Akt+ IKKα/β+ Rab5+ Signalosome Mediate the Endosomal Recruitment of Sec61 and Contribute to Cross-Presentation in Bone Marrow Precursor Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Reagents and Antibodies
2.3. Murine Bone Marrow Precursor Cell Culture
2.4. RNAi Transfection
2.5. Bone Marrow Precursor Cell Treatments
2.6. Flow Cytometric Measurements
2.7. Co-Immunoprecipitation
2.8. Western Blots
2.9. Statistical Analysis
3. Results
3.1. Endotoxin-Containing Pathogen and Endotoxin-Free Antigen Induce Different Effect on the Phosphorylation of Akt, IKKα/β in Bone Marrow Precursor Cells
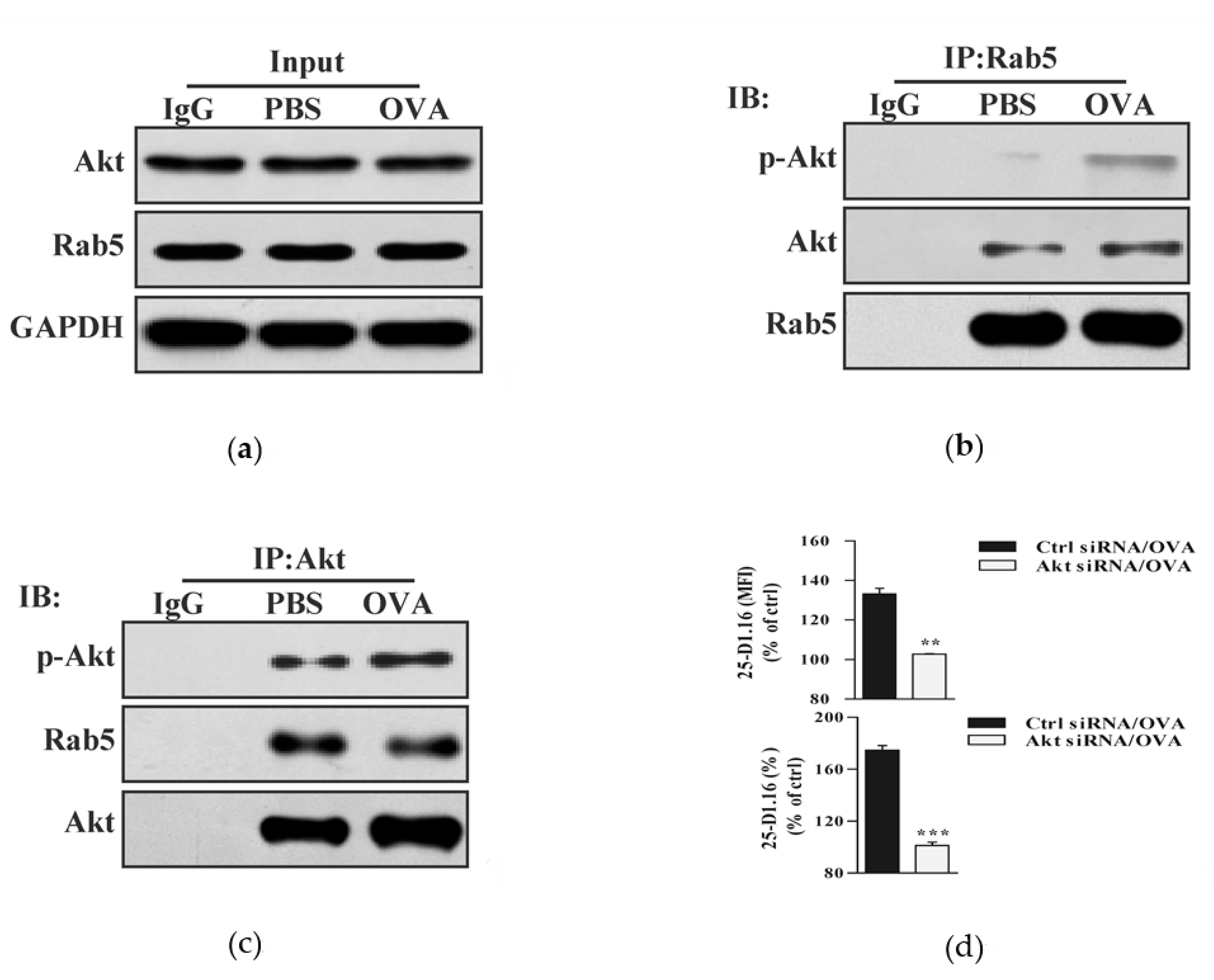
3.2. Akt Phosphorylation Is Essential for Endotoxin-Containing Pathogen-Derived Cross-Presentation in Bone Marrow Precursor Cells
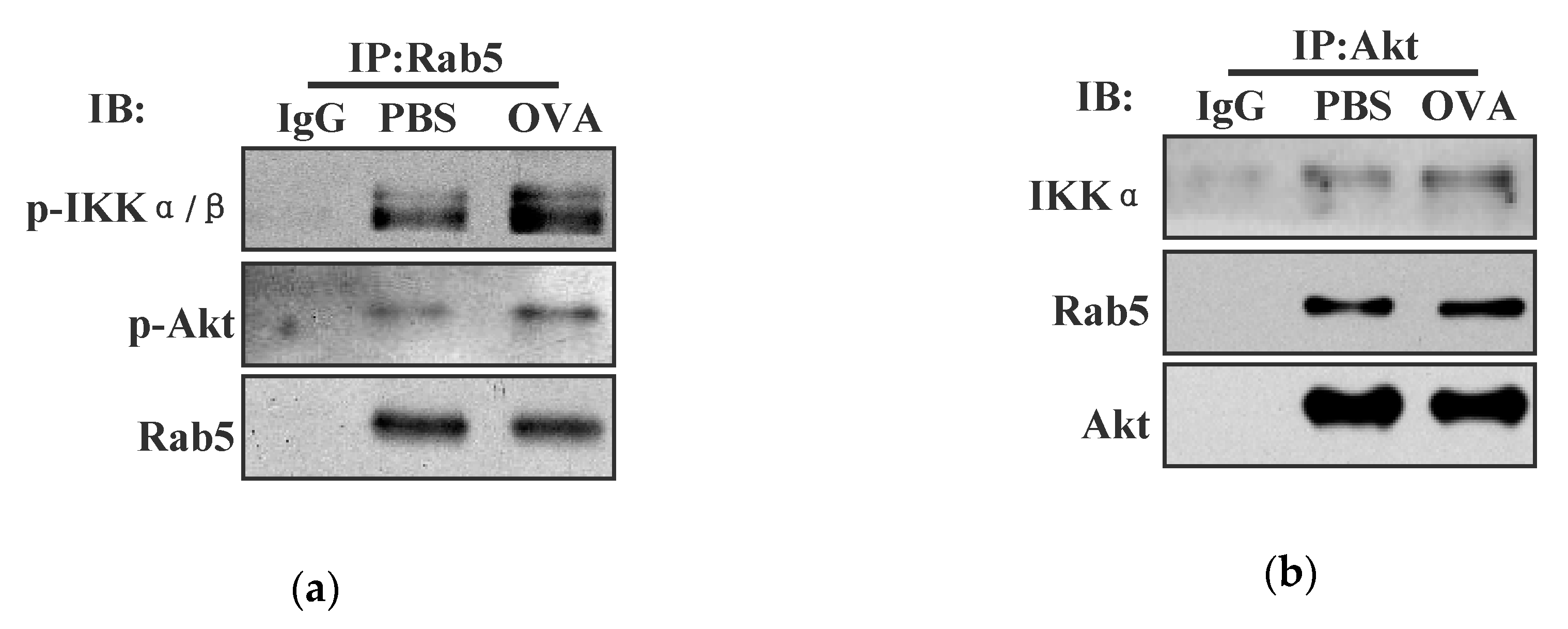
3.3. Both IKKα and IKKβ Phosphorylation Augment Endotoxin-Containing Pathogen-Derived Cross-Presentation in Bone Marrow Precursor Cells
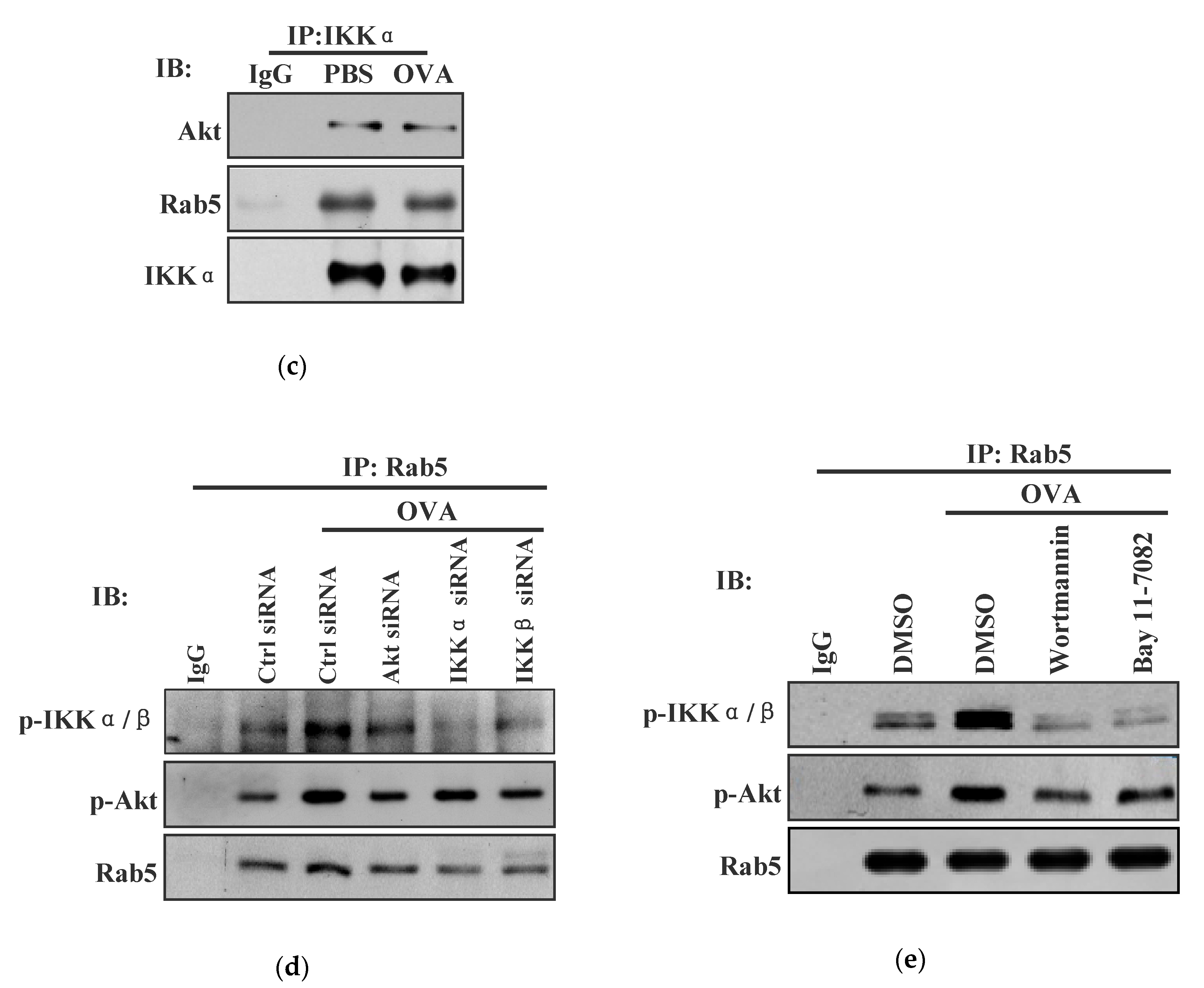
3.4. Endotoxin-Containing Pathogen Promotes the Formation of Akt+ Ikkα/Β+ Rab5+ Signalosome in Bone Marrow Precursor Cells
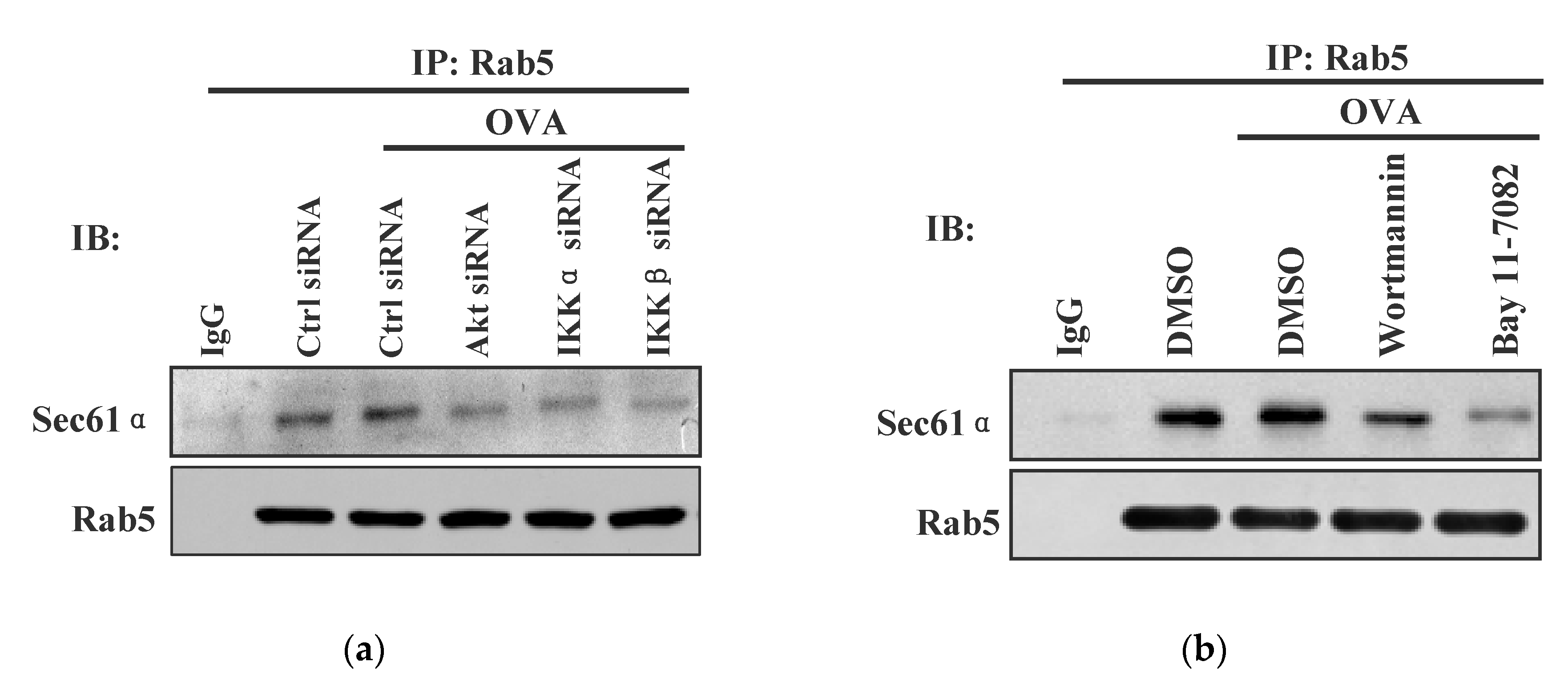
3.5. Akt+ IKKα/β+ Rab5+ Signalosome Facilitates Endotoxin-Containing Pathogen-Induced the Endosomal Relocation of Sec61α
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Seya, T.; Shime, H.; Takeda, Y.; Tatematsu, M.; Takashima, K.; Matsumoto, M. Adjuvant for vaccine immunotherapy of cancer—Focusing on Toll-like receptor 2 and 3 agonists for safely enhancing antitumor immunity. Cancer Sci. 2015, 106, 1659–1668. [Google Scholar] [CrossRef] [Green Version]
- Villadangos, J.A.; Heath, W.R.; Carbon, F.R. Outside looking in: The inner workings of the cross-presentation pathway within dendritic cells. Trends Immunol. 2007, 28, 45–47. [Google Scholar] [CrossRef] [PubMed]
- Gros, M.; Amigorena, S. Regulation of Antigen Export to the Cytosol During Cross-Presentation. Front. Immunol. 2019, 10, 41. [Google Scholar] [CrossRef] [PubMed]
- Zehner, M.; Chasan, A.I.; Schuette, V.; Embgenbroich, M.; Quast, T.; Kolanus, W.; Burgdorf, S. Mannose receptor polyubiquitination regulates endosomal recruitment of p97 and cytosolic antigen translocation for cross-presentation. Proc. Natl. Acad. Sci. USA 2011, 108, 9933–9938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zehner, M.; Burgdorf, S. Regulation of antigen transport into the cytosol for cross-presentation by ubiquitination of the mannose receptor. Mol. Immunol. 2013, 55, 146–148. [Google Scholar] [CrossRef] [PubMed]
- Burgdorf, S.; Schölz, C.; Kautz, A.; Tampé, R.; Kurts, C. Spatial and mechanistic separation of cross-presentation and endogenous antigen presentation. Nat. Immunol. 2008, 9, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Schuette, V.; Burgdorf, S. The ins-and-outs of endosomal antigens for cross-presentation. Curr. Opin. Immunol. 2014, 26, 63–68. [Google Scholar] [CrossRef]
- Baker, K.; Qiao, S.W.; Kuo, T.T.; Aveson, V.G.; Platzer, B.; Andersen, J.T.; Sandlie, I.; Chen, Z.; de Haar, C.; Lencer, W.I.; et al. Neonatal Fc receptor for IgG (FcRn) regulates cross-presentation of IgG immune complexes by CD8-CD11b+ dendritic cells. Proc. Natl. Acad. Sci. USA 2011, 108, 9927–9932. [Google Scholar] [CrossRef] [Green Version]
- Wen, H.; Fu, Z.; Wei, Y.; Zhang, X.; Ma, L.; Gu, L.; Li, J. Antioxidant Activity and Neuroprotective Activity of Stilbenoids in Rat Primary Cortex Neurons via the PI3K/Akt Signalling Pathway. Molecules 2018, 23, E2328. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, M.; McAuliffe, B.; Subramani, J.; Basu, S.; Shapiro, L.H. CD13 regulates dendritic cell cross-presentation and T cell responses by inhibiting receptor-mediated antigen uptake. J. Immunol. 2012, 188, 5489–5499. [Google Scholar] [CrossRef] [Green Version]
- Linley, A.; Krysov, S.; Ponzoni, M.; Johnson, P.W.; Packham, G.; Stevenson, F.K. Lectin binding to surface Ig variable regions provides a universal persistent activating signal for follicular lymphoma cells. Blood 2015, 126, 1902–1910. [Google Scholar] [CrossRef] [Green Version]
- Jane-wit, D.; Surovtseva, Y.V.; Qin, L.; Li, G.; Liu, R.; Clark, P.; Manes, T.D.; Wang, C.; Kashgarian, M.; Kirkiles-Smith, N.C.; et al. Complement membrane attack complexes activate noncanonical NF-κB by forming an Akt+ NIK+ signalosome on Rab5+ endosomes. Proc. Natl. Acad. Sci. USA 2015, 112, 9686–9691. [Google Scholar] [CrossRef] [Green Version]
- Fornaro, M.; Plescia, J.; Chheang, S.; Tallini, G.; Zhu, Y.M.; King, M.; Altieri, D.C.; Languino, L.R. Fibronectin protects prostate cancer cells from tumor necrosis factor-alpha-induced apoptosis via the AKT/survivin pathway. J. Biol. Chem. 2003, 278, 50402–50411. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Latif, M.M.; Kelleher, D.; Reynolds, J.V. Molecular mechanisms of constitutive and inducible NF-κB activation in activation in oesophageal adenocarcinoma. Eur. J. Cancer 2015, 51, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Hu, C.F.; Li, J.; You, X.; Gao, F.G. Increased translocation of antigens to endosomes and TLR4 mediated endosomal recruitment of TAP contribute to nicotine augmented cross-presentation. Oncotarget 2016, 7, 38451–38466. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Meyer, H.H.; Rapoport, T.A. The AAA ATPase Cdc48/p97 and its partners transport proteins from the ER into the cytosol. Nature 2001, 414, 652–656. [Google Scholar] [CrossRef]
- Ménager, J.; Ebstein, F.; Oger, R.; Hulin, P.; Nedellec, S.; Duverger, E.; Lehmann, A.; Kloetzel, P.M.; Jotereau, F.; Guilloux, Y. Cross-presentation of synthetic long peptides by human dendritic cells: A process dependent on ERAD component p97/VCP but Not sec61 and/or Derlin-1. PLoS ONE 2014, 9, e89897. [Google Scholar]
- Grotzke, J.E.; Kozik, P.; Morel, J.D.; Impens, F.; Pietrosemoli, N.; Cresswell, P.; Amigorena, S.; Demangel, C. Sec61 blockade by mycolactone inhibits antigen cross-presentation independently of endosome-to-cytosol export. Proc. Natl. Acad. Sci. USA 2017, 114, E5910–E5919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zehner, M.; Marschall, A.L.; Bos, E.; Schloetel, J.G.; Kreer, C.; Fehrenschild, D.; Limmer, A.; Ossendorp, F.; Lang, T.; Koster, A.J.; et al. The translocon protein Sec61 mediates antigen transport from endosomes in the cytosol for cross-presentation to CD8(+) T cells. Immunity 2015, 42, 850–863. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.G.; Wan, D.F.; Gu, J.R. Ex vivo nicotine stimulation augments the efficacy of therapeutic bone marrow-derived dendritic cell vaccination. Clin. Cancer Res. 2007, 13, 3706–37012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, H.X.; Ke, S.Z.; Xu, D.D.; Lu, N.N.; Wang, Y.N.; Zhang, Y.H.; Gao, F.G. Nicotine induces TIPE2 upregulation and Stat3 phosphorylation contributes to cholinergic anti-inflammatory effect. Int. J. Oncol. 2017, 51, 987–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everts, B.; Amiel, E.; Huang, S.C.; Smith, A.M.; Chang, C.H.; Lam, W.Y.; Redmann, V.; Freitas, T.C.; Blagih, J.; van der Windt, G.J.; et al. TLR-driven early glycolytic reprogramming via the kinases TBK1-IKKε supports the anabolic demands of dendritic cell activation. Nat. Immunol. 2014, 15, 323–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.A.; Kim, T.J.; Moon, H.J.; Kim, Y.J.; Yoon, H.K.; Seong, S.Y. Cardiolipin activates antigen-presenting cells via TLR2-PI3K-PKN1-AKT/p38-NF-kB signaling to prime antigen-specific naïve T cells in mice. Eur. J. Immunol. 2018, 48, 777–790. [Google Scholar] [CrossRef] [Green Version]
- Bazan, S.B.; Geginat, G.; Breinig, T.; Schmitt, M.J.; Breinig, F. Uptake of various yeast genera by antigen-presenting cells and influence of subcellular antigen localization on the activation of ovalbumin-specific CD8 T lymphocytes. Vaccine 2011, 29, 8165–8173. [Google Scholar] [CrossRef]
- Lee, Y.R.; Yang, I.H.; Lee, Y.H.; Im, S.A.; Song, S.; Li, H.; Han, K.; Kim, K.; Eo, S.K.; Lee, C.K. Cyclosporin A and tacrolimus, but not rapamycin, inhibit MHC-restricted antigen presentation pathways in dendritic cells. Blood 2005, 105, 3951–3955. [Google Scholar] [CrossRef] [Green Version]
- Nair-Gupta, P.; Baccarini, A.; Tung, N.; Seyffer, F.; Florey, O.; Huang, Y.; Banerjee, M.; Overholtzer, M.; Roche, P.A.; Tampé, R.; et al. TLR signals induce phagosomal MHC-I delivery from the endosomal recycling compartment to allow cross-presentation. Cell 2014, 158, 506–521. [Google Scholar] [CrossRef] [Green Version]
- Lu, B.; Ren, Y.; Sun, X.; Han, C.; Wang, H.; Chen, Y.; Peng, Q.; Cheng, Y.; Cheng, X.; Zhu, Q.; et al. Induction of INKIT by Viral Infection Negatively Regulates Antiviral Responses through Inhibiting Phosphorylation of p65 and IRF3. Cell Host Microbe 2017, 22, 86–98.e4. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Jin, R.; Gao, M.; Xu, H.; Zou, S.; Li, X.; Xing, C.; Wang, Q.; Wang, H.; Feng, J.; et al. Transcriptional repression of IKKβ by p53 in arsenite-induced GADD45α accumulation and apoptosis. Oncogene 2019, 38, 731–746. [Google Scholar] [CrossRef] [Green Version]
- Farrand, L.; Kim, J.Y.; Byun, S.; Im-aram, A.; Lee, J.; Suh, J.Y.; Lee, K.W.; Lee, H.J.; Tsang, B.K. The diarylheptanoid hirsutenone sensitizes chemoresistant ovarian cancer cells to cisplatin via modulation of apoptosis-inducing factor and X-linked inhibitor of apoptosis. J. Biol. Chem. 2014, 289, 1723–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, H.Q.; Park, S.J.; Shin, E.J.; Tran, T.V.; Sharma, N.; Lee, Y.J.; Jeong, J.H.; Jang, C.G.; Kim, D.J.; Nabeshima, T.; et al. Clozapine attenuates mitochondrial burdens and abnormal behaviors elicited by phencyclidine in mice via inhibition of p47 phox; Possible involvements of phosphoinositide 3-kinase/Akt signaling. J. Psychopharmacol. 2018, 32, 1233–1251. [Google Scholar] [CrossRef]
- Puc, J.; Keniry, M.; Li, H.S.; Pandita, T.K.; Choudhury, A.D.; Memeo, L.; Mansukhani, M.; Murty, V.V.; Gaciong, Z.; Meek, S.E.; et al. Lack of PTEN sequesters CHK1 and initiates genetic instability. Cancer Cell 2005, 7, 193–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacina, K.S.; Park, G.Y.; Bae, S.S.; Guzzetta, A.W.; Schaefer, E.; Birnbaum, M.J.; Roth, R.A. Identification of a proline-rich Akt substrate as a 14-3-3 binding partner. J. Biol. Chem. 2003, 278, 10189–10194. [Google Scholar] [CrossRef] [Green Version]
- Peruchetti, D.B.; Silva-Aguiar, R.P.; Siqueira, G.M.; Dias, W.B.; Caruso-Neves, C. High glucose reduces megalin-mediated albumin endocytosis in renal proximal tubule cells through protein kinase B O-GlcNAcylation. J. Biol. Chem. 2018, 293, 11388–11400. [Google Scholar] [CrossRef] [Green Version]
- Borg, M.L.; Massart, J.; Schönke, M.; De Castro Barbosa, T.; Guo, L.; Wade, M.; Alsina-Fernandez, J.; Miles, R.; Ryan, A.; Bauer, S.; et al. Modified UCN2 Peptide Acts as an Insulin Sensitizer in Skeletal Muscle of Obese Mice. Diabetes 2019, 68, 1403–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasuriya, N.; Kunkel, M.T.; Liu, X.; Biggar, K.K.; Li, S.S.; Newton, A.C.; O’Donoghue, P. Genetic code expansion and live cell imaging reveal that Thr-308 phosphorylation is irreplaceable and sufficient for Akt1 activity. J. Biol. Chem. 2018, 293, 10744–10756. [Google Scholar] [CrossRef] [Green Version]
- Mao, H.; Tang, Z.; Li, H.; Sun, B.; Tan, M.; Fan, S.; Zhu, Y.; Sun, Y. Neddylation inhibitor MLN4924 suppresses cilia formation by modulating AKT1. Protein Cell 2019, 10, 726–744. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, A.S., Jr. The NF-kappa B and I kappa B proteins: New discoveries and insights. Annu. Rev. Immunol. 1996, 14, 649–683. [Google Scholar] [CrossRef] [Green Version]
- Wang, V.Y.; Li, Y.; Kim, D.; Zhong, X.; Du, Q.; Ghassemian, M.; Ghosh, G. Bcl3 Phosphorylation by Akt, Erk2, and IKK Is Required for Its Transcriptional Activity. Mol. Cell 2017, 67, 484–497. [Google Scholar] [CrossRef] [PubMed]
- Avila, M.; Martinez-Juarez, A.; Ibarra-Sanchez, A.; Gonzalez-Espinosa, C. Lyn kinase controls TLR4-dependent IKK and MAPK activation modulating the activity of TRAF-6/TAK-1 protein complex in mast cells. Innate Immun. 2012, 18, 648–660. [Google Scholar] [CrossRef] [Green Version]
- Walker, L.R.; Hussein, H.A.M.; Akula, S.M. Subcellular fractionation method to study endosomal trafficking of Kaposi’s sarcoma-associated herpesvirus. Cell Biosci. 2016, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- López-Peláez, M.; Soria-Castro, I.; Boscá, L.; Fernández, M.; Alemany, S. Cot/tpl2 activity is required for TLR-induced activation of the Akt p70 S6k pathway in macrophages: Implications for NO synthase 2 expression. Eur. J. Immunol. 2011, 41, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Klein, D.C.; Skjesol, A.; Kers-Rebel, E.D.; Sherstova, T.; Sporsheim, B.; Egeberg, K.W.; Stokke, B.T.; Espevik, T.; Husebye, H. CD14, TLR4 and TRAM Show Different Trafficking Dynamics During LPS Stimulation. Traffic 2015, 16, 677–690. [Google Scholar] [CrossRef] [PubMed]
- Beutler, B. Inferences, questions and possibilities in Toll-like receptor signalling. Nature 2004, 430, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Pelka, K.; Bertheloot, D.; Reimer, E.; Phulphagar, K.; Schmidt, S.V.; Christ, A.; Stahl, R.; Watson, N.; Miyake, K.; Hacohen, N.; et al. The Chaperone UNC93B1 Regulates Toll-like Receptor Stability Independently of Endosomal TLR Transport. Immunity 2018, 48, 911–922. [Google Scholar] [CrossRef]
- You, X.; Xu, D.D.; Zhang, D.; Chen, J.; Gao, F.G. PYR-41 and Thalidomide Impair Dendritic Cell Cross-Presentation by Inhibiting Myddosome Formation and Attenuating the Endosomal Recruitments of p97 and Sec61 via NF-κB Inactivation. J. Immunol. Res. 2018, 2018, 5070573. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, D.D.; Hu, C.F.; You, X.; Lu, N.N.; Gao, F.G. Akt+ IKKα/β+ Rab5+ Signalosome Mediate the Endosomal Recruitment of Sec61 and Contribute to Cross-Presentation in Bone Marrow Precursor Cells. Vaccines 2020, 8, 539. https://doi.org/10.3390/vaccines8030539
Xu DD, Hu CF, You X, Lu NN, Gao FG. Akt+ IKKα/β+ Rab5+ Signalosome Mediate the Endosomal Recruitment of Sec61 and Contribute to Cross-Presentation in Bone Marrow Precursor Cells. Vaccines. 2020; 8(3):539. https://doi.org/10.3390/vaccines8030539
Chicago/Turabian StyleXu, Dan Dan, Chun Fang Hu, Xiang You, Nan Nan Lu, and Feng Guang Gao. 2020. "Akt+ IKKα/β+ Rab5+ Signalosome Mediate the Endosomal Recruitment of Sec61 and Contribute to Cross-Presentation in Bone Marrow Precursor Cells" Vaccines 8, no. 3: 539. https://doi.org/10.3390/vaccines8030539