
Long-Term Anti-Bacterial Immunity against Systemic Infection by Salmonella enterica Serovar Typhimurium Elicited by a GMMA-Based Vaccine

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. STmGMMA/Alhydrogel
2.2. Animals
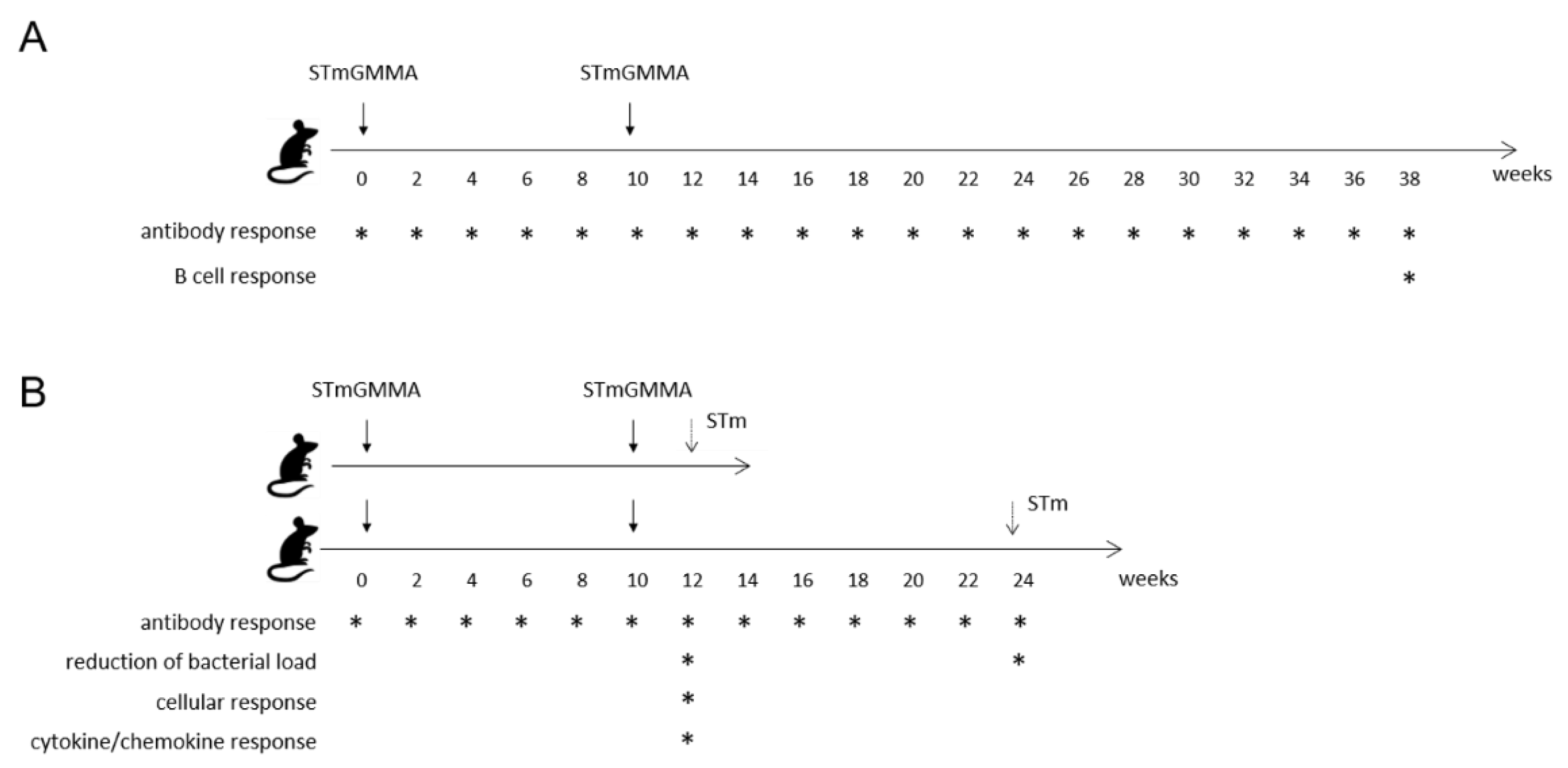
2.3. Immunizations
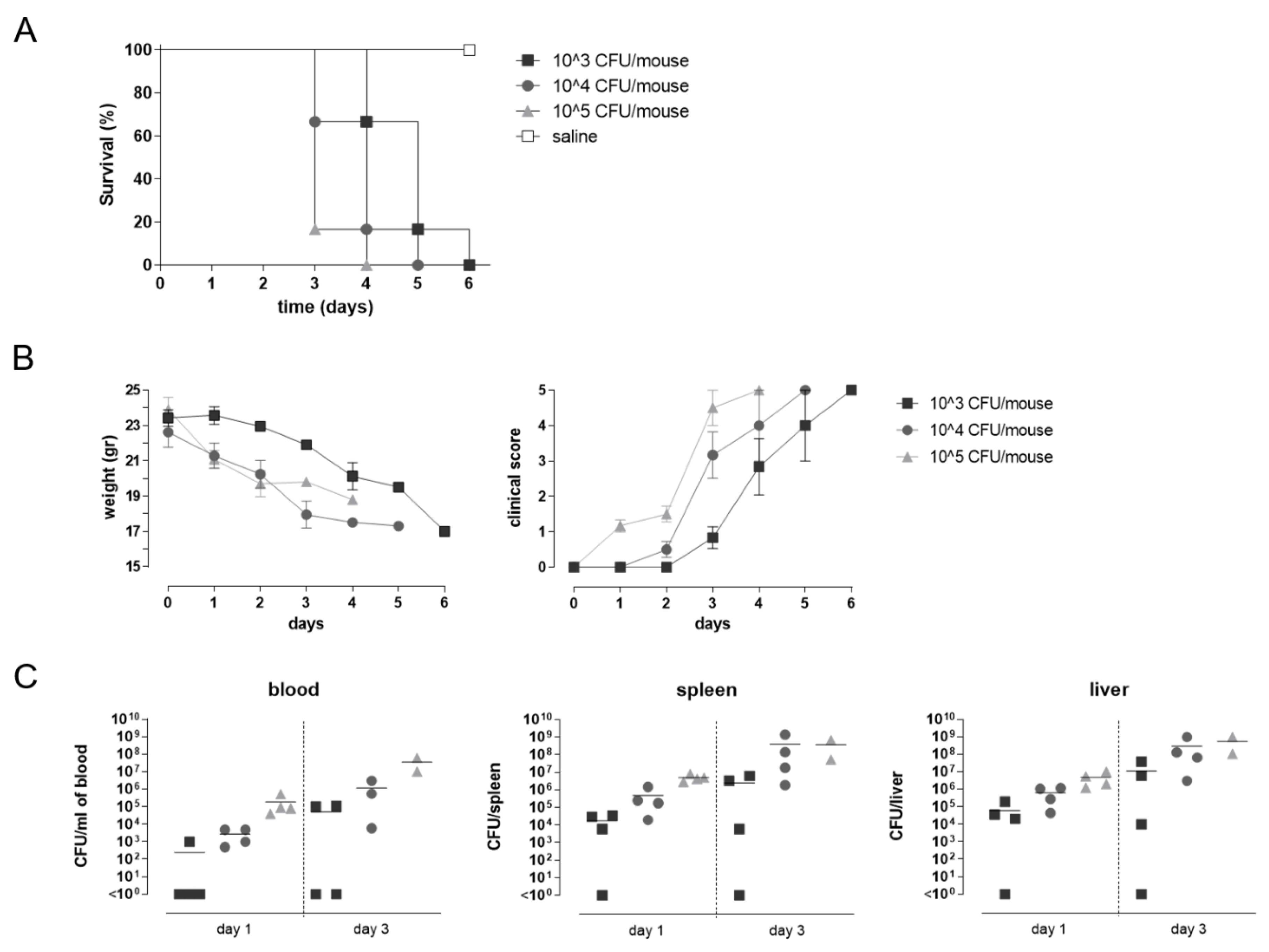
2.4. Challenge Model
2.5. Samples Collection and Treatment
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
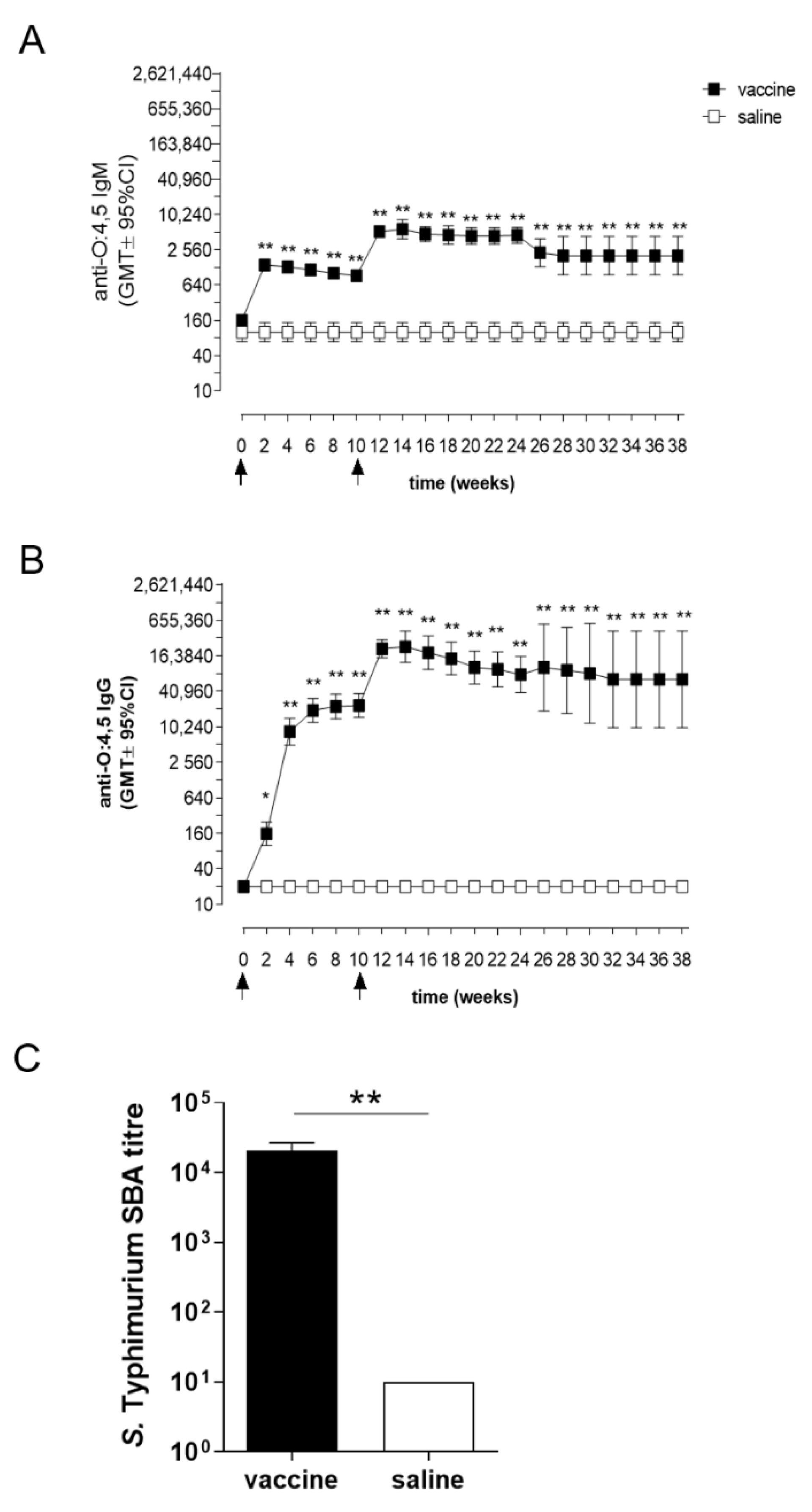
2.7. Serum Bactericidal Activity (SBA) Assay
2.8. Multiparametric Flow Cytometric Analysis
2.9. Memory B Cell ELISpot Assay
2.10. Multiplex Cytokine and Chemokine Assay
2.11. Statistical Analysis
3. Results
3.1. Serum Antibody Response
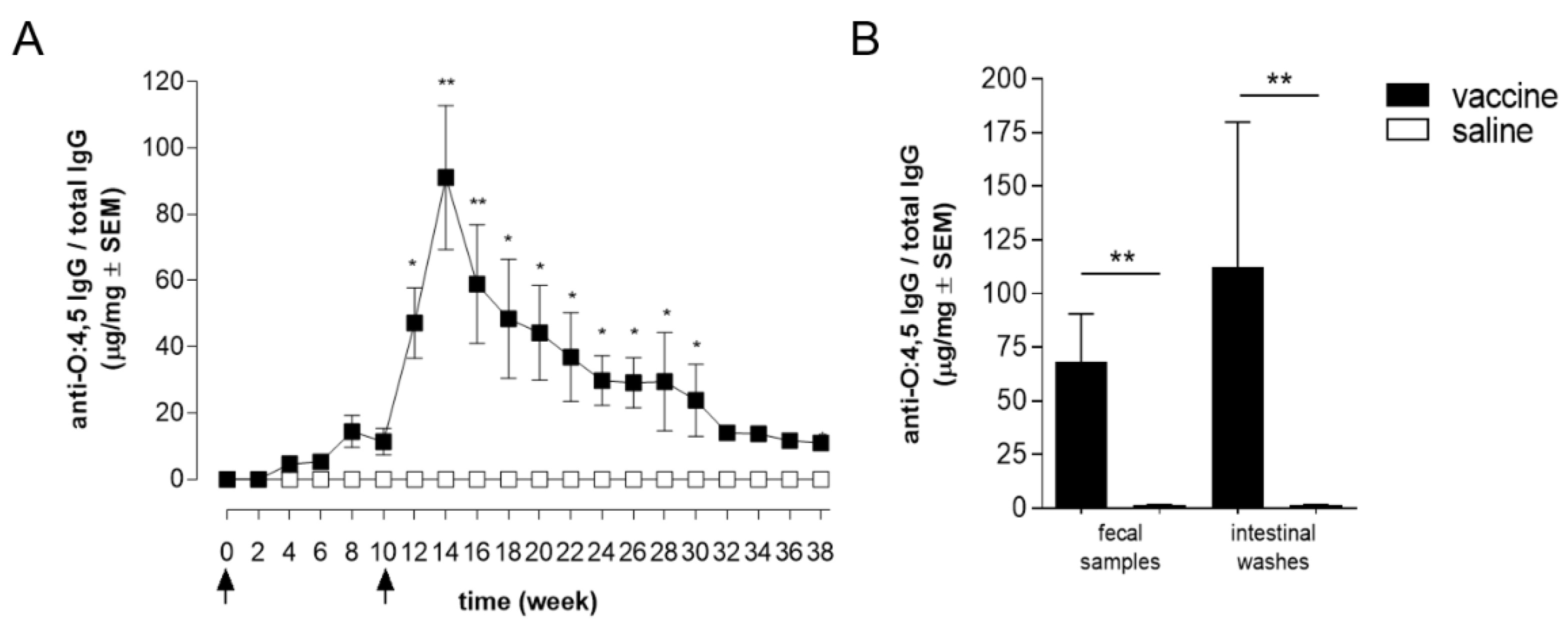
3.2. Intestinal Antibody Response
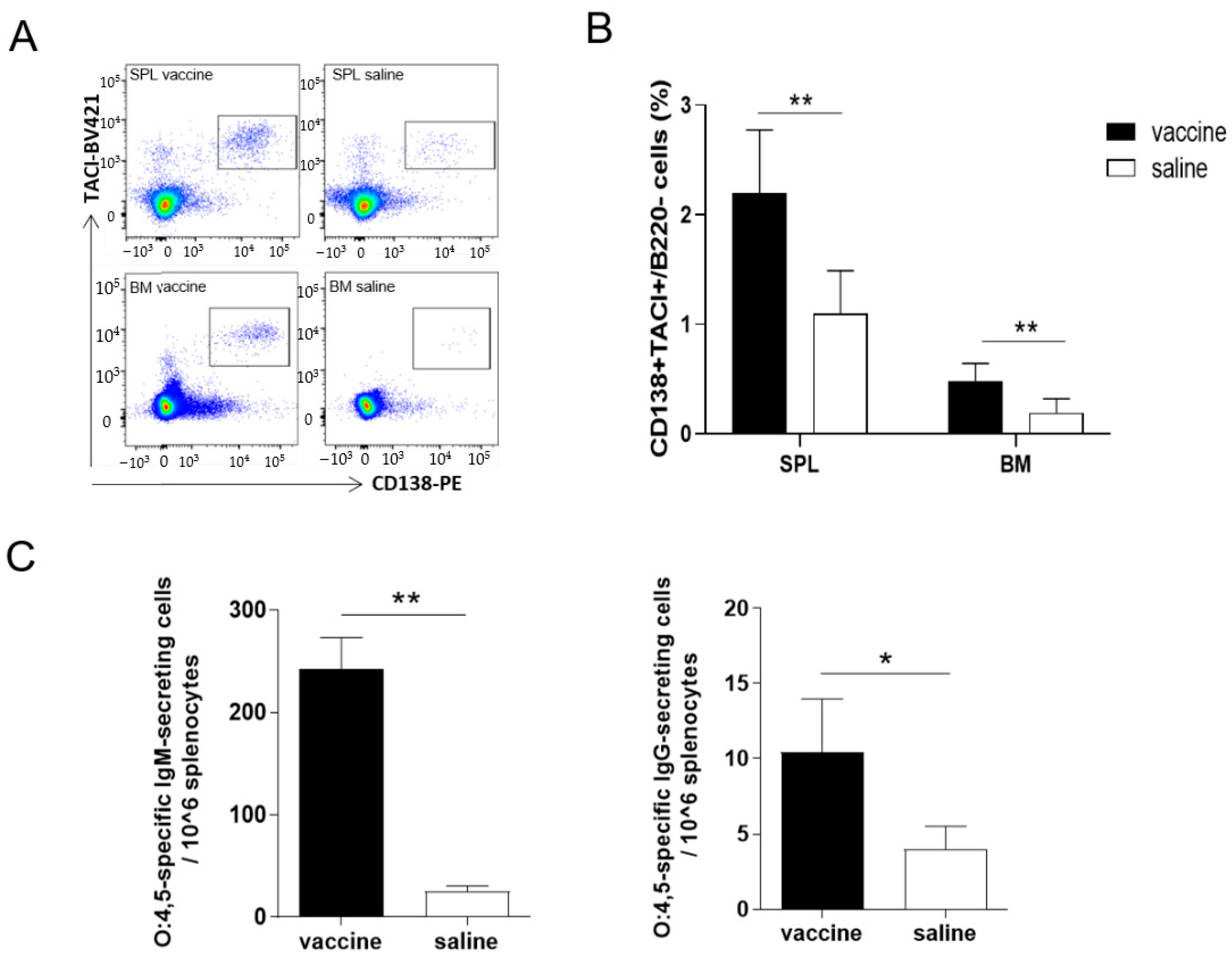
3.3. B Cell Response
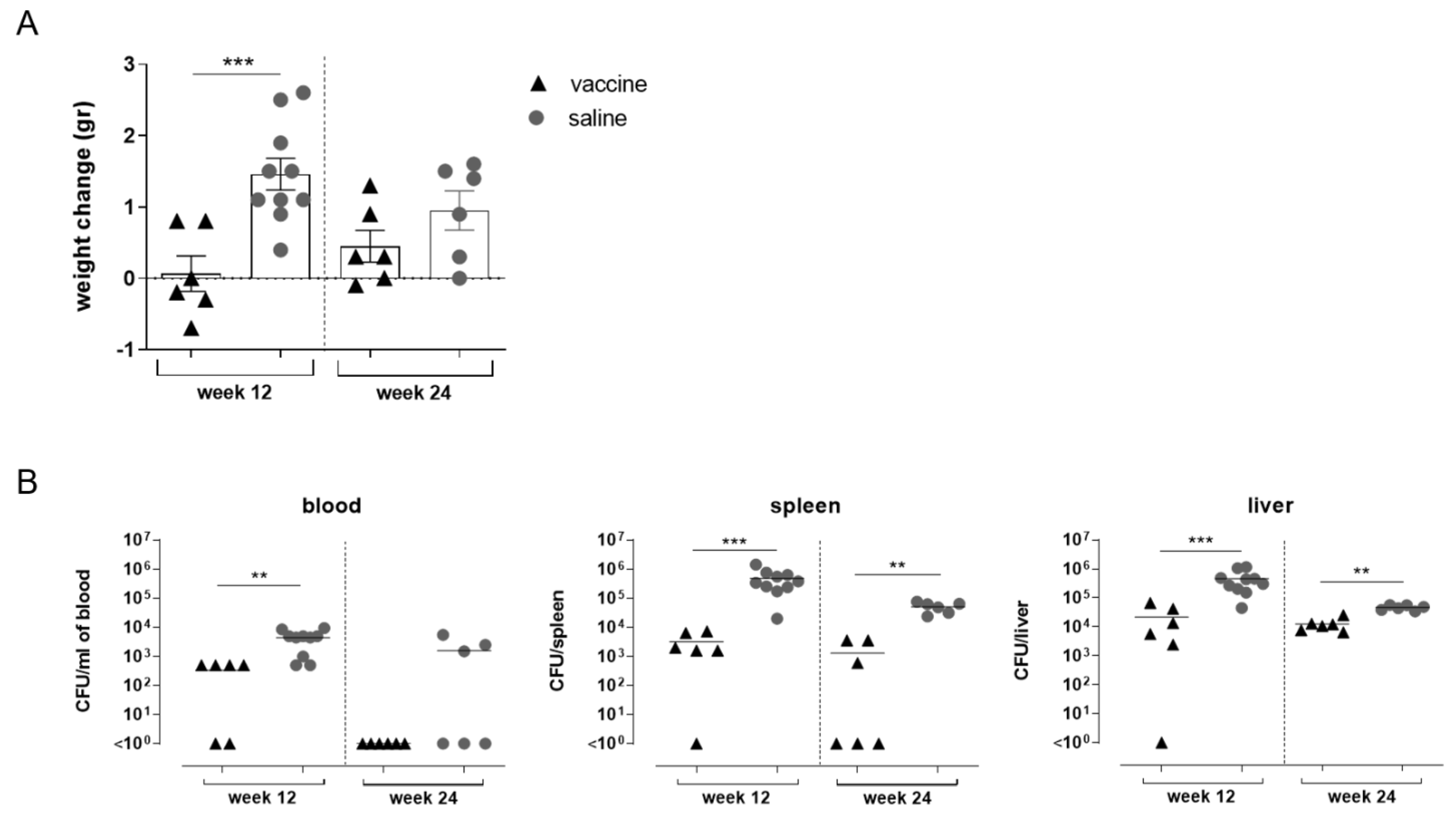
3.4. Impact of STmGMMA/Alhydrogel Immunization on Challenge Outcomes
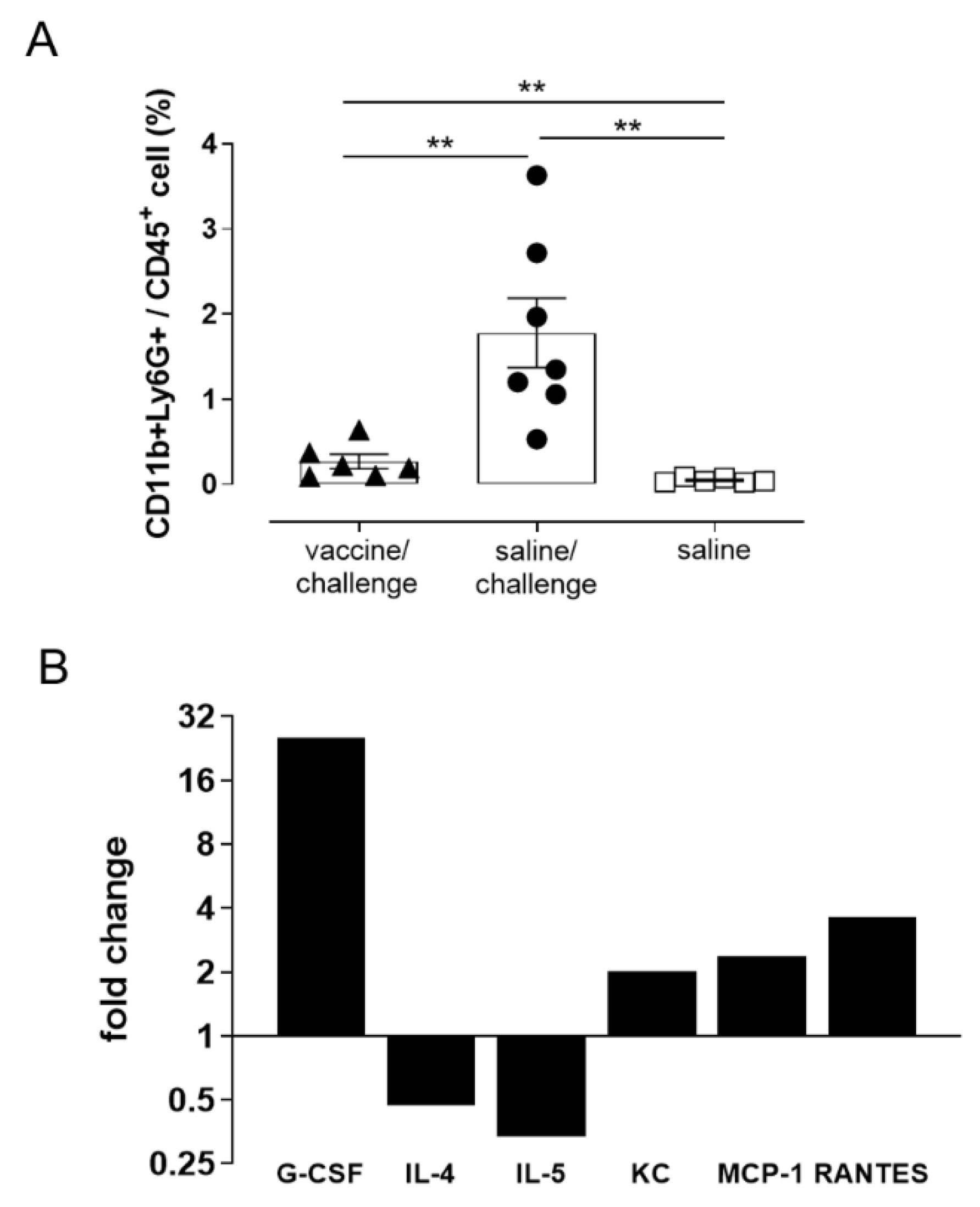
3.5. Characterization of Blood Cell Populations
3.6. Analysis of Cytokine and Chemokine Production
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2017 Non-Typhoidal Salmonella Invasive Disease Collaborators. The Global Burden of Non-Typhoidal Salmonella Invasive Disease: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet Infect Dis. 2019, 19, 1312–1324. [Google Scholar] [CrossRef]
- Ao, T.T.; Feasey, N.A.; Gordon, M.A.; Keddy, K.H.; Angulo, F.J.; Crump, J.A. Global Burden of Invasive Nontyphoidal Salmonella Disease, 2010. Emerging Infect. Dis. 2015, 21. [Google Scholar] [CrossRef] [PubMed]
- Feasey, N.A.; Dougan, G.; Kingsley, R.A.; Heyderman, R.S.; Gordon, M.A. Invasive Non-Typhoidal Salmonella Disease: An Emerging and Neglected Tropical Disease in Africa. Lancet 2012, 379, 2489–2499. [Google Scholar] [CrossRef]
- Gordon, M.A. Invasive Nontyphoidal Salmonella Disease: Epidemiology, Pathogenesis and Diagnosis. Curr. Opin. Infect. Dis. 2011, 24, 484–489. [Google Scholar] [CrossRef]
- Uche, I.V.; MacLennan, C.A.; Saul, A. A Systematic Review of the Incidence, Risk Factors and Case Fatality Rates of Invasive Nontyphoidal Salmonella (INTS) Disease in Africa (1966 to 2014). PLoS Negl. Trop. Dis. 2017, 11, e0005118. [Google Scholar] [CrossRef]
- Haselbeck, A.H.; Panzner, U.; Im, J.; Baker, S.; Meyer, C.G.; Marks, F. Current Perspectives on Invasive Nontyphoidal Salmonella Disease. Curr. Opin. Infect. Dis. 2017, 30, 498–503. [Google Scholar] [CrossRef]
- Crump, J.A.; Heyderman, R.S. A Perspective on Invasive Salmonella Disease in Africa. Clin. Infect. Dis. 2015, 61 (Suppl. 4), S235–S240. [Google Scholar] [CrossRef]
- Crump, J.A.; Sjölund-Karlsson, M.; Gordon, M.A.; Parry, C.M. Epidemiology, Clinical Presentation, Laboratory Diagnosis, Antimicrobial Resistance, and Antimicrobial Management of Invasive Salmonella Infections. Clin. Microbiol. Rev. 2015, 28, 901–937. [Google Scholar] [CrossRef] [PubMed]
- Kirk, M.D.; Pires, S.M.; Black, R.E.; Caipo, M.; Crump, J.A.; Devleesschauwer, B.; Döpfer, D.; Fazil, A.; Fischer-Walker, C.L.; Hald, T.; et al. World Health Organization Estimates of the Global and Regional Disease Burden of 22 Foodborne Bacterial, Protozoal, and Viral Diseases, 2010: A Data Synthesis. PLoS Med. 2015, 12, e1001921. [Google Scholar] [CrossRef]
- MacLennan, C.A.; Levine, M.M. Invasive Nontyphoidal Salmonella Disease in Africa: Current Status. Expert Rev. Anti-Infect. Ther. 2013, 11, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, R.; Im, J.; Lee, J.-S.; Jeon, H.J.; Mogeni, O.D.; Kim, J.H.; Rakotozandrindrainy, R.; Baker, S.; Marks, F. The Global Burden and Epidemiology of Invasive Non-Typhoidal Salmonella Infections. Hum. Vaccines Immunother. 2019, 15, 1421–1426. [Google Scholar] [CrossRef] [PubMed]
- Feasey, N.A.; Gaskell, K.; Wong, V.; Msefula, C.; Selemani, G.; Kumwenda, S.; Allain, T.J.; Mallewa, J.; Kennedy, N.; Bennett, A.; et al. Rapid Emergence of Multidrug Resistant, H58-Lineage Salmonella Typhi in Blantyre, Malawi. PLoS Negl. Trop. Dis. 2015, 9, e0003748. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.A. Salmonella Infections in Immunocompromised Adults. J. Infect. 2008, 56, 413–422. [Google Scholar] [CrossRef]
- MacLennan, C.A.; Gilchrist, J.J.; Gordon, M.A.; Cunningham, A.F.; Cobbold, M.; Goodall, M.; Kingsley, R.A.; van Oosterhout, J.J.G.; Msefula, C.L.; Mandala, W.L.; et al. Dysregulated Humoral Immunity to Nontyphoidal Salmonella in HIV-Infected African Adults. Science 2010, 328, 508–512. [Google Scholar] [CrossRef]
- Enwere, G.; Biney, E.; Cheung, Y.B.; Zaman, S.M.A.; Okoko, B.; Oluwalana, C.; Vaughan, A.; Greenwood, B.; Adegbola, R.; Cutts, F.T. Epidemiologic and Clinical Characteristics of Community-Acquired Invasive Bacterial Infections in Children Aged 2–29 Months in The Gambia. Pediatr. Infect. Dis. J. 2006, 25, 700–705. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, J.J.; MacLennan, C.A. Invasive Nontyphoidal Salmonella Disease in Africa. EcoSal Plus 2019, 8. [Google Scholar] [CrossRef]
- MacLennan, C.A.; Martin, L.B.; Micoli, F. Vaccines against Invasive Salmonella Disease: Current Status and Future Directions. Hum. Vaccines Immunother. 2014, 10, 1478–1493. [Google Scholar] [CrossRef]
- Tennant, S.M.; MacLennan, C.A.; Simon, R.; Martin, L.B.; Khan, M.I. Nontyphoidal Salmonella Disease: Current Status of Vaccine Research and Development. Vaccine 2016, 34, 2907–2910. [Google Scholar] [CrossRef]
- Micoli, F.; Rondini, S.; Alfini, R.; Lanzilao, L.; Necchi, F.; Negrea, A.; Rossi, O.; Brandt, C.; Clare, S.; Mastroeni, P.; et al. Comparative Immunogenicity and Efficacy of Equivalent Outer Membrane Vesicle and Glycoconjugate Vaccines against Nontyphoidal Salmonella. Proc. Natl. Acad. Sci. USA 2018, 115, 10428–10433. [Google Scholar] [CrossRef]
- Kis, Z.; Shattock, R.; Shah, N.; Kontoravdi, C. Emerging Technologies for Low-Cost, Rapid Vaccine Manufacture. Biotechnol. J. 2019, 14, e1800376. [Google Scholar] [CrossRef] [PubMed]
- Colwell, D.E.; Michalek, S.M.; Briles, D.E.; Jirillo, E.; McGhee, J.R. Monoclonal Antibodies to Salmonella Lipopolysaccharide: Anti-O-Polysaccharide Antibodies Protect C3H Mice against Challenge with Virulent Salmonella Typhimurium. J. Immunol. 1984, 133, 950–957. [Google Scholar]
- Mastroeni, P.; Rossi, O. Immunology, Epidemiology and Mathematical Modelling towards a Better Understanding of Invasive Non-Typhoidal Salmonella Disease and Rational Vaccination Approaches. Expert Rev. Vaccines 2016, 15, 1545–1555. [Google Scholar] [CrossRef]
- Nagy, G.; Pál, T. Lipopolysaccharide: A Tool and Target in Enterobacterial Vaccine Development. Biol. Chem. 2008, 389, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Rondini, S.; Lanzilao, L.; Necchi, F.; O’Shaughnessy, C.M.; Micoli, F.; Saul, A.; MacLennan, C.A. Invasive African Salmonella Typhimurium Induces Bactericidal Antibodies against O-Antigens. Microb. Pathog. 2013, 63, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Simon, R.; Levine, M.M. Glycoconjugate Vaccine Strategies for Protection against Invasive Salmonella Infections. Hum. Vaccines Immunother. 2012, 8, 494–498. [Google Scholar] [CrossRef] [PubMed]
- Medaglini, D.; Andersen, P.; Rappuoli, R. Editorial: Advanced Immunization Technologies for Next Generation Vaccines. Front. Immunol. 2020, 11, 878. [Google Scholar] [CrossRef]
- Rappuoli, R.; Medaglini, D. ADITEC: Joining Forces for next-Generation Vaccines. Sci. Transl. Med. 2012, 4, 128cm4. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Medaglini, D.; De Azero, M.R.; Leroy, O.; Bietrix, F.; Denoel, P. Innovation Partnership for a Roadmap on Vaccines in Europe (IPROVE): A Vision for the Vaccines of Tomorrow. Vaccine 2018, 36, 1136–1145. [Google Scholar] [CrossRef]
- Pettini, E.; Pastore, G.; Fiorino, F.; Medaglini, D.; Ciabattini, A. Short or Long Interval between Priming and Boosting: Does It Impact on the Vaccine Immunogenicity? Vaccines 2021, 9, 289. [Google Scholar] [CrossRef]
- Fiorino, F.; Ciabattini, A.; Rondini, S.; Pozzi, G.; Martin, L.B.; Medaglini, D. Immunization with the Conjugate Vaccine Vi-CRM197 against Salmonella Typhi Induces Vi-Specific Mucosal and Systemic Immune Responses in Mice. Vaccine 2012, 30, 6111–6114. [Google Scholar] [CrossRef][Green Version]
- Pollard, A.J.; Perrett, K.P.; Beverley, P.C. Maintaining Protection against Invasive Bacteria with Protein-Polysaccharide Conjugate Vaccines. Nat. Rev. Immunol. 2009, 9, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Szu, S.C.; Klugman, K.P.; Hunt, S. Re-Examination of Immune Response and Estimation of Anti-Vi IgG Protective Threshold against Typhoid Fever-Based on the Efficacy Trial of Vi Conjugate in Young Children. Vaccine 2014, 32, 2359–2363. [Google Scholar] [CrossRef]
- Fiorino, F.; Rondini, S.; Micoli, F.; Lanzilao, L.; Alfini, R.; Mancini, F.; MacLennan, C.A.; Medaglini, D. Immunogenicity of a Bivalent Adjuvanted Glycoconjugate Vaccine against Salmonella Typhimurium and Salmonella Enteritidis. Front. Immunol. 2017, 8, 168. [Google Scholar] [CrossRef] [PubMed]
- Rondini, S.; Micoli, F.; Lanzilao, L.; Gavini, M.; Alfini, R.; Brandt, C.; Clare, S.; Mastroeni, P.; Saul, A.; MacLennan, C.A. Design of Glycoconjugate Vaccines against Invasive African Salmonella Enterica Serovar Typhimurium. Infect. Immun. 2015, 83, 996–1007. [Google Scholar] [CrossRef] [PubMed]
- Svenson, S.B.; Nurminen, M.; Lindberg, A.A. Artificial Salmonella Vaccines: O-Antigenic Oligosaccharide-Protein Conjugates Induce Protection against Infection with Salmonella Typhimurium. Infect. Immun. 1979, 25, 863–872. [Google Scholar] [CrossRef]
- Berlanda Scorza, F.; Colucci, A.M.; Maggiore, L.; Sanzone, S.; Rossi, O.; Ferlenghi, I.; Pesce, I.; Caboni, M.; Norais, N.; Di Cioccio, V.; et al. High Yield Production Process for Shigella Outer Membrane Particles. PLoS ONE 2012, 7, e35616. [Google Scholar] [CrossRef] [PubMed]
- Rossi, O.; Pesce, I.; Giannelli, C.; Aprea, S.; Caboni, M.; Citiulo, F.; Valentini, S.; Ferlenghi, I.; MacLennan, C.A.; D’Oro, U.; et al. Modulation of Endotoxicity of Shigella Generalized Modules for Membrane Antigens (GMMA) by Genetic Lipid A Modifications: Relative Activation of TLR4 and TLR2 Pathways in Different Mutants. J. Biol. Chem. 2014, 289, 24922–24935. [Google Scholar] [CrossRef]
- Rossi, O.; Caboni, M.; Negrea, A.; Necchi, F.; Alfini, R.; Micoli, F.; Saul, A.; MacLennan, C.A.; Rondini, S.; Gerke, C. Toll-Like Receptor Activation by Generalized Modules for Membrane Antigens from Lipid A Mutants of Salmonella Enterica Serovars Typhimurium and Enteritidis. Clin. Vaccine Immunol. 2016, 23, 304–314. [Google Scholar] [CrossRef]
- Gerke, C.; Colucci, A.M.; Giannelli, C.; Sanzone, S.; Vitali, C.G.; Sollai, L.; Rossi, O.; Martin, L.B.; Auerbach, J.; Di Cioccio, V.; et al. Production of a Shigella Sonnei Vaccine Based on Generalized Modules for Membrane Antigens (GMMA), 1790GAHB. PLoS ONE 2015, 10, e0134478. [Google Scholar] [CrossRef]
- Meloni, E.; Colucci, A.M.; Micoli, F.; Sollai, L.; Gavini, M.; Saul, A.; Di Cioccio, V.; MacLennan, C.A. Simplified Low-Cost Production of O-Antigen from Salmonella Typhimurium Generalized Modules for Membrane Antigens (GMMA). J. Biotechnol. 2015, 198, 46–52. [Google Scholar] [CrossRef]
- Launay, O.; Lewis, D.J.M.; Anemona, A.; Loulergue, P.; Leahy, J.; Sciré, A.S.; Maugard, A.; Marchetti, E.; Zancan, S.; Huo, Z.; et al. Safety Profile and Immunologic Responses of a Novel Vaccine Against Shigella Sonnei Administered Intramuscularly, Intradermally and Intranasally: Results From Two Parallel Randomized Phase 1 Clinical Studies in Healthy Adult Volunteers in Europe. EBioMedicine 2017, 22, 164–172. [Google Scholar] [CrossRef]
- Launay, O.; Ndiaye, A.G.W.; Conti, V.; Loulergue, P.; Sciré, A.S.; Landre, A.M.; Ferruzzi, P.; Nedjaai, N.; Schütte, L.D.; Auerbach, J.; et al. Booster Vaccination With GVGH Shigella Sonnei 1790GAHB GMMA Vaccine Compared to Single Vaccination in Unvaccinated Healthy European Adults: Results From a Phase 1 Clinical Trial. Front. Immunol. 2019, 10, 335. [Google Scholar] [CrossRef]
- Obiero, C.W.; Ndiaye, A.G.W.; Sciré, A.S.; Kaunyangi, B.M.; Marchetti, E.; Gone, A.M.; Schütte, L.D.; Riccucci, D.; Auerbach, J.; Saul, A.; et al. A Phase 2a Randomized Study to Evaluate the Safety and Immunogenicity of the 1790GAHB Generalized Modules for Membrane Antigen Vaccine against Shigella Sonnei Administered Intramuscularly to Adults from a Shigellosis-Endemic Country. Front. Immunol. 2017, 8, 1884. [Google Scholar] [CrossRef]
- Kingsley, R.A.; Msefula, C.L.; Thomson, N.R.; Kariuki, S.; Holt, K.E.; Gordon, M.A.; Harris, D.; Clarke, L.; Whitehead, S.; Sangal, V.; et al. Epidemic Multiple Drug Resistant Salmonella Typhimurium Causing Invasive Disease in Sub-Saharan Africa Have a Distinct Genotype. Genome Res. 2009, 19, 2279–2287. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, C.A.; Gondwe, E.N.; Msefula, C.L.; Kingsley, R.A.; Thomson, N.R.; White, S.A.; Goodall, M.; Pickard, D.J.; Graham, S.M.; Dougan, G.; et al. The Neglected Role of Antibody in Protection against Bacteremia Caused by Nontyphoidal Strains of Salmonella in African Children. J. Clin. Investig. 2008, 118, 1553–1562. [Google Scholar] [CrossRef] [PubMed]
- Fiorino, F.; Pettini, E.; Pozzi, G.; Medaglini, D.; Ciabattini, A. Prime-Boost Strategies in Mucosal Immunization Affect Local IgA Production and the Type of Th Response. Front. Immunol. 2013, 4, 128. [Google Scholar] [CrossRef] [PubMed]
- Lanzilao, L.; Stefanetti, G.; Saul, A.; MacLennan, C.A.; Micoli, F.; Rondini, S. Strain Selection for Generation of O-Antigen-Based Glycoconjugate Vaccines against Invasive Nontyphoidal Salmonella Disease. PLoS ONE 2015, 10, e0139847. [Google Scholar] [CrossRef] [PubMed]
- Hammarlof, D.L.; Kroger, C.; Owen, S.V.; Canals, R.; Lacharme-Lora, L.; Wenner, N.; Schager, A.E.; Wells, T.J.; Henderson, I.R.; Wigley, P.; et al. Role of a Single Noncoding Nucleotide in the Evolution of an Epidemic African Clade of Salmonella. Proc. Natl. Acad. Sci. USA 2018, 115, E2614–E2623. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, A.D.; Rosenstreich, D.L.; Scher, I.; Campbell, G.H.; MacDermott, R.P.; Formal, S.B. Genetic Control of Susceptibility to Salmonella Typhimurium in Mice: Role of the LPS Gene. J. Immunol. 1980, 124, 20–24. [Google Scholar]
- Song, J.; Willinger, T.; Rongvaux, A.; Eynon, E.E.; Stevens, S.; Manz, M.G.; Flavell, R.A.; Galán, J.E. A Mouse Model for the Human Pathogen Salmonella Typhi. Cell Host Microbe 2010, 8, 369–376. [Google Scholar] [CrossRef]
- Mastroeni, P.; Grant, A.J. Spread of Salmonella Enterica in the Body during Systemic Infection: Unravelling Host and Pathogen Determinants. Expert Rev. Mol. Med. 2011, 13. [Google Scholar] [CrossRef]
- Gondwe, E.N.; Molyneux, M.E.; Goodall, M.; Graham, S.M.; Mastroeni, P.; Drayson, M.T.; MacLennan, C.A. Importance of Antibody and Complement for Oxidative Burst and Killing of Invasive Nontyphoidal Salmonella by Blood Cells in Africans. Proc. Natl. Acad. Sci. USA 2010, 107, 3070–3075. [Google Scholar] [CrossRef]
- Nyirenda, T.S.; Gilchrist, J.J.; Feasey, N.A.; Glennie, S.J.; Bar-Zeev, N.; Gordon, M.A.; MacLennan, C.A.; Mandala, W.L.; Heyderman, R.S. Sequential Acquisition of T Cells and Antibodies to Nontyphoidal Salmonella in Malawian Children. J. Infect. Dis. 2014, 210, 56–64. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, C.A. Vaccines for Low-Income Countries. Semin. Immunol. 2013, 25, 114–123. [Google Scholar] [CrossRef] [PubMed][Green Version]
- MacLennan, C.A.; Saul, A. Vaccines against Poverty. Proc. Natl. Acad. Sci. USA 2014, 111, 12307–12312. [Google Scholar] [CrossRef] [PubMed]
- Mancini, F.; Gasperini, G.; Rossi, O.; Aruta, M.G.; Raso, M.M.; Alfini, R.; Biagini, M.; Necchi, F.; Micoli, F. Dissecting the Contribution of O-Antigen and Proteins to the Immunogenicity of Shigella Sonnei Generalized Modules for Membrane Antigens (GMMA). Sci. Rep. 2021, 11, 906. [Google Scholar] [CrossRef] [PubMed]
- Goh, Y.S.; Clare, S.; Micoli, F.; Saul, A.; Mastroeni, P.; MacLennan, C.A. Monoclonal Antibodies of a Diverse Isotype Induced by an O-Antigen Glycoconjugate Vaccine Mediate In Vitro and In Vivo Killing of African Invasive Nontyphoidal Salmonella. Infect. Immun. 2015, 83, 3722–3731. [Google Scholar] [CrossRef]
- Trebicka, E.; Jacob, S.; Pirzai, W.; Hurley, B.P.; Cherayil, B.J. Role of Antilipopolysaccharide Antibodies in Serum Bactericidal Activity against Salmonella Enterica Serovar Typhimurium in Healthy Adults and Children in the United States. Clin. Vaccine Immunol. 2013, 20, 1491–1498. [Google Scholar] [CrossRef]
- Cunningham, A.F.; Gaspal, F.; Serre, K.; Mohr, E.; Henderson, I.R.; Scott-Tucker, A.; Kenny, S.M.; Khan, M.; Toellner, K.-M.; Lane, P.J.L.; et al. Salmonella Induces a Switched Antibody Response without Germinal Centers That Impedes the Extracellular Spread of Infection. J. Immunol. 2007, 178, 6200–6207. [Google Scholar] [CrossRef]
- Gil-Cruz, C.; Bobat, S.; Marshall, J.L.; Kingsley, R.A.; Ross, E.A.; Henderson, I.R.; Leyton, D.L.; Coughlan, R.E.; Khan, M.; Jensen, K.T.; et al. The Porin OmpD from Nontyphoidal Salmonella Is a Key Target for a Protective B1b Cell Antibody Response. Proc. Natl. Acad. Sci. USA 2009, 106, 9803–9808. [Google Scholar] [CrossRef]
- Marshall, J.L.; Flores-Langarica, A.; Kingsley, R.A.; Hitchcock, J.R.; Ross, E.A.; López-Macías, C.; Lakey, J.; Martin, L.B.; Toellner, K.-M.; MacLennan, C.A.; et al. The Capsular Polysaccharide Vi from Salmonella Typhi Is a B1b Antigen. J. Immunol. 2012, 189, 5527–5532. [Google Scholar] [CrossRef]
- Secundino, I.; López-Macías, C.; Cervantes-Barragán, L.; Gil-Cruz, C.; Ríos-Sarabia, N.; Pastelin-Palacios, R.; Villasis-Keever, M.A.; Becker, I.; Puente, J.L.; Calva, E.; et al. Salmonella Porins Induce a Sustained, Lifelong Specific Bactericidal Antibody Memory Response. Immunology 2006, 117, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Langereis, J.D.; Henriet, S.S.; Kuipers, S.; Weemaes, C.M.R.; van der Burg, M.; de Jonge, M.I.; van der Flier, M. IgM Augments Complement Bactericidal Activity with Serum from a Patient with a Novel CD79a Mutation. J. Clin. Immunol. 2018, 38, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Robbins, J.B.; Kenny, K.; Suter, E. The isolation and biological activities of rabbit gamma M-and gamma G-anti-Salmonella typhimurium antibodies. J. Exp. Med. 1965, 122, 385–402. [Google Scholar] [CrossRef]
- Saxen, H.; Mäkelä, O. The Protective Capacity of Immune Sera in Experimental Mouse Salmonellosis Is Mainly Due to IgM Antibodies. Immunol. Lett. 1982, 5, 267–272. [Google Scholar] [CrossRef]
- Saxén, H. Mechanism of the Protective Action of Anti-Salmonella IgM in Experimental Mouse Salmonellosis. J. Gen. Microbiol. 1984, 130, 2277–2283. [Google Scholar] [CrossRef]
- Cunningham, A.F.; Flores-Langarica, A.; Bobat, S.; Dominguez Medina, C.C.; Cook, C.N.L.; Ross, E.A.; Lopez-Macias, C.; Henderson, I.R. B1b Cells Recognize Protective Antigens after Natural Infection and Vaccination. Front. Immunol. 2014, 5, 535. [Google Scholar] [CrossRef]
- Ly, K.T.; Casanova, J.E. Mechanisms of Salmonella Entry into Host Cells. Cell. Microbiol. 2007, 9, 2103–2111. [Google Scholar] [CrossRef]
- Trammell, R.A.; Toth, L.A. Markers for Predicting Death as an Outcome for Mice Used in Infectious Disease Research. Comp. Med. 2011, 61, 492–498. [Google Scholar]
- Pettini, E.; Fiorino, F.; Cuppone, A.M.; Iannelli, F.; Medaglini, D.; Pozzi, G. Interferon-γ from Brain Leukocytes Enhances Meningitis by Type 4 Streptococcus Pneumoniae. Front. Microbiol. 2015, 6, 1340. [Google Scholar] [CrossRef]
- Mastroeni, P.; Villarreal-Ramos, B.; Hormaeche, C.E. Adoptive Transfer of Immunity to Oral Challenge with Virulent Salmonellae in Innately Susceptible BALB/c Mice Requires Both Immune Serum and T Cells. Infect. Immun. 1993, 61, 3981–3984. [Google Scholar] [CrossRef] [PubMed]
- McSorley, S.J.; Jenkins, M.K. Antibody Is Required for Protection against Virulent but Not Attenuated Salmonella Enterica Serovar Typhimurium. Infect. Immun. 2000, 68, 3344–3348. [Google Scholar] [CrossRef]
- Uppington, H.; Menager, N.; Boross, P.; Wood, J.; Sheppard, M.; Verbeek, S.; Mastroeni, P. Effect of Immune Serum and Role of Individual Fcgamma Receptors on the Intracellular Distribution and Survival of Salmonella Enterica Serovar Typhimurium in Murine Macrophages. Immunology 2006, 119, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Nutt, S.L.; Hodgkin, P.D.; Tarlinton, D.M.; Corcoran, L.M. The Generation of Antibody-Secreting Plasma Cells. Nat. Rev. Immunol. 2015, 15, 160–171. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fiorino, F.; Pettini, E.; Koeberling, O.; Ciabattini, A.; Pozzi, G.; Martin, L.B.; Medaglini, D. Long-Term Anti-Bacterial Immunity against Systemic Infection by Salmonella enterica Serovar Typhimurium Elicited by a GMMA-Based Vaccine. Vaccines 2021, 9, 495. https://doi.org/10.3390/vaccines9050495
Fiorino F, Pettini E, Koeberling O, Ciabattini A, Pozzi G, Martin LB, Medaglini D. Long-Term Anti-Bacterial Immunity against Systemic Infection by Salmonella enterica Serovar Typhimurium Elicited by a GMMA-Based Vaccine. Vaccines. 2021; 9(5):495. https://doi.org/10.3390/vaccines9050495
Chicago/Turabian StyleFiorino, Fabio, Elena Pettini, Oliver Koeberling, Annalisa Ciabattini, Gianni Pozzi, Laura B. Martin, and Donata Medaglini. 2021. "Long-Term Anti-Bacterial Immunity against Systemic Infection by Salmonella enterica Serovar Typhimurium Elicited by a GMMA-Based Vaccine" Vaccines 9, no. 5: 495. https://doi.org/10.3390/vaccines9050495
APA StyleFiorino, F., Pettini, E., Koeberling, O., Ciabattini, A., Pozzi, G., Martin, L. B., & Medaglini, D. (2021). Long-Term Anti-Bacterial Immunity against Systemic Infection by Salmonella enterica Serovar Typhimurium Elicited by a GMMA-Based Vaccine. Vaccines, 9(5), 495. https://doi.org/10.3390/vaccines9050495