adjuvant system, induces a transient localized innate immune response leading to enhanced adaptive immunity. J. Immunol. 2009, 183, 6186–6197. [Google Scholar] [CrossRef]
- Garcon, N.; Chomez, P.; Van Mechelen, M. GlaxoSmithKline Adjuvant Systems in vaccines: concepts, achievements and perspectives. Expert Rev. Vaccines 2007, 6, 723–739. [Google Scholar] [CrossRef]
- Roth, J.A. Veterinary vaccines and their importance to animal health and public health. Procedia Vaccinol. 2011, 5, 127–136. [Google Scholar] [CrossRef]
- Spickler, A.R.; Roth, J.A. Adjuvants in veterinary vaccines: modes of action and adverse effects. J. Vet. Intern. Med. 2003, 17, 273–281. [Google Scholar] [CrossRef]
- Heegaard, P.M.H.; Dedieu, L.; Johnson, N.; Le Potier, M.F.; Mockey, M.; Mutinelli, F.; Vahlenkamp, T.; Vascellari, M.; Sorensen, N.S. Adjuvants and delivery systems in veterinary vaccinology: current state and future developments. Arch. Virol. 2011, 156, 183–202. [Google Scholar] [CrossRef]
- Yamayoshi, S.; Kawaoka, Y. Current and future influenza vaccines. Nat. Med. 2019. [Google Scholar] [CrossRef]
- Pica, N.; Palese, P. Toward a universal influenza virus vaccine: prospects and challenges. Annu. Rev. Med. 2013, 64, 189–202. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Compound | Mean of Ab Titer-Primary Screen b | Ab Titer Ratio-Primary Sceen d (Compounds/Alum) | Mean of Ab Titer-Secondary Screen b | Ab Titer Ratio-Secondary Sceen d (Compounds/alum) | Function in Food | Status e | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Compound | Vaccine Alone | Vaccine and Compound | Vaccine and Alumc | Compound | Vaccine Alone | Vaccine and Compound | Vaccine and Alum c | |||||
| Abietic acid | <10 | <10 | 533.33 | 233.33 | 2.29 | <10 | 80, 160 f | 1280.00 | 560.00 | 2.29 | Preservative, acidity regulator | a |
| Acid red 1 | <10 | 10.00 f | 1173.33 | 1386.67 | 0.85 | <10 | 205.00 | 1760.00 | 2240.00 | 0.79 | Color | a |
| Alginic acid | <10 | <10 | 533.33 | 233.33 | 2.29 | <10 | 205.00 | 1280.00 | 2240.00 | 0.57 | Thickener, stabilizer | c |
| Bentonite | <10 | <10 | 346.67 | 233.33 | 1.49 | <10 | <10 | 140.00 | 600.00 | 0.23 | Anticaking agent | b |
| Borneol | <10 | 10 f | 533.33 | 1386.67 | 0.38 | <10 | 380.00 | 1920.00 | 2560.00 | 0.75 | Flavor | a |
| Brilliant blue FCF | <10 | <10 | 1333.33 | 1920.00 | 0.69 | <10 | <10 | 990.00 | 600.00 | 1.65 | Color | a |
| Calcium glycerophosphate hydrate | <10 | <10 | 1920.00 | 1120.00 | 1.71 | <10 | <10 | 4800.00 | 1440.00 | 3.33 | Thickener, gelling agent, stabilizer | b |
| Calcium sorbate | <10 | <10 | 2133.33 | 1920.00 | 1.11 | <10 | <10 | 3520.00 | 1440.00 | 2.44 | Preservative | b |
| Capsanthin | <10 | <10 | 1706.67 | 1706.67 | 1.00 | <10 | 380.00 | 800.00 | 840.00 | 0.95 | Color | a |
| Carminic acid | <10 | <10 | 746.67 | 1920.00 | 0.39 | <10 | 80, 40, 320 f | 960.00 | 1440.00 | 0.67 | Color | a |
| β-Carotene | <10 | <10 | 960.00 | 1706.67 | 0.56 | <10 | 80, 160 f | 1120.00 | 560.00 | 2.00 | Color | b |
| Chondroitin sulfate sodium salt | <10 | <10 | 1493.33 | 1120.00 | 1.33 | <10 | <10 | 2880.00 | 1440.00 | 2.00 | Emulsifier, stabilizer | b |
| 1,8-Cineole | <10 | 10 f | 1386.67 | 1386.67 | 1.00 | <10 | <10 | 990.00 | 1440.00 | 0.69 | Flavor | c |
| (+/−)-Citronellol | <10 | <10 | 1280, 640 f | 1706.67 | N/A | <10 | <10 | 260.00 | 720.00 | 0.36 | Flavor | a |
| (R)-(+)-Citronellal | <10 | <10 | 333.33 | 1706.67 | 0.20 | <10 | <10 | 40, 80, 20 f | 1600.00 | NA | Flavor | a |
| Crocin | <10 | <10 | 1066.67 | 1706.67 | 0.63 | <10 | <10 | 480.00 | 720.00 | 0.67 | Color | a |
| Curcumin | <10 | <10 | 1066.67 | 3413.33 | 0.31 | <10 | 380.00 | 480.00 | 840.00 | 0.57 | Color | a |
| β-Cyclodextrin | <10 | <10 | 373.33 | 1706.67 | 0.22 | <10 | <10 | 1440.00 | 1600.00 | 0.90 | Emulsifier | c |
| Fast green FCF | <10 | <10 | 2133.33 | 2133.33 | 1.00 | <10 | <10 | 440.00 | 600.00 | 0.73 | Color | a |
| Ferulic acid | <10 | <10 | 746.67 | 3413.33 | 0.22 | <10 | 380.00 | 640.00 | 840.00 | 0.76 | Antioxidant | b |
| Geranyl formate | <10 | <10 | 2560 f | 853.33 | N/A | <10 | <10 | 1280, 1280, 1280 f | 1600.00 | NA | Flavor | a |
| Ginger extract | <10 | <10 | 2773.33 | 3413.33 | 0.81 | <10 | 80, 40, 320 f | 1440.00 | 1440.00 | 1.00 | Antioxidant | a |
| β-d-Glucan | <10 | <10 | 2560.00 | 1706.67 | 1.50 | <10 | 80, 160 f | 2560.00 | 560.00 | 4.57 | Antioxidant | b |
| Glycyrrhizic acid ammonium salt | <10 | <10 | 5973.33 | 3413.33 | 1.75 | <10 | 80, 40, 320 f | 2560.00 | 1440.00 | 1.78 | Sweetener | b |
| Hematoxylin | <10 | <10 | 2560.00 | 3413.33 | 0.75 | <10 | 380.00 | 1200.00 | 840.00 | 1.43 | Color | a |
| Hesperidin | <10 | <10 | 3840.00 | 3413.33 | 1.13 | <10 | 80, 160 f | 2240.00 | 560.00 | 4.00 | Antioxidant, nutrient supplement | b |
| Hydroxycitronellal | <10 | <10 | 2773.33 | 853.33 | 3.25 | <10 | 80, 40, 320 f | 1280.00 | 1440.00 | 0.89 | Flavor | a |
| Indigo carmine | <10 | <10 | 1280.00 | 853.33 | 1.50 | <10 | <10 | 485.00 | 600.00 | 0.81 | Color | a |
| β-Ionone | <10 | 113.33 | 1493.33 | 960.00 | 1.56 | <10 | 380.00 | 220.00 | 2560.00 | 0.09 | Flavor | a |
| Iron (II) gluconate n-hydrate | <10 | <10 | 853.33 | 1120.00 | 0.76 | <10 | <10 | 3040.00 | 1600.00 | 1.90 | Color retention agent | a |
| Isoeugenol | <10 | <10 | 2986.67 | 853.33 | 3.50 | <10 | <10 | 140.00 | 720.00 | 0.19 | Flavor | a |
| Isoquinoline | <10 | <10 | 1493.33 | 853.33 | 1.75 | <10 | 80, 40, 320 f | 1200.00 | 1440.00 | 0.83 | Color | b |
| Methyl anthranilate | <10 | <10 | 693.33 | 853.33 | 0.81 | <10 | 380.00 | 2240.00 | 2560.00 | 0.88 | Flavor enhancer | a |
| Naringin | <10 | 320.00 | 2133.33 | 5120.00 | 0.42 | <10 | 380.00 | 2240.00 | 840.00 | 2.67 | Antioxidant | a |
| Natural red 25 | <10 | 320.00 | 2986.67 | 5120.00 | 0.58 | <10 | 300.00 | 440.00 | 2560.00 | 0.17 | Color | a |
| Neotame | <10 | 40, 80 f | 2560.00 | 1280.00 | 2.00 | <10 | <10 | 720.00 | 720.00 | 1.00 | Sweetener | a |
| Norbixin | <10 | 113.33 | 7680.00 | 960.00 | 8.00 | <10 | 80, 40, 320 f | 2880.00 | 560.00 | 5.14 | Color | a |
| Pectin | <10 | 320.00 | 640.00 | 5120.00 | 0.13 | <10 | <10 | 320.00 | 600.00 | 0.53 | Vegetable gum, emulsifier | b |
| Poly-L-γ-glutamic acid sodium salt | <10 | 320.00 | 2560.00 | 5120.00 | 0.50 | <10 | <10 | 40, 40, 160 f | 1600.00 | 0.04 | Flavor enhancer | c |
| Polysorbate 20 | <10 | 40, 80 f | 1920.00 | 1280.00 | 1.50 | <10 | 205.00 | 5760.00 | 2240.00 | 2.57 | Emulsifier | b |
| Polysorbate 60 | <10 | 40, 80 f | 2560.00 | 1280.00 | 2.00 | <10 | 205.00 | 2560.00 | 2240.00 | 1.14 | Emulsifier | b |
| Polyvinylpyrrolidone (MW 10,000) | <10 | <10 | 533.33 | 480.00 | 1.11 | <10 | <10 | 652.50 | 1600.00 | 0.41 | Emulsifier, stabilizer | c |
| Polysorbate 80 | <10 | 40, 80 f | 3416.33 | 1280.00 | 2.67 | <10 | <10 | 4160.00 | 1600.00 | 2.60 | Emulsifier | b |
| Polyvinylpyrrolidone (MW 3,600,000) | <10 | <10 | 506.67 | 480.00 | 1.06 | <10 | <10 | 2720.00 | 1600.00 | 1.70 | Emulsifier, stabilizer | c |
| Pullulan | <10 | <10 | 960.00 | 1706.67 | 0.56 | <10 | 380.00 | 720.00 | 840.00 | 0.86 | Thickener, glazing agent | c |
| Quercetin | <10 | 320.00 | 640.00 | 5120.00 | 0.13 | <10 | <10 | 300.00 | 1600.00 | 0.19 | Color | b |
| Riboflavin | <10 | <10 | 2560.00 | 2133.33 | 1.20 | <10 | <10 | 2240.00 | 1440.00 | 1.56 | Color | c |
| Rutin | <10 | 320.00 | 2560.00 | 5120.00 | 0.50 | <10 | <10 | 1320.00 | 1440.00 | 0.92 | Color, antioxidant, nutrient supplement | b |
| Rutin hydrate | <10 | 320.00 | 1280.00 | 5120.00 | 0.25 | <10 | <10 | 720.00 | 720.00 | 1.00 | Color, antioxidant, nutrient supplement | b |
| Safflower oil | <10 | <10 | 693.33 | 1120.00 | 0.62 | <10 | 80, 40, 320 f | 800.00 | 1440.00 | 0.56 | Flavor | a |
| Sage oil g | <10 | 320, 80 f | 746.67 | 960.00 | 0.78 | <10 | <10 | 160, 10 f | 1600.00 | N/A | Flavor enhancer | b |
| Saponin | <10 | <10 | 4266.67 | 480.00 | 8.89 | <10 | 205.00 | 12160.00 | 2240.00 | 5.43 | Foaming agent | c |
| Sepiolite | <10 | 320.00 | 2986.67 | 5120.00 | 0.58 | <10 | 300.00 | 2080.00 | 2560.00 | 0.81 | Acidity regulator, anticaking agent | c |
| Sodium alginate 300–400 | <10 | <10 | 1280.00 | 1706.67 | 0.75 | <10 | 205.00 | 480.00 | 2240.00 | 0.21 | Thickener, stabilizer | c |
| Sodium alginate 80–120 | <10 | <10 | 853.33 | 1706.67 | 0.50 | <10 | 205.00 | 1280.00 | 2240.00 | 0.57 | Thickener, stabilizer | c |
| Terpineol | <10 | <10 | 853.33 | 1280.00 | 0.67 | <10 | 380.00 | 2880.00 | 2560.00 | 1.13 | Flavor | a |
| Theobromine | <10 | <10 | 1333.33 | 1706.67 | 0.78 | <10 | 80, 160 f | 1440.00 | 560.00 | 2.57 | Flavor | b |
| trans-β-Apo-8′-carotenal | <10 | <10 | 1280.00 | 1280.00 | 1.00 | <10 | 380.00 | 1400.00 | 2560.00 | 0.55 | Color | a |
| γ-Undecalactone | <10 | 113.33 | 960.00 | 960.00 | 1.00 | <10 | 380.00 | 1920.00 | 840.00 | 2.29 | Flavor | a |
| Compound | Maximum Body Weight Loss % ± SD | Protective Efficacy (Survival/Total) | Enhanced Protective Efficacy (%) Compared to b | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Compound Alone | Vaccine Alone | Vaccine and Compound | Vaccine and Alum | Compound Alone | Vaccine Alone | Vaccine and Compound | Vaccine and Alum | Vaccine Alone | Vaccine and Alum | |
| Abietic acid e,* | 23.6 ± 0.6 | 23.9 | 19.3 ± 2.8 | 20.4 ± 2.0 | 0/4 | 1/4 | 3/4 | 3/4 | 50 | 0 |
| Brilliant blue FCF * | 23.1 ± 2.0 | 24.3 | 18.2 ± 4.9 | 16.3 ± 3.7 | 0/4 | 0/4 | 3/4 | 3/4 | 75 | 0 |
| Calcium glycerophosphate hydrate | 23.6 ± 0.9 | 23.0 ± 1.5 | 23.1 ± 2.4 | 20.2 ± 5.1 | 0/4 | 0/4 | 3/4 | 2/4 | 75 | 25 |
| Calcium sorbate | 23.4 ± 0.4 | 23.0 ± 1.5 | 19.5 ± 3.1 | 20.2 ± 5.1 | 0/4 | 0/4 | 4/4 | 2/4 | 100 | 50 |
| Carminic acid * | 22.8 ± 1.9 | 22.7 ± 1.7 | 22.8 ± 1.4 | 15.4 ± 7.6 | 0/4 | 1/4 | 4/4 | 3/4 | 75 | 25 |
| β-Carotene | 24.7 | 23.9 | 17.4 ± 6.0 | 20.4 ± 3.7 | 0/4 | 1/4 | 4/4 | 3/4 | 75 | 25 |
| Chondroitin sulfate sodium salt | 22.6 ± 1.3 | 23.0 ± 1.5 | 16.8 ± 2.4 | 20.2 ± 5.1 | 0/4 | 0/4 | 4/4 | 2/4 | 100 | 50 |
| (+/−)-Citronellol * | 22.0 ± 1.8 | 22.4 ± 0.7 | 19.4 ± 2.1 | 20.9 ± 2.4 | 0/4 | 0/4 | 3/4 | 3/4 | 75 | 0 |
| (R)-(+)-Citronellal * | 24.1 ± 0.5 | 24.1 | 19.7 ± 4.3 | 17.6 ± 3.1 | 0/4 | 0/4 | 2/4 | 2/4 | 50 | 0 |
| Crocin * | 19.4 ± 0.7 | 21.6 | 18.2 ± 4.0 | 20.9 ± 2.4 | 0/4 | 0/4 | 4/4 | 3/4 | 100 | 25 |
| β-Cyclodextrin | 23.9 | 24.1 | 20.7 ± 2.8 | 17.6 ± 3.1 | 0/4 | 0/4 | 4/4 | 2/4 | 100 | 50 |
| Fast green FCF * | 22.1 ± 3.6 | 24.3 | 18.1 ± 3.2 | 16.3 ± 3.7 | 0/4 | 0/4 | 2/4 | 3/4 | 50 | –25 |
| Geranyl formate * | 23.0 | 24.3 | 20.2 ± 5.0 | 16.3 ± 3.7 | 0/4 | 0/4 | 2/4 | 3/4 | 50 | –25 |
| β-d-Glucan | 23.1 ± 0.6 | 23.9 | 17.4 ± 1.8 | 20.4 ± 2.0 | 0/4 | 1/4 | 4/4 | 3/4 | 75 | 25 |
| Glycyrrhizic acid ammonium salt | 24.7 | 23.5 ± 1.7 | 20.3 ± 3.4 | 15.4 ± 7.6 | 0/4 | 1/4 | 4/4 | 3/4 | 75 | 25 |
| Hesperidin | 21.8 ± 0.6 | 23.9 | 20.2 ± 2.2 | 20.4 ± 2.0 | 0/4 | 1/4 | 4/4 | 3/4 | 75 | 25 |
| Hydroxycitronellal * | 24.2 ± 0.7 | 23.5 ± 1.7 | 18.7 ± 2.2 | 16.1 ± 4.7 | 0/4 | 1/4 | 4/4 | 3/4 | 75 | 25 |
| Indigo carmine * | 24.2 ± 1.0 | 24.3 | 18.5 ± 3.6 | 16.3 ± 3.7 | 0/4 | 0/4 | 2/4 | 3/4 | 50 | −25 |
| Iron (II) gluconate n-hydrate * | 23.6 ± 0.8 | 24.3 | 18.7 ± 2.1 | 18.6 ± 4.2 | 0/4 | 0/4 | 3/4 | 2/4 | 75 | 25 |
| Isoeugenol * | 22.3 ± 0.9 | 22.4 ± 0.7 | 21.7 ± 0.1 | 20.9 ± 2.4 | 0/4 | 0/4 | 2/4 | 3/4 | 50 | −25 |
| Isoquinoline | 24.1 | 23.5 ± 1.7 | 21.2 ± 2.9 | 16.1 ± 4.7 | 0/4 | 1/4 | 3/4 | 3/4 | 50 | 0 |
| Methyl anthranilate * | 21.5 ± 3.2 | 22.1 ± 1.6 | 19.3 ± 1.0 | 20.0 ± 1.2 | 0/4 | 1/4 | 4/4 | 3/4 | 75 | 25 |
| Naringin * | 23.6 ± 0.8 | 22.9 ± 1.5 | 20.9 ± 2.9 | 16.4 ± 7.5 | 0/4 | 1/4 | 3/4 | 3/4 | 50 | 0 |
| Neotame * | 23.4 ± 0.8 | 22.4 ± 0.7 | 20.2 ± 2.4 | 20.9 ± 2.4 | 0/4 | 0/4 | 4/4 | 3/4 | 100 | 25 |
| Norbixin * | 23.9 ± 0.8 | 23.9 | 18.3 ± 3.3 | 20.4 ± 2.0 | 0/4 | 1/4 | 4/4 | 3/4 | 75 | 25 |
| Pectin | 24.5 | 24.3 | 21.8 ± 1.6 | 16.3 ± 3.7 | 0/4 | 0/4 | 2/4 | 3/4 | 50 | −25 |
| Poly-L-γ-glutamic acid sodium salt | 23.7 | 24.1 | 18.9 ± 3.7 | 17.6 ± 3.1 | 0/4 | 0/4 | 2/4 | 2/4 | 50 | 0 |
| Polysorbate 20 | 24.6 | 22.5 | 13.5 ± 2.8 | 18.8 ± 5.5 | 0/4 | 1/4 | 4/4 | 2/4 | 75 | 50 |
| Polysorbate 60 c | 21.9 ± 0.6 | 22.5 | 19.5 ± 4.7 | 18.8 ± 5.5 | 0/4 | 1/4 | 2/4 | 2/4 | 25 | 0 |
| Polysorbate 80 | 23.2 | 24.3 | 22.8 ± 1.5 | 20.0 ± 4.2 | 0/4 | 0/4 | 2/3d | 2/4 | 66.7 | 16.7 |
| Polyvinylpyrrolidone (MW 3,600,000) | 22.6 ± 1.3 | 24.1 | 19.9 ± 3.0 | 17.6 ± 3.1 | 0/4 | 0/4 | 3/4 | 2/4 | 75 | 25 |
| Pullulan | 22.2 ± 0.8 | 22.9 ± 1.5 | 21.4 ± 1.7 | 16.4 ± 7.5 | 0/4 | 1/4 | 3/4 | 3/4 | 50 | 0 |
| Riboflavin | 23.2 ± 1.1 | 23.0 ± 1.5 | 22.0 ± 2.2 | 20.2 ± 5.1 | 0/4 | 0/4 | 4/4 | 2/4 | 100 | 50 |
| Rutin | 22.8 ± 1.4 | 23.0 ± 1.5 | 20.7 ± 2.5 | 20.2 ± 5.1 | 0/4 | 0/4 | 2/4 | 2/4 | 50 | 0 |
| Rutin hydrate | 21.7 ± 1.1 | 19.4 ± 0.7 | 18.0 ± 2.9 | 20.9 ± 2.4 | 0/4 | 0/4 | 4/4 | 3/4 | 100 | 25 |
| Saponin | 23.3 | 22.5 | 16.6 ± 3.9 | 18.8 ± 5.5 | 0/4 | 1/4 | 4/4 | 2/4 | 75 | 50 |
| Sepiolite | 22.2 ± 1.3 | 22.1 ± 1.6 | 19.0 ± 1.9 | 20.0 ± 1.2 | 0/4 | 1/4 | 3/4 | 3/4 | 50 | 0 |
| Sodium alginate 80–120 | 24.2 ± 0.4 | 22.5 | 16.9 ± 2.3 | 18.8 ± 5.5 | 0/4 | 1/4 | 4/4 | 2/4 | 75 | 50 |
| Terpineol * | 22.7 ± 0.7 | 22.1 ± 1.6 | 19.2 ± 2.6 | 20.0 ± 1.2 | 0/4 | 1/4 | 4/4 | 3/4 | 75 | 25 |
| Theobromine | 23.1 ± 0.5 | 23.9 | 18.4 ± 2.4 | 20.4 ± 2.0 | 0/4 | 1/4 | 4/4 | 3/4 | 75 | 25 |
| γ- Undecalactone * | 23.8 ± 0.8 | 22.9 ± 1.5 | 18.8 ± 1.1 | 16.4 ± 7.5 | 0/4 | 1/4 | 4/4 | 3/4 | 75 | 25 |
| Immunogen | Mean Virus Titers (Log10 PFU/g) ± SD | |||
|---|---|---|---|---|
| Nasal Turbinates | Lungs | |||
| Day 3 p. i. | Day 6 p. i. | Day 3 p. i. | Day 6 p. i. | |
| PBS | 6.6 ± 0.2 | 5.9 ± 0.3 | 7.7 ± 0.1 | 6.7 ± 0.4 |
| β-d-Glucan | 6.4 ± 0.1 | N/A b, 5.3, 5.3 | 7.6 ± 0.1 | N/A, 6.2, 5.5 |
| Neotame | 6.3 ± 0.1 | 6.2, 5.5, N/A | 7.5 ± 0.2 | 7, 6.5, N/A |
| Norbixin | 6.7 ± 0.2 | 5.6 ± 0.3 | 7.4 ± 0.3 | 6.4 ± 0.3 |
| Polysorbate 20 | 6.8 ± 0.02 | 5.8 ± 0.3 | 7.5 ± 0.2 | 6.5 ± 0.4 |
| γ-Undecalactone | 6.7 ± 0.1 | 6.0 ± 0.3 | 7.4 ± 0.2 | 6.2 ± 0.2 |
| Vaccine alone | 6.3 ± 0.2 | 5.4 ± 0.7 | 7.4 ± 0.2 | 5.7 ± 1.0 |
| Vaccine and β-d-glucan | 6.0 ± 0.3 | 4.7 ± 0.9 | 7.2 ± 0.3 | 5.2 ± 0.7 |
| Vaccine and neotame | 6.1 ± 0.05 | 3.3 ± 0.4 | 7.5 ± 0.3 | 5.9, 4.4 |
| Vaccine and norbixin | 6.0 ± 0.3 | 4.2 ± 1.1 | 7.2 ± 0.3 | 4.4 ± 1.0 |
| Vaccine and polysorbate 20 | 6.1 ± 0.1 | 5.4, 5.4 | 7.2 ± 0.2 | 5.7, 5.0 |
| Vaccine and γ-undecalactone | 6.2 ± 0.2 | 4.8 ± 0.3 | 7.0 ± 0.3 | 4.4 ± 1.2 |
| Vaccine and Alum | 6.3 ± 0.1 | 3. 6 ± 0.7 | 7.4 ± 0.3 | 2.8 c |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, H.; Yamashita, M.; Wu, L.; Jose da Silva Lopes, T.; Watanabe, T.; Kawaoka, Y. Food Additives as Novel Influenza Vaccine Adjuvants. Vaccines 2019, 7, 127. https://doi.org/10.3390/vaccines7040127
Feng H, Yamashita M, Wu L, Jose da Silva Lopes T, Watanabe T, Kawaoka Y. Food Additives as Novel Influenza Vaccine Adjuvants. Vaccines. 2019; 7(4):127. https://doi.org/10.3390/vaccines7040127
Chicago/Turabian StyleFeng, Huapeng, Makoto Yamashita, Li Wu, Tiago Jose da Silva Lopes, Tokiko Watanabe, and Yoshihiro Kawaoka. 2019. "Food Additives as Novel Influenza Vaccine Adjuvants" Vaccines 7, no. 4: 127. https://doi.org/10.3390/vaccines7040127
APA StyleFeng, H., Yamashita, M., Wu, L., Jose da Silva Lopes, T., Watanabe, T., & Kawaoka, Y. (2019). Food Additives as Novel Influenza Vaccine Adjuvants. Vaccines, 7(4), 127. https://doi.org/10.3390/vaccines7040127