1. Introduction
Multiple sclerosis (MS) is an autoimmune disorder that impacts the central nervous system (CNS) and is characterized by gliosis, demyelination, the inflammation process, and the degeneration of nerve cells [
1]. The accumulation of demyelinating lesions in the grey and white matter of the brain/spinal cord is the pathological hallmark of MS [
2]. Young adults with MS, typically between the ages of 20 and 30, present with unilateral optic neuritis, partial myelitis, sensory abnormalities, or brainstem syndromes such as internuclear ophthalmoplegia. Worldwide, between 5 and 300 cases of MS per 100,000 people are reported, with a higher incidence in higher latitudes. The overall life expectancy is shorter than the population average (75.9 years vs. 83.4 years), and the risk of developing MS is higher in females than in males (approximately a 3:1 distribution between the genders) [
3]. An autoimmune process has long been hypothesized as a mediating factor in MS. Research on experimental autoimmune encephalomyelitis (EAE), an animal model for MS, has suggested a crucial role for T helper lymphocytes. Researchers have studied how activated T cell subtypes contribute to the pathogenesis of MS, focusing on the genetic factors linked to the major histocompatibility complex (MHC) class II locus and the inflammatory response in the affected area [
4]. Also, serum levels of interleukin-12 (IL-2), interleukin-4 (IL-4), interleukin-6 (IL-6), interleukin-13 (IL-13), interleukin-17 (IL-17), interleukin-21 (IL-21), interleukin-22 (IL-22), and interleukin-33 (IL-33) tend to be higher in MS patients in the active disease phase than in healthy controls and patients in remission, although interleukin-10 (IL-10) seems to help slow the disease’s progression. Moreover, certain gene variants of interleukin-2 receptor (IL-2R), IL-4, IL-6, IL-13, and IL-22 have been linked to the development of MS [
5].
MS is defined as an immune system malfunction resulting in immune cells infiltrating the CNS [
6]. After being activated outside the CNS, autoreactive T cells cross the blood–brain barrier (BBB) and are reactivated by nearby antigen-presenting cells. The release of proinflammatory cytokines activates microglia and astrocytes, attracts further inflammatory cells, and induces plasma cells to produce antibodies. This inflammatory process ultimately damages the tissue within the plaque [
7].
MS and signal transducer and activator of transcription 4 (STAT4) may be related since MS has been linked to immune system dysfunction [
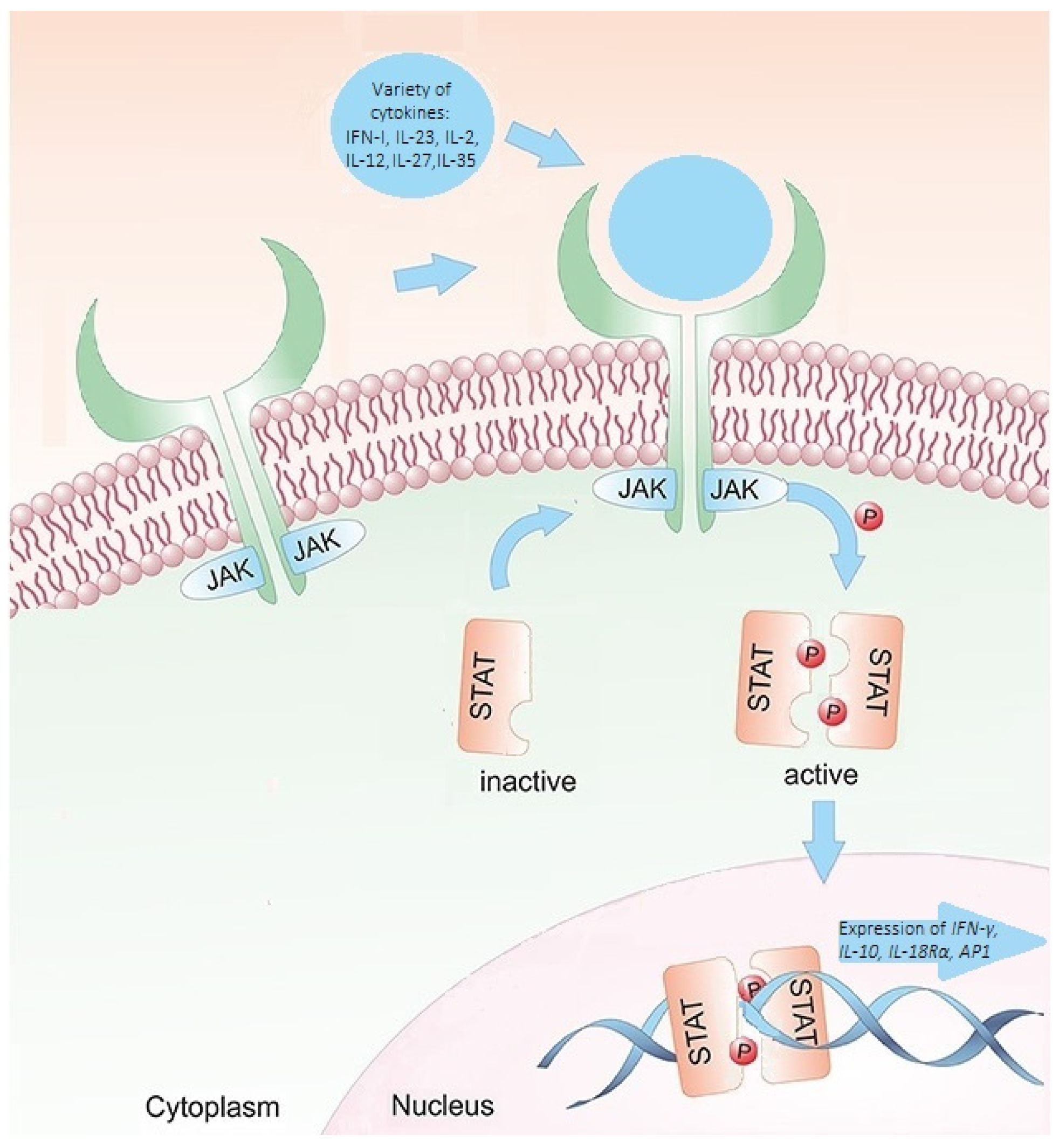
6]. Janus kinases (JAKs) are the proteins through which members of class I and class II cytokine receptor families deliver their signals. Activated JAKs phosphorylate the STATs. After phosphorylation, the STAT proteins undergo cytoplasmic dimerization before migrating to the nucleus, where they bind to deoxyribonucleic acid (DNA) regulatory elements and initiate gene transcription. The STAT signaling cascade is highly selective. A specific subset of genes dependent on STAT proteins is transcribed by any cytokine or combination of cytokines that exerts an effect [
8,
9]. Consequently, a variation in STAT4 expression or activity might impact the regular immune system’s response and function, resulting in immunosuppression or autoimmune disorders. STAT4 is a crucial modulator of the immunological response (
Figure 1) [
8]. In addition, the
STAT4 gene is responsible for relaying signals from interleukin-12 (IL-12), interleukin-23 (IL-23), and interferon type 1 (INF-1) in T cells and monocytes. These signals ultimately lead to the differentiation of type 1 T helper cells and type 17 T helper cells, monocyte activation, and the production of interferon-gamma (IFN-γ) [
10]. It is hypothesized that
STAT4 variants may influence the occurrence and function of immune cells involved in the pathogenesis of MS [
11].
It is important to note that genetic factors alone cannot explain the occurrence of MS, as environmental factors also play a significant role in the development of the disorder [
6,
14]. In addition, a positive family history increases the risk of MS for siblings of affected patients by around 30% compared to the general population. More than 200 genetic loci have been linked to MS by genome-wide association study (GWAS) [
15]. The epidemiology of MS suggests that smoking, low serum vitamin D levels, childhood obesity, and Epstein–Barr virus infection may contribute to the onset of the disease [
16]. Research on the connection between genetic and environmental factors in MS is ongoing to develop new prevention and therapeutic strategies. Overall, the link between the
STAT4 gene and MS suggests that dysregulation of the immune system plays a significant role in disease development [
6]. The basis of traditional MS treatment is immunomodulatory and anti-inflammatory medications. However, these measures cannot stop the degeneration of the nerve tissue. Neurologists should be aware of the latest findings on the development, pathophysiology, diagnosis, and treatment of MS [
17]. Further research is essential to fully clarify the role of the
STAT4 gene and STAT4 serum levels in MS and ascertain whether focusing on this gene could be an effective treatment strategy.
2. Materials and Methods
2.1. Patients and Ethical Requirements
This research was authorized by the Kaunas Regional Biomedical Research Ethics Committee at the Lithuanian University of Health Sciences (LUHS) (No. BE-2-/61, approval date: 11 October 2017) and adhered to the Declaration of Helsinki’s criteria. The objective and procedure of the study were explained to each participant. Before participating, all 400 study individuals gave their written informed consent. The MS group was formed with 200 individuals. Criteria for inclusion in the MS group:
Exclusion criteria for the MS group:
Patients younger than 18 years.
The patient has received a transfusion of blood or blood components within the last four weeks.
The patient has received treatment with growth factors that counteract blood production in the last four weeks.
The control group included 200 patients. The control group comprised healthy individuals who matched the age and gender distribution of the MS group and who attended LSMUL, KK, the Neurology Clinic, and the Eye Clinic for a preventive examination. Criteria for inclusion in the control group:
Exclusion criteria for the control group:
Patients with subjective neurological complaints.
Patients having spinal anesthesia.
Patients with other neurological diseases without abnormalities in the demyelinating disorder of the brain and/or spinal cord.
After the subject groups were formed, the single-nucleotide polymorphisms (SNPs)
STAT4 rs10181656, rs7574865, rs7601754, and rs10168266 were analyzed. The MS group consisted of 200 people: 88 males (44%) and 112 females (56%). The patients’ median age was 38 years (IQR = 15). The control group consisted of 200 people: 79 males (39.5%) and 121 females (60.5%). The control group’s median age was 33 (IQR = 21). No statistically significant differences between gender and age were found within the control and MS groups.
Table 1 presents the subjects’ demographic information.
2.2. SNP Selection
Encoding a transcription factor belonging to the STAT family, the
STAT4 gene is found on human chromosome 2q32.3 [
7]. The
STAT4 rs7574865, rs10181656, rs7601754, and rs10168266 were chosen for genotyping based on prior research on other autoimmune diseases. The SNP substitutions, SNP regions, chromosomal positions, and primer sequences are listed in
Table 2.
The
STAT4 gene is thought to be linked to several autoimmune disorders; however, distinct susceptibility to the disease may result from different SNPs. The molecular mechanism of the
STAT4 gene’s involvement in the etiology of MS is still unknown because all mutations identified in this study are found in introns and do not directly affect STAT4 transcription or translation [
20].
2.3. DNA Extraction and Genotyping
Each participant’s blood was collected into tubes with ethylenediaminetetraacetic acid (EDTA) Following the manufacturer’s instructions, a genomic DNA extraction kit based on silica-based membrane technology (Thermo Fisher Scientific, Vilnius, Lithuania) was used in the Laboratory of Ophthalmology, Neuroscience Institute, LUHS, to extract DNA. UV spectrophotometry (Agilent Technologies (Andover, MA, USA), Cary 60 UV-Vis) was used to determine the DNA concentrations and purity index in each blood sample as a ratio of absorbance 260/280 nm. Each sample displayed a purity index of 1.8 to 2.0. RT-PCR is a technique used to amplify and quantify DNA in real time, allowing for detecting and quantifying specific DNA sequences in a sample. The RT-PCR method comprised the following steps:
Primer design: Specific primers were designed to amplify the target DNA sequence. Primer sequences [VIC/FAM] are shown in
Table 3.
Probe design: A fluorescent probe was designed to detect the amplified DNA sequence.
PCR reaction setup: The extracted DNA was mixed with the primers, probe, and other reagents needed for PCR amplification.
PCR amplification: The PCR reaction runs through cycles of denaturation, annealing, and extension, resulting in the exponential amplification of the target DNA sequence.
To ensure consistency of the genotyping process and accuracy of the results, a random sample comprising 5% (n = 20) of the total DNA samples was retested.
The data obtained from the RT-PCR were analyzed.
2.4. ELISA
Blood from peripheral vessels was collected to prepare serum. After 30 min of room temperature incubation, the blood samples were centrifuged. Following the pellet’s extraction, the serum was transferred into 2 mL tubes, refrigerated, and kept at −80 °C until analysis. The STAT4 serum levels of the control and MS patient groups were measured using the enzymatic immunoassay (ELISA) for human STAT4 (Human STAT4 ELISA Kit, Abbexa, Cambridge, UK) based on the conventional sandwich ELISA technique. The measurements were taken according to the manufacturer’s specifications. The optical density at 450 nm was measured using a microplate reader (Multiskan FC microplate photometer, Thermo Scientific, Waltham, MA, USA). The STAT4 serum levels were determined using the standard curve. The standard curve displayed a sensitivity of < 0.12 ng/mL and a range of 0.312–20 ng/mL.
2.5. Statistical Analysis
SPSS/W 29.0 (Statistical Package for the Social Sciences for Windows, Inc., Chicago, IL, USA) was the software used for the statistical analysis. The Kolmogorov–Smirnov test was used to determine whether the age was normally distributed. Continuous variables were shown as the median with the interquartile range (IQR) for data that were not normally distributed. To compare the two groups, the Mann–Whitney U test was performed. The chi-square (χ2) test examined the allele distributions, genotype, and gender differences between the MS and control groups. The categorical data were presented as absolute numbers with percentages. The binary logistic regression analysis was used to evaluate the effect of SNPs on MS. An odds ratio (OR) with a 95% confidence interval (CI) were provided for the results. Statistical genetic models were used to present the results of logistic regression. The best genetic model was identified using the Akaike information criterion (AIC). We evaluated four SNPs in the STAT4 gene, and a two-tailed test with a value of less than 0.05 was considered statistically significant. The Bonferroni adjustment was used to modify the significance level for multiple comparisons (p = 0.0125 (0.05/4)). Serum STAT4 levels were compared between groups of MS patients and healthy individuals using the Mann–Whitney U test.
4. Discussion
STAT4 is a transcription factor that plays a crucial role in developing autoimmune diseases [
22]. It encodes an essential transcription factor that carries signals from specific cytokines linked to autoimmune disorders [
8]. Since MS is an autoimmune disease, we looked for associations between
STAT4 SNPs, STAT4 serum levels, and MS. Even though
STAT4 has been linked to a variety of autoimmune disorders—neuromyelitis optica (NMO), systemic lupus erythematosus (SLE), rheumatoid arthritis (RA) systemic sclerosis (SS), MS [
11,
23,
24,
25,
26]—this is, as far as we know, the first study to investigate the relationship between the
STAT4 (rs10181656, rs7574865, rs7601754, and rs10168266), STAT4 serum levels, and the occurrence of MS in the Lithuanian population.
To our knowledge, there is only one study that has investigated an association between an
STAT4 variant and MS. Nageeb et al. hypothesized that
STAT4 rs7582694 gene polymorphism contributes to autoimmune diseases. The results showed that the CC genotype was statistically significantly more frequent in MS patients compared to the control group. Furthermore, the C allele was statistically significantly higher in patients with MS compared to controls [
26].
The demyelinating condition known as NMO is a neurological disorder that matches many clinical characteristics with MS and fulfills all the requirements for an autoimmune origin [
23]. Like MS, NMO causes episodes of optic neuritis and transverse myelitis. In both cases, a person’s immune system sees a healthy part of their body as a threat and attacks it. Shi et al. investigated the association between
STAT4 rs7601754 and NMO. The study showed that the G allele protects against NMO spectrum disorders (
p = 0.006) [
20]. Another autoimmune disease that can damage the CNS is SLE, characterized by various immunological abnormalities [
24]. Several genetic studies have looked into the link between
STAT4 SNPs and SLE risk in different populations, but the results are inconsistent. A meta-analysis showed that
STAT4 rs7601754 and rs7574865 are significantly associated with SLE in European and African populations (
p < 0.001) [
27]. Another meta-analysis conducted by Wang and co-authors confirmed a strong association between the
STAT4 rs7574865 and rs10168266 and susceptibility to SLE (
p < 0.001,
p < 0.001, respectively). This study included 17,389 patients with SLE and 29,273 control subjects [
28]. Ebrahimiyan et al. found that the
STAT4 rs7601754 A allele was significantly associated with a 0.679 lower susceptibility to SLE (OR = 0.679; 95% CI: 0.610–0.747,
p < 0.001) [
22]. Another study showed that the
STAT4 rs7574865 TT genotype and T allele are significant molecular risk markers for predicting susceptibility to SLE and that the GG genotype is a valuable marker against SLE risk [
29]. Analysis of rs10168266 revealed that only the minor allele T was significantly associated with SLE in the Malaysian population (OR = 1.435; 95% CI: 1.143–1.802;
p = 0.014) [
30]. However, another study conducted by Salmaninejad et al. showed that both alleles A and G and the genotypes of rs7601754 did not show statistically significant differences between juvenile SLE patients and the control group [
31].
As the studies show controversial results, we found that the A allele of rs7601754 is significantly associated with higher odds of MS occurrence according to the dominant model (OR = 1.912; 95% CI: 1.237–2.954; p = 0.004) and the additive model (OR = 1.732; 95% CI: 1.193–2.516; p = 0.004) after Bonferroni correction. In addition, the rs7601754 AG genotype is more common in males with MS than in the control group (35.2% vs. 17.7%, p = 0.011). Binary logistic regression analysis in males also revealed that only rs7601754 was associated with 2.5-fold increased odds of MS in males under the overdominant model (OR: 2.525; CI: 1.224–5.211; p = 0.012).
A great model for investigating how the immune system controls neural activity is MS. Accordingly, there is increasing evidence that pro-inflammatory mediators at high levels can seriously disrupt synaptic processes, neuronal excitability in general, and synaptic plasticity [
32]. STAT4 is known for its regulatory role in proinflammatory signaling [
33]. Additionally, STAT4 plays a critical role as a mediator in the development of inflammation in immunological-mediated diseases and protective immune responses. As a result of abrogated Th1 responses, STAT4-deficient mice are resistant to the development of Th1-mediated autoimmune diseases, including EAE, RA, colitis, myocarditis, and diabetes, because they produce a smaller amount of pro-inflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α). [
11]. A meta-analysis showed that the
STAT4 rs7574865 T allele was associated with RA in Europeans (OR = 1.300; 95% CI = 1.195–1.414;
p < 0.001) [
34]. Another study found a statistical association between rs10181656 and RA (
p = 0.007) [
35]. Furthermore, Hanan et al. found that patients carrying the T allele of rs7574865 have a high risk of RA and SLE compared to healthy controls (
p < 0.001) [
36]. It was also noticed that the rs7574865 T allele was statistically significantly associated with susceptibility to SS in the Spanish population (OR = 1.61; 95% CI: 1.29–1.99;
p < 0.001) [
25]. According to a study carried out by Zhang et al., the results showed a statistically significant association between the
STAT4 rs7601754 A allele and the risk of primary biliary cholangitis (OR = 1.35; 95% CI: 1.17–1.55;
p < 0.001) [
37]. Although various sources indicate associations of
STAT4 rs10181656, rs7574865, rs7601754, and rs10168266 with inflammatory and autoimmune diseases, in our study, only rs7601754 was statistically significantly associated with the occurrence of MS.
Inflammation depends on STAT, which controls the behavior of immune cells by facilitating the extracellular signaling of inflammatory mediators. Research shows that cytokines and growth factors can usually bind to their corresponding cell surface receptors to initiate an intracellular tyrosine kinase phosphorylation cascade. This cascade can be modified by kinases such as JAK2, which can alter immune responses, growth, and metabolic processes. Only a few studies have examined the association of STAT4 serum levels with disease risk. A study carried out by Zhang et al. revealed that the placenta of preeclampsia patients had statistically significantly higher STAT4 levels compared to normal late-term pregnant females [
38]. It is also known that the increased systemic inflammatory response triggered by endotoxins is coordinated by excessive cytokine production. A study by Lentsch et al. showed that STAT4 is a vital regulator of the systemic inflammatory response to endotoxins. Mice lacking STAT4 are highly susceptible to lethal endotoxemia. These results indicate that STAT4 protects against endotoxin-induced death [
39]. We found that serum STAT4 levels were statistically significantly lower in MS patients compared to the control group (median (IQR): 0.16 (0.09) vs. 0.26 (0.42),
p = 0.007).
In conclusion, this was the first attempt to evaluate the association of STAT4 SNPs and STAT4 serum levels with MS in the Lithuanian population. Although STAT4 rs10181656, rs7574865, and rs10168266 have been associated with various types of autoimmune and inflammatory diseases, they were not considered as genetic factors contributing to MS in our patient group. Only STAT4 rs7601754 is associated with MS and increases the disease occurence in the Lithuanian population. However, given the small number of patients in the case group of this study, further investigations with a sufficient sample size and in other populations, as well as an evaluation of different potential SNPs, will be helpful interpretations to reach a comprehensive conclusion about the role of STAT4 in MS etiopathogenesis. The lack of association could be due to the small number of patients in the study group. Further studies with larger samples are needed to confirm these results and draw a conclusion.

 ,
, 


{kind=link}
{kind=link}