
Myelin Peptide–Mannan Conjugate Multiple Sclerosis Vaccines: Conjugation Efficacy and Stability of Vaccine Ingredient

 ,
,  , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
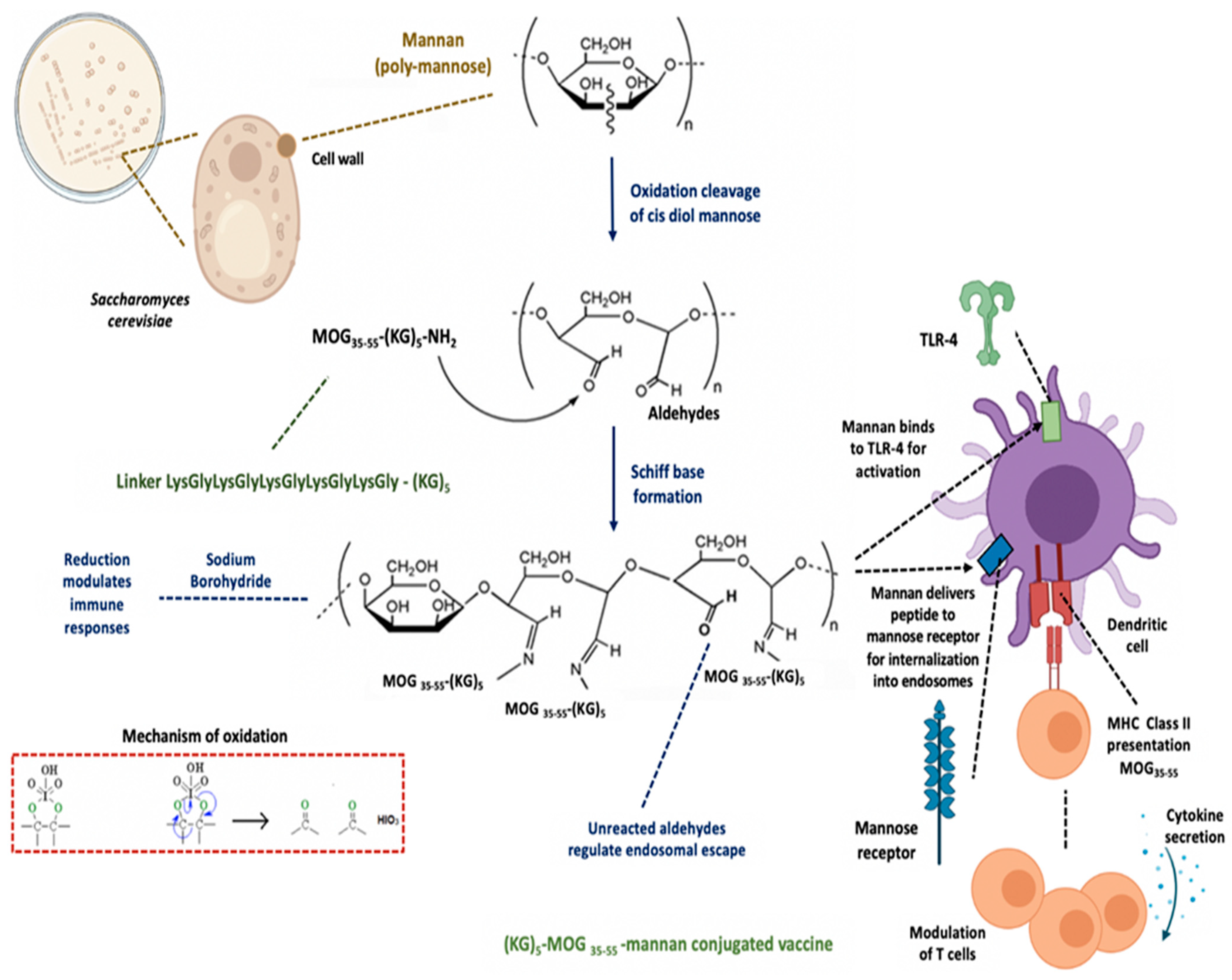
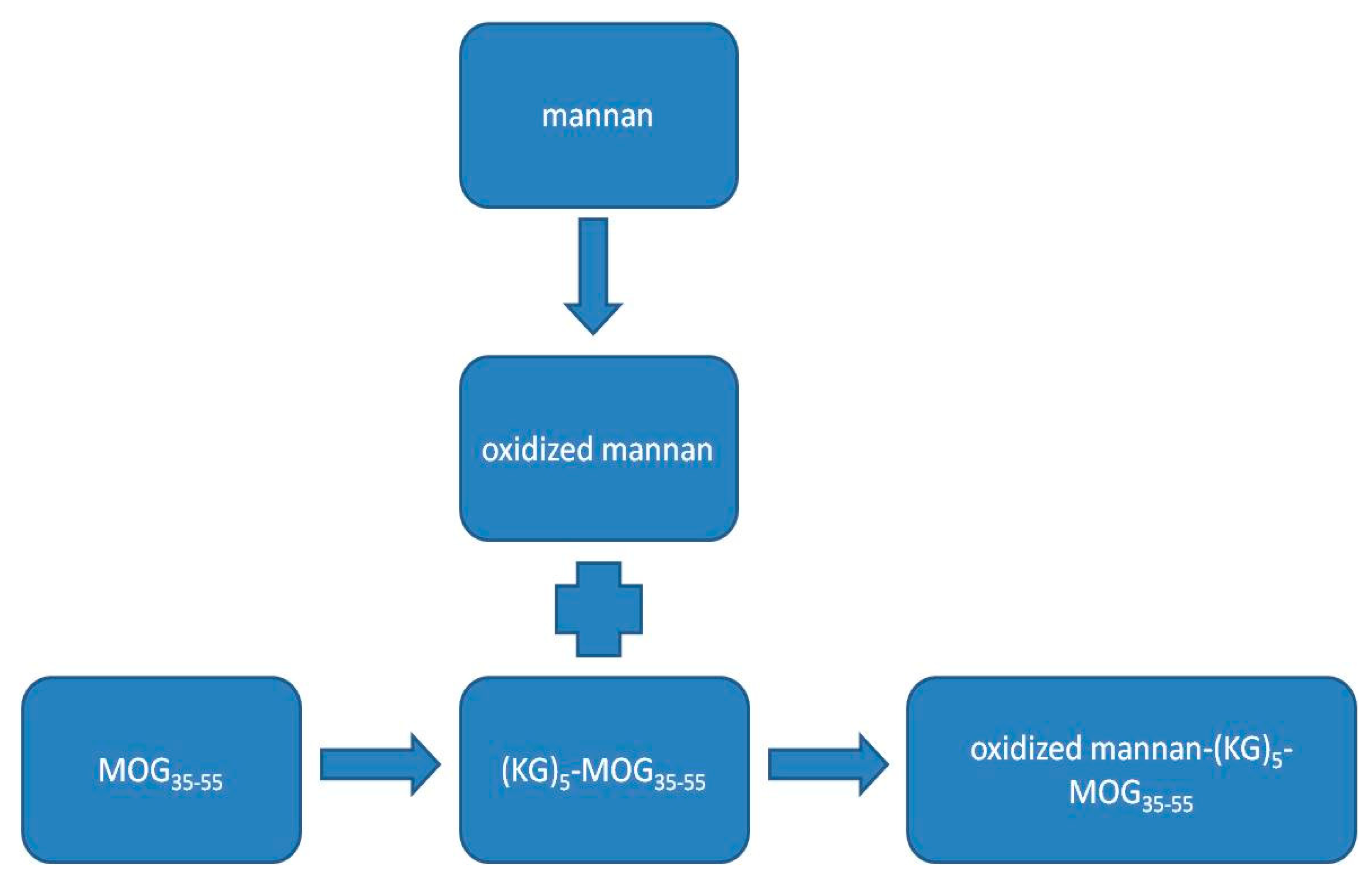
2.1. Preparation of the Mannan–Peptide Conjugate
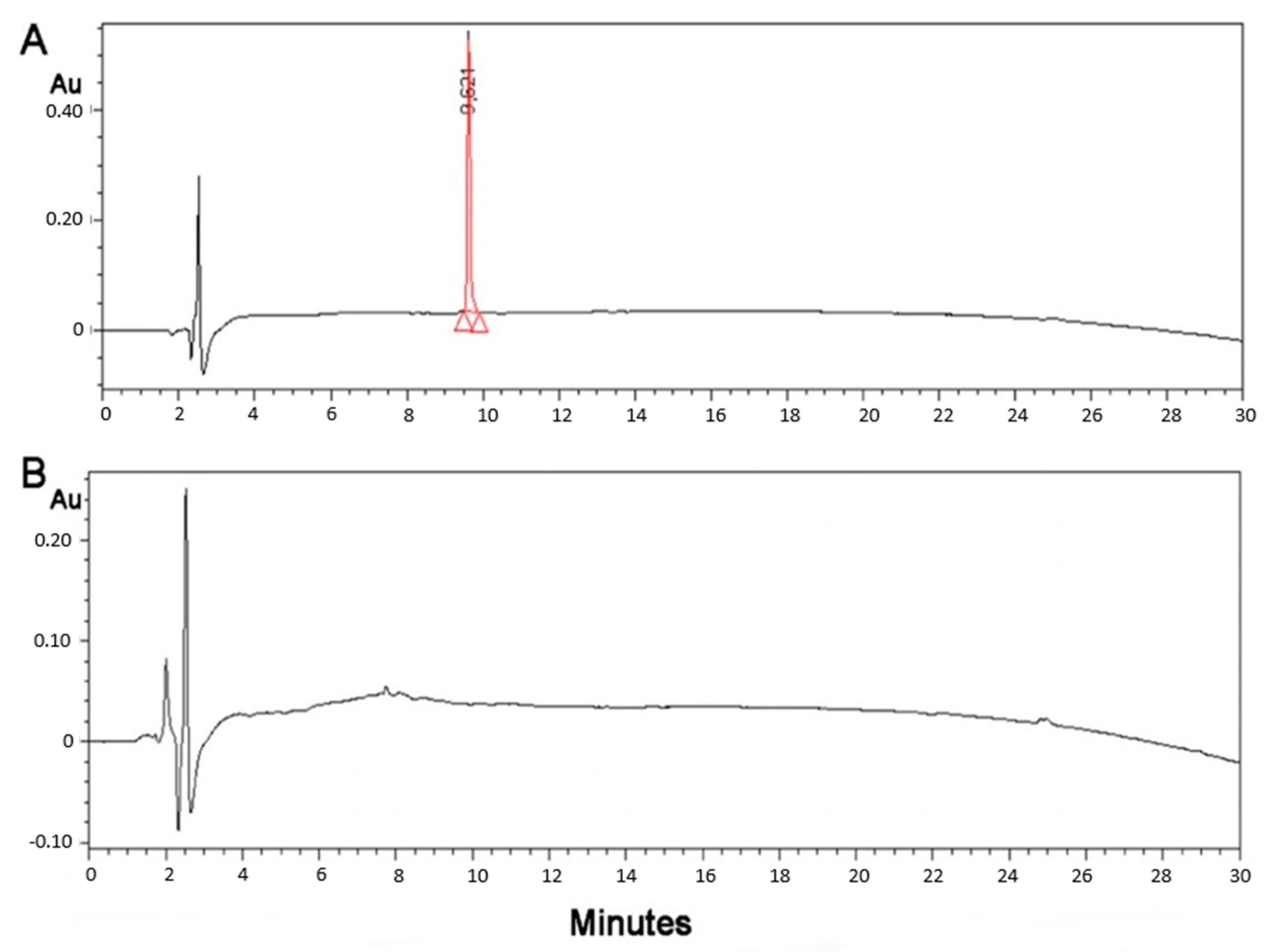
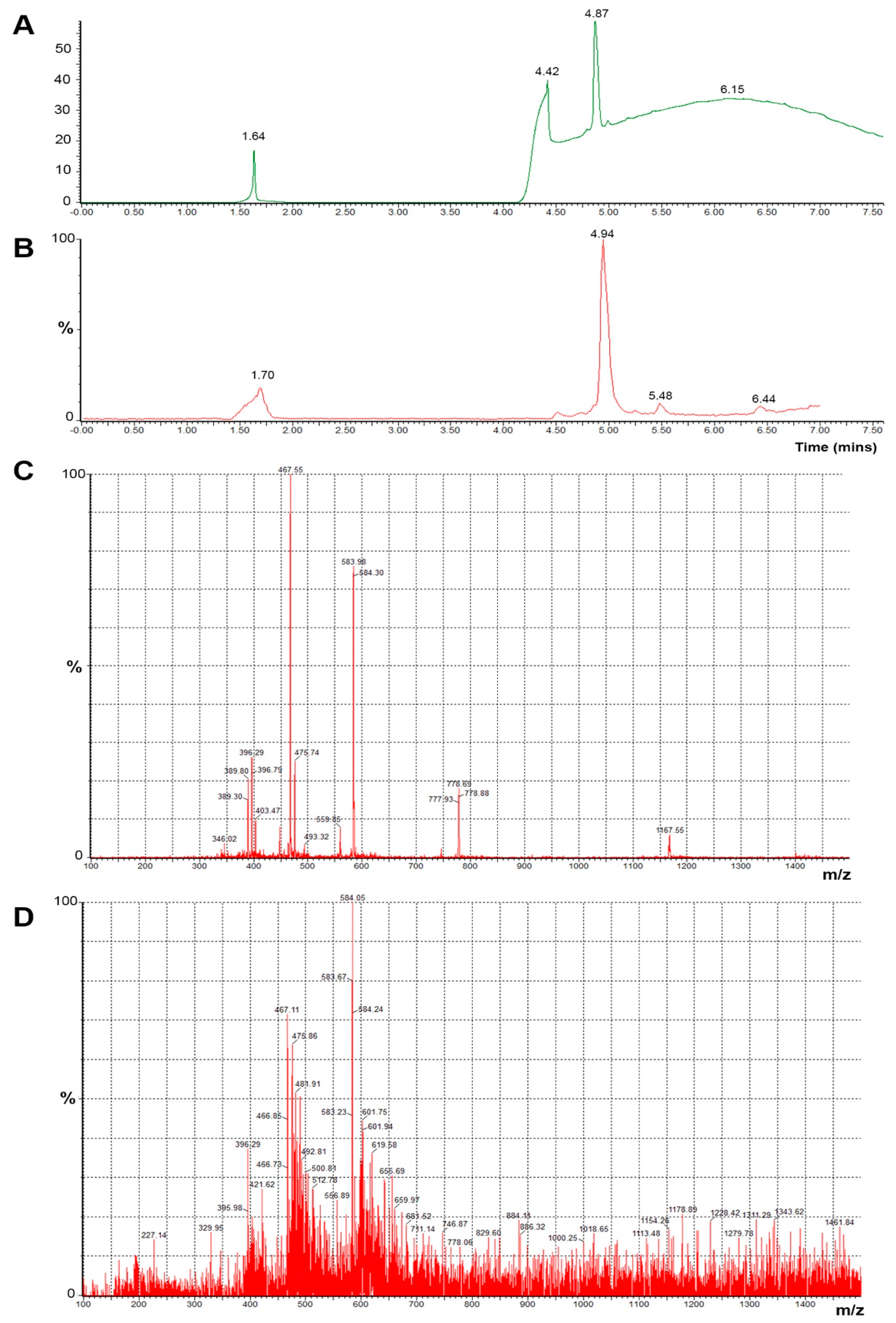
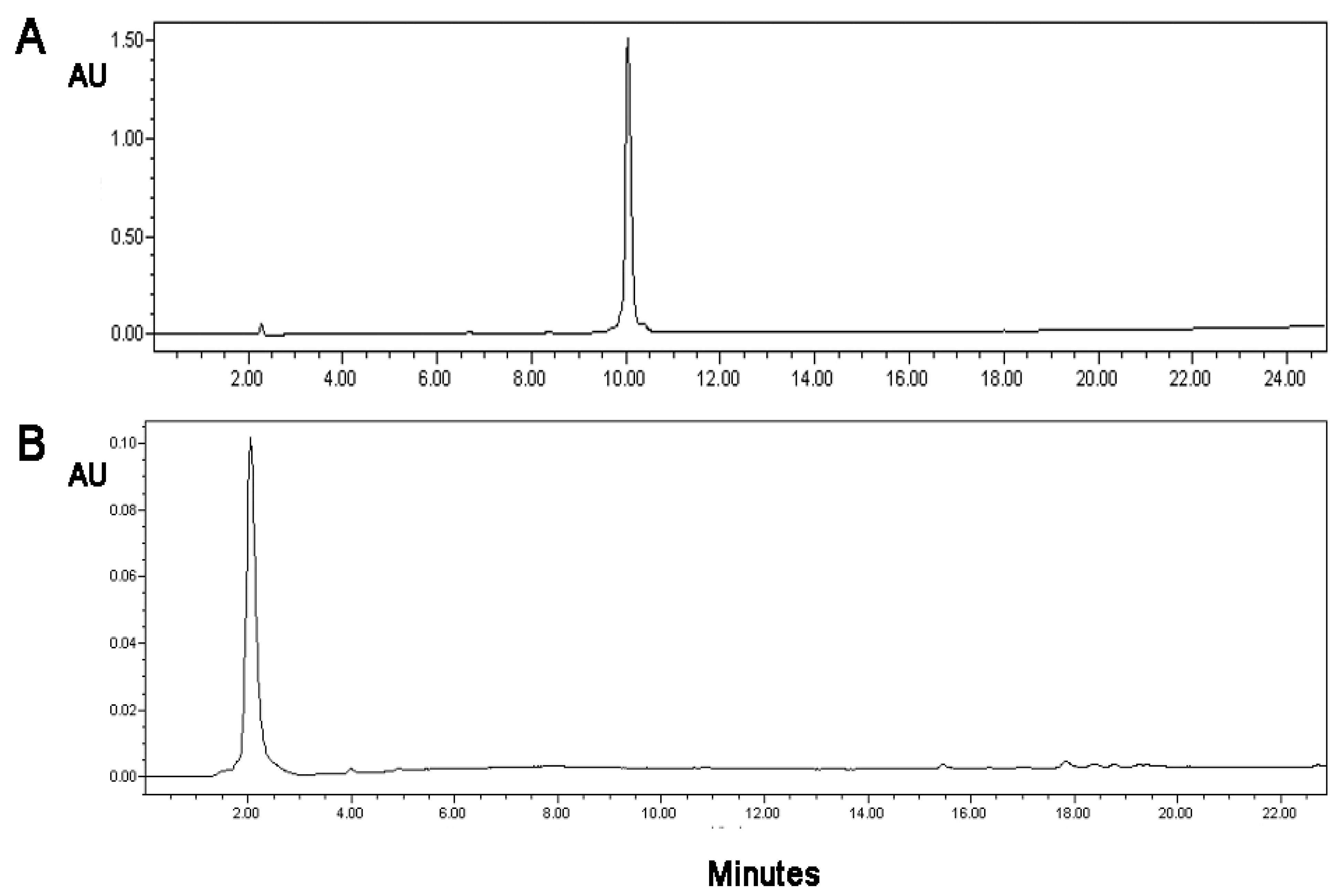
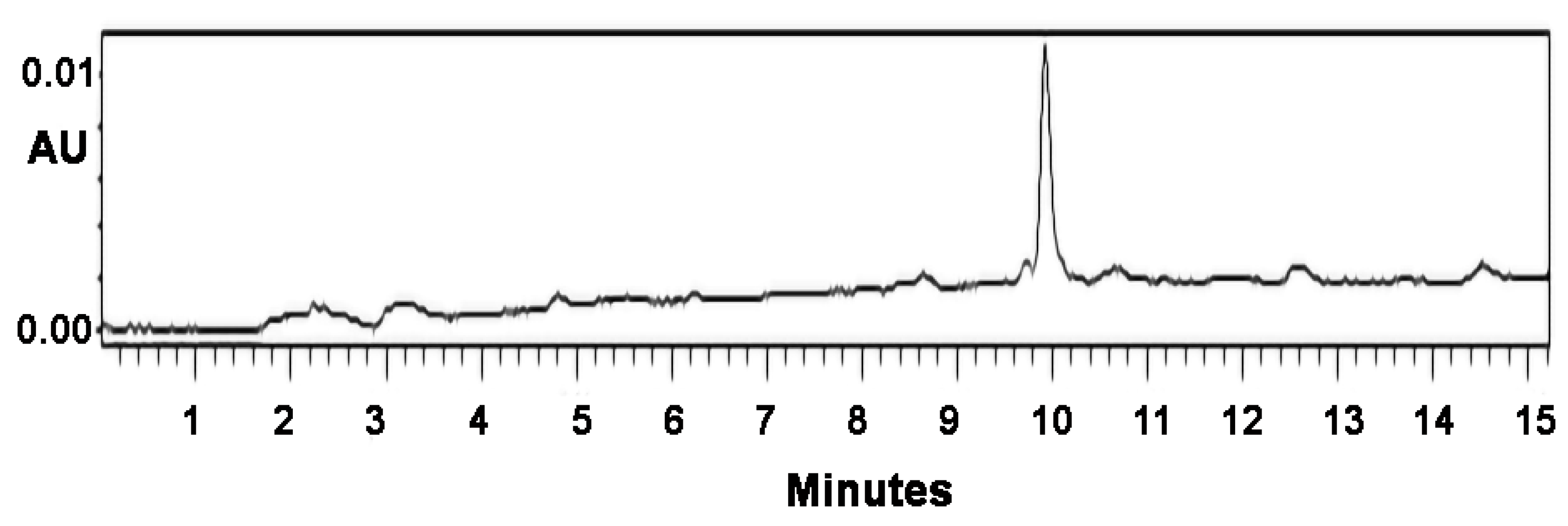
2.2. Monitoring of the Conjugation by HPLC
2.3. Preparation of the Electrodes and Monitoring of the Conjugation Cyclic Voltammetry
3. Results
3.1. Conjugation of the (KG)5-MOG35–55 Peptide with Mannan
3.2. Attempt to Conjugate MOG35–55 Peptide with Oxidized Mannan without (KG)5 Was Not Successful
3.3. Enzymatic Cleavage of the Peptide (KG)5-MOG35–55 Results to the Fragment MOG37–55
3.4. Cyclic Voltammetry Showing Conjugation of (KG)5-MOG35–55 to Oxidized Mannan
3.5. Stability of the Conjugate and Its Sensitivity
4. Discussion
4.1. Mannan as a Carrier
4.2. The Role of the Linker (KG)5 for Conjugation
4.3. Main Properties of Peptide Conjugate Excipients
4.4. Mannan–Peptide Conjugate: Requirements for Immunoregulation of Dendritic Cells
4.5. Antigen Presentation by Tolerogenic Dendritic Cells Using the MOG35–55–Mannan Conjugate
4.6. Biological Activity of Mannan–Peptide Conjugates
4.7. Advantages of HPLC Methods Confirming Conjugation and the Stability of Peptide Conjugates
4.8. Analysis by HPLC Techniques
4.9. Cyclic Voltammetry
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steinman, M.D.L. Multiple Sclerosis: A Coordinated Immunological Attack against Myelin in the Central Nervous System. Cell 1996, 85, 299–302. [Google Scholar] [CrossRef] [Green Version]
- Steinman, L. Multiple sclerosis: A two-stage disease. Nat. Immunol. 2001, 2, 762–764. [Google Scholar] [CrossRef] [PubMed]
- Kappos, L.; Comi, G.; Panitch, H.; Oger, J.; Antel, J.; Conlon, P.; Steinman, L.; Comi, G.; Kappos, L.; Oger, J.; et al. Induction of a non-encephalitogenic type 2 T helper-cell autoimmune response in multiple sclerosis after administration of an altered peptide ligand in a placebo-controlled, randomized phase II trial. Nat. Med. 2000, 6, 1176–1182. [Google Scholar] [CrossRef]
- Piatek, P.; Namiecinska, M.; Domowicz, M.; Przygodzka, P.; Wieczorek, M.; Michlewska, S.; Lewkowicz, N.; Tarkowski, M.; Lewkowicz, P. MS CD49d+CD154+ Lymphocytes Reprogram Oligodendrocytes into Immune Reactive Cells Affecting CNS Regeneration. Cells 2019, 8, 1508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apostolopoulos, V.; Matsoukas, J. Advances in Multiple Sclerosis Research–Series I. Brain Sci. 2020, 10, 795. [Google Scholar] [CrossRef]
- Gasperi, C.; Andlauer, T.F.M.; Keating, A.; Knier, B.; Klein, A.; Pernpeintner, V.; Lichtner, P.; Gold, R.; Zipp, F.; Then Bergh, F.; et al. Genetic determinants of the humoral immune response in MS. Neurol.—Neuroimmunol. Neuroinflammation 2020, 7, e827. [Google Scholar] [CrossRef]
- Häusler, D.; Hajiyeva, Z.; Traub, J.W.; Zamvil, S.S.; Lalive, P.H.; Brück, W.; Weber, M.S. Glatiramer acetate immune modulates B-cell antigen presentation in treatment of MS. Neurol.—Neuroimmunol. Neuroinflammation 2020, 7, e698. [Google Scholar] [CrossRef] [Green Version]
- Haines, J.D.; Inglese, M.; Casaccia, P. Axonal Damage in Multiple Sclerosis. Mt. Sinai J. Med. A J. Transl. Pers. Med. 2011, 78, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Ziello, A.; Scavone, C.; Di Battista, M.E.; Salvatore, S.; Di Giulio Cesare, D.; Moreggia, O.; Allegorico, L.; Sagnelli, A.; Barbato, S.; Manzo, V.; et al. Influenza Vaccine Hesitancy in Patients with Multiple Sclerosis: A Monocentric Observational Study. Brain Sci. 2021, 11, 890. [Google Scholar] [CrossRef]
- Horwitz, D.A.; Fahmy, T.M.; Piccirillo, C.A.; La Cava, A. Rebalancing Immune Homeostasis to Treat Autoimmune Diseases. Trends Immunol. 2019, 40, 888–908. [Google Scholar] [CrossRef]
- Iberg, C.A.; Hawiger, D. Natural and Induced Tolerogenic Dendritic Cells. J. Immunol. 2020, 204, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Ciric, B.; Zhang, G.X.; Rostami, A. Immunotherapy using lipopolysaccharide-stimulated bone marrow-derived dendritic cells to treat experimental autoimmune encephalomyelitis. Clin. Exp. Immunol. 2014, 178, 447–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benedek, G.; Meza-Romero, R.; Jordan, K.; Keenlyside, L.; Offner, H.; Vandenbark, A.A. HLA-DRα1-mMOG-35-55 treatment of experimental autoimmune encephalomyelitis reduces CNS inflammation, enhances M2 macrophage frequency, and promotes neuroprotection. J. Neuroinflammation 2015, 12, 123. [Google Scholar] [CrossRef] [Green Version]
- Tabansky, I.; Keskin, D.B.; Watts, D.; Petzold, C.; Funaro, M.; Sands, W.; Wright, P.; Yunis, E.J.; Najjar, S.; Diamond, B.; et al. Targeting DEC-205−DCIR2+ dendritic cells promotes immunological tolerance in proteolipid protein-induced experimental autoimmune encephalomyelitis. Mol. Med. 2018, 24, 17. [Google Scholar] [CrossRef] [Green Version]
- Boyko, A.; Therapontos, C.; Horakova, D.; Szilasiová, J.; Kalniņa, J.; Kolontareva, J.; Gross-Paju, K.; Selmaj, K.; Sereike, I.; Milo, R.; et al. Approaches and challenges in the diagnosis and management of secondary progressive multiple sclerosis: A Central Eastern European perspective from healthcare professionals. Mult. Scler. Relat. Disord. 2021, 50, 102778. [Google Scholar] [CrossRef]
- Lee, J.Y.; Taghian, K.; Petratos, S. Axonal degeneration in multiple sclerosis: Can we predict and prevent permanent disability? Acta Neuropathol. Commun. 2014, 2, 97. [Google Scholar] [CrossRef]
- Lublin, F.D.; Reingold, S.C.; Cohen, J.A.; Cutter, G.R.; Sorensen, P.S.; Thompson, A.J.; Wolinsky, J.S.; Balcer, L.J.; Banwell, B.; Barkhof, F.; et al. Defining the clinical course of multiple sclerosis: The 2013 revisions. Neurology 2014, 83, 278–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsara, M.; Deraos, G.; Tselios, T.; Matsoukas, M.-T.; Friligou, I.; Matsoukas, J.; Apostolopoulos, V. Design and Synthesis of a Cyclic Double Mutant Peptide (cyclo(87−99)[A91,A96]MBP87−99) Induces Altered Responses in Mice after Conjugation to Mannan: Implications in the Immunotherapy of Multiple Sclerosis. J. Med. Chem. 2008, 52, 214–218. [Google Scholar] [CrossRef]
- Katsara, M.; Yuriev, E.; Ramsland, P.A.; Deraos, G.; Tselios, T.; Matsoukas, J.; Apostolopoulos, V. Mannosylation of mutated MBP83–99 peptides diverts immune responses from Th1 to Th2. Mol. Immunol. 2008, 45, 3661–3670. [Google Scholar] [CrossRef]
- Dagkonaki, A.; Avloniti, M.; Evangelidou, M.; Papazian, I.; Kanistras, I.; Tseveleki, V.; Lampros, F.; Tselios, T.; Jensen, L.T.; Möbius, W.; et al. Mannan-MOG35-55 Reverses Experimental Autoimmune Encephalomyelitis, Inducing a Peripheral Type 2 Myeloid Response, Reducing CNS Inflammation, and Preserving Axons in Spinal Cord Lesions. Front. Immunol. 2020, 11, 575451. [Google Scholar] [CrossRef]
- Vassilaros, S.; Tsibanis, A.; Tsikkinis, A.; Pietersz, G.A.; McKenzie, I.F.C.; Apostolopoulos, V. Up to 15-year clinical follow-up of a pilot Phase III immunotherapy study in stage II breast cancer patients using oxidized mannan–MUC1. Immunotherapy 2013, 5, 1177–1182. [Google Scholar] [CrossRef] [Green Version]
- Freedman, M.S.; Bar-Or, A.; Oger, J.; Traboulsee, A.; Patry, D.; Young, C.; Olsson, T.; Li, D.; Hartung, H.P.; Krantz, M.; et al. A phase III study evaluating the efficacy and safety of MBP8298 in secondary progressive MS. Neurology 2011, 77, 1551–1560. [Google Scholar] [CrossRef] [PubMed]
- Warren, K.G.; Catz, I.; Ferenczi, L.Z.; Krantz, M.J. Intravenous synthetic peptide MBP8298 delayed disease progression in an HLA Class II-defined cohort of patients with progressive multiple sclerosis: Results of a 24-month double-blind placebo-controlled clinical trial and 5 years of follow-up treatment. Eur. J. Neurol. 2006, 13, 887–895. [Google Scholar] [CrossRef]
- Deskoulidis, E.; Petrouli, S.; Apostolopoulos, V.; Matsoukas, J.; Topoglidis, E. The Use of Electrochemical Voltammetric Techniques and High-Pressure Liquid Chromatography to Evaluate Conjugation Efficiency of Multiple Sclerosis Peptide-Carrier Conjugates. Brain Sci. 2020, 10, 577. [Google Scholar] [CrossRef]
- Drapeau, G.R.; Boily, Y.; Houmard, J. Purification and properties of an extracellular protease of Staphylococcus aureus. J. Biol. Chem. 1972, 247, 6720–6726. [Google Scholar] [CrossRef]
- Houmard, J.; Drapeau, G.R. Staphylococcal Protease: A Proteolytic Enzyme Specific for Glutamoyl Bonds. Proc. Natl. Acad. Sci. USA 1972, 69, 3506–3509. [Google Scholar] [CrossRef] [Green Version]
- Klein, T.; Eckhard, U.; Dufour, A.; Solis, N.; Overall, C.M. Proteolytic Cleavage—Mechanisms, Function, and “Omic” Approaches for a Near-Ubiquitous Posttranslational Modification. Chem. Rev. 2017, 118, 1137–1168. [Google Scholar] [CrossRef]
- Apostolopoulos, V.; Pietersz, G.A.; Tsibanis, A.; Tsikkinis, A.; Drakaki, H.; Loveland, B.E.; Piddlesden, S.J.; Plebanski, M.; Pouniotis, D.S.; Alexis, M.N.; et al. Pilot phase III immunotherapy study in early-stage breast cancer patients using oxidized mannan-MUC1 [ISRCTN71711835]. Breast Cancer Res. 2006, 8, R27. [Google Scholar] [CrossRef] [PubMed]
- Rodi, M.; Dimisianos, N.; de Lastic, A.-L.; Sakellaraki, P.; Deraos, G.; Matsoukas, J.; Papathanasopoulos, P.; Mouzaki, A. Regulatory Cell Populations in Relapsing-Remitting Multiple Sclerosis (RRMS) Patients: Effect of Disease Activity and Treatment Regimens. Int. J. Mol. Sci. 2016, 17, 1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseveleki, V.; Tselios, T.; Kanistras, I.; Koutsoni, O.; Karamita, M.; Vamvakas, S.-S.; Apostolopoulos, V.; Dotsika, E.; Matsoukas, J.; Lassmann, H.; et al. Mannan-conjugated myelin peptides prime non-pathogenic Th1 and Th17 cells and ameliorate experimental autoimmune encephalomyelitis. Exp. Neurol. 2015, 267, 254–267. [Google Scholar] [CrossRef] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsoukas, J.; Deraos, G.; Kelaidonis, K.; Hossain, M.K.; Feehan, J.; Tzakos, A.G.; Matsoukas, E.; Topoglidis, E.; Apostolopoulos, V. Myelin Peptide–Mannan Conjugate Multiple Sclerosis Vaccines: Conjugation Efficacy and Stability of Vaccine Ingredient. Vaccines 2021, 9, 1456. https://doi.org/10.3390/vaccines9121456
Matsoukas J, Deraos G, Kelaidonis K, Hossain MK, Feehan J, Tzakos AG, Matsoukas E, Topoglidis E, Apostolopoulos V. Myelin Peptide–Mannan Conjugate Multiple Sclerosis Vaccines: Conjugation Efficacy and Stability of Vaccine Ingredient. Vaccines. 2021; 9(12):1456. https://doi.org/10.3390/vaccines9121456
Chicago/Turabian StyleMatsoukas, John, George Deraos, Kostas Kelaidonis, Md Kamal Hossain, Jack Feehan, Andreas G. Tzakos, Elizabeth Matsoukas, Emmanuel Topoglidis, and Vasso Apostolopoulos. 2021. "Myelin Peptide–Mannan Conjugate Multiple Sclerosis Vaccines: Conjugation Efficacy and Stability of Vaccine Ingredient" Vaccines 9, no. 12: 1456. https://doi.org/10.3390/vaccines9121456