Linking Sap Flow and Trunk Diameter Measurements to Assess Water Dynamics of Touriga-Nacional Grapevines Trained in Cordon and Guyot Systems

 ,
,  and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Site Description and Plant Material
2.2. Sap Flow Measurements
2.3. Trunk Diameter Measurements
2.4. Other Measurements
2.5. Statistical Analysis
3. Results
3.1. Weather Conditions
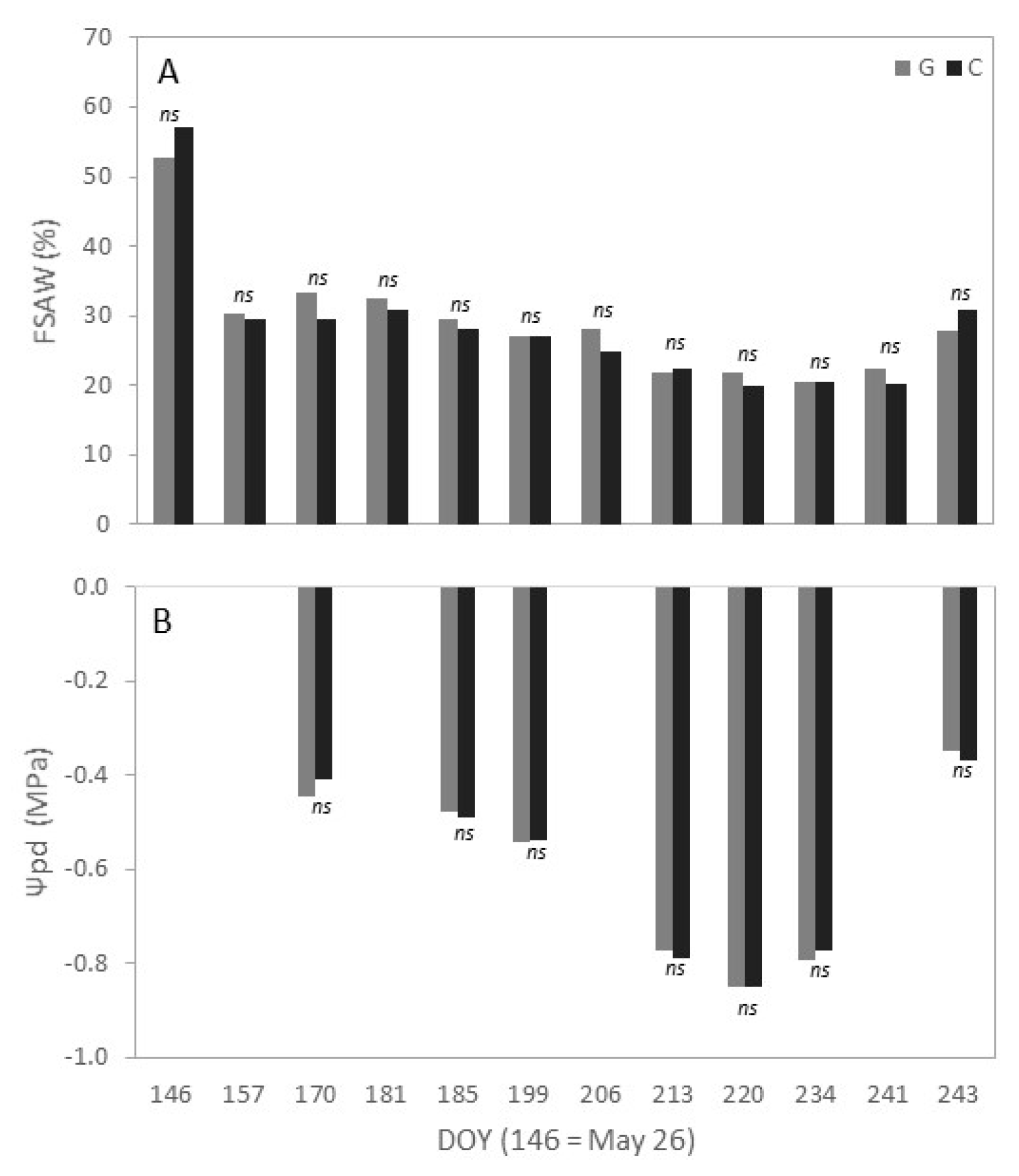
3.2. Fraction of Available Soil Water and Predawn Leaf Water Potential
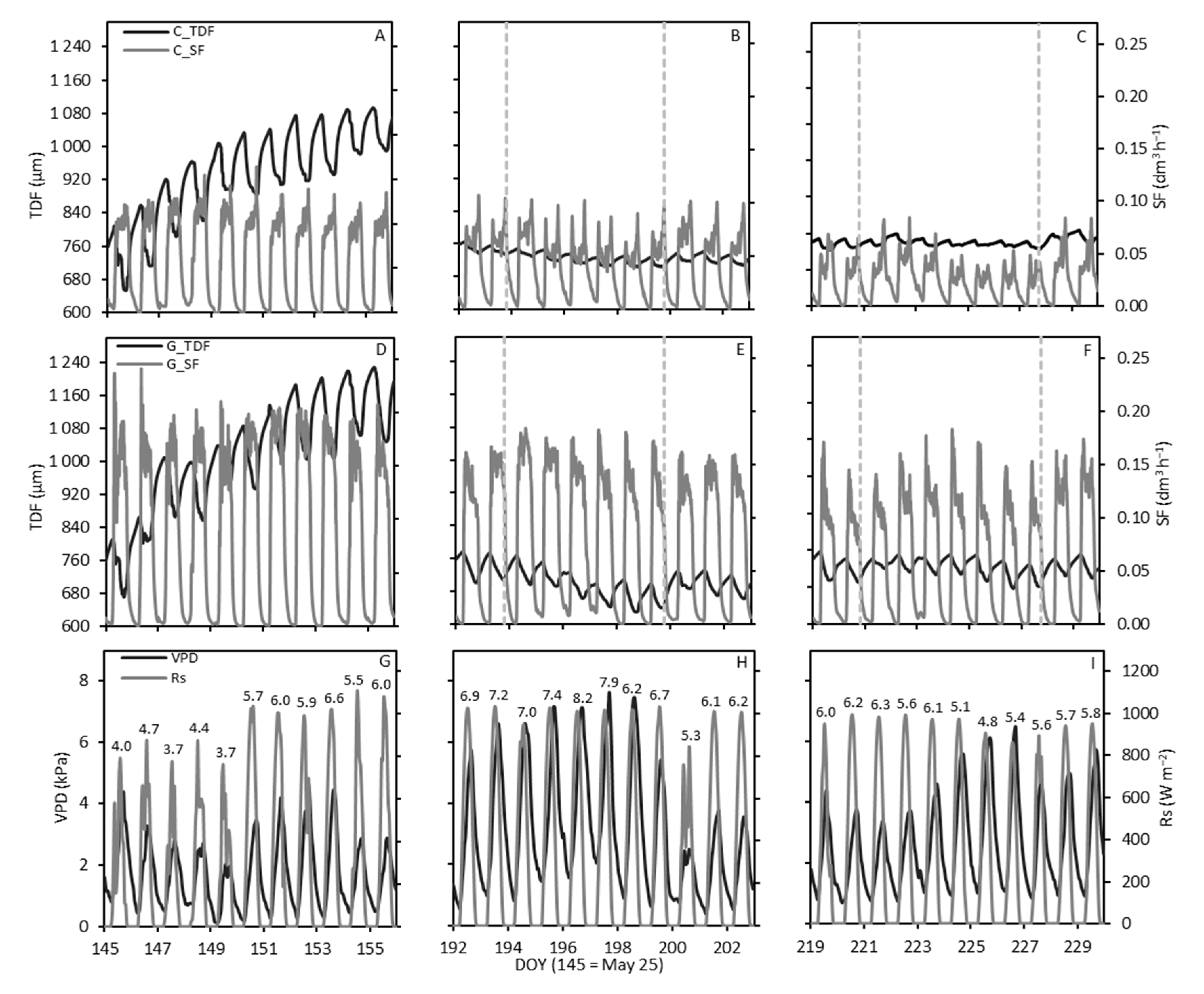
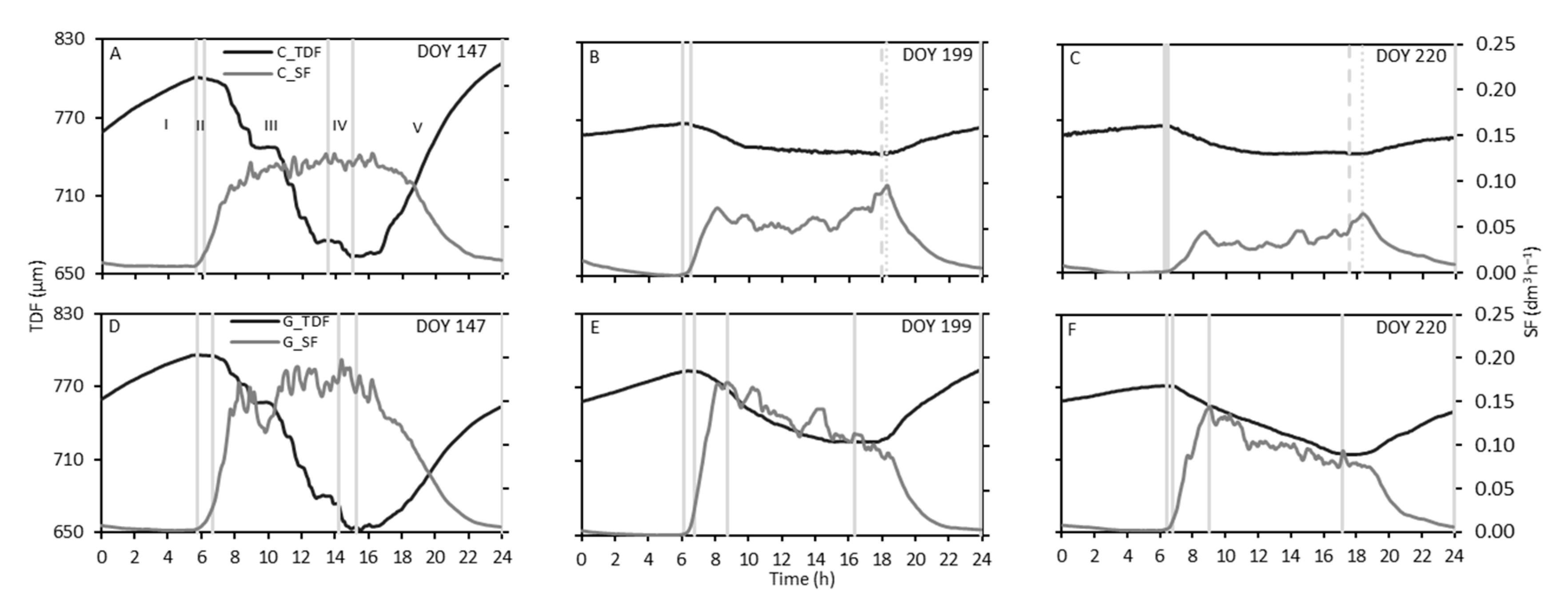
3.3. Sap Flow and Trunk Diameter Fluctuations
3.4. Sap Flow and Leaf Area Index
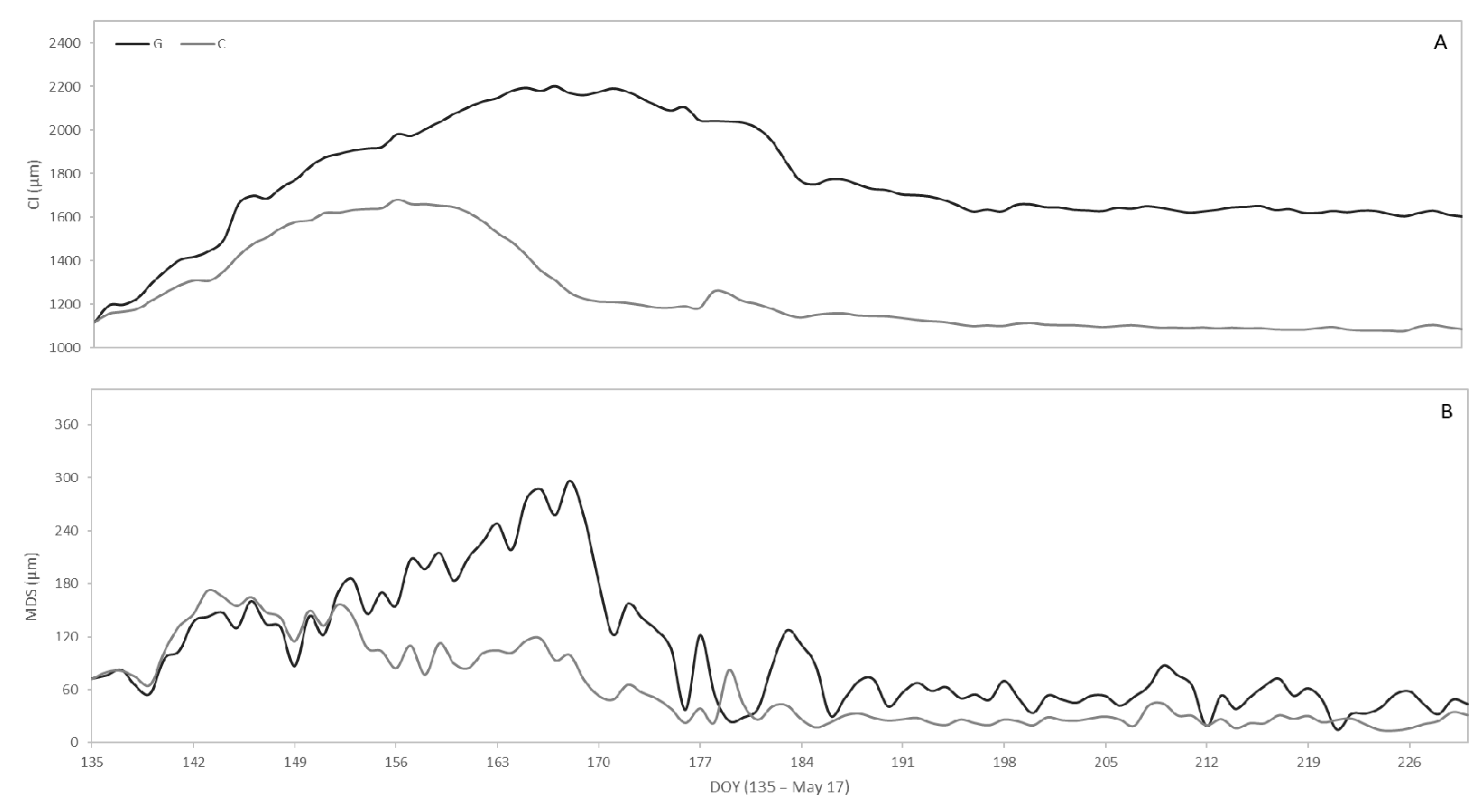
3.5. Seasonal Variation of CI and MDS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Leeuwen, C.; Friant, P.; Choné, X.; Tregoat, O.; Koundouras, S.; Dubourdieu, D. Influence of climate, soil, and cultivar on terroir. Am. J. Enol. Vitic. 2004, 55, 207–217. [Google Scholar]
- Bernardo, S.; Dinis, L.T.; Machado, N.; Moutinho-Pereira, J. Grapevine abiotic stress assessment and search for sustainable adaptation strategies in Mediterranean-like climates. A review. Agron. Sustain. Dev. 2018, 38, 66. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.M.; Zarrouk, O.; Francisco, R.; Costa, J.M.; Santos, T.; Regalado, A.P.; Rodrigues, M.L.; Lopes, C.M. Grapevine under deficit irrigation: Hints from physiological and molecular data. Ann. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinis, L.T.; Correia, C.M.; Ferreira, H.F.; Gonçalves, B.; Gonçalves, I.; Coutinho, J.F.; Ferreira, M.I.; Malheiro, A.C.; Moutinho-Pereira, J. Physiological and biochemical responses of Semillon and Muscat Blanc à Petits Grains winegrapes grown under Mediterranean climate. Sci. Hortic. 2014, 175, 128–138. [Google Scholar] [CrossRef]
- Ferreira, M.I.; Silvestre, J.; Conceição, N.; Malheiro, A.C. Crop and stress coefficients in rainfed and deficit irrigation vineyards using sap flow techniques. Irrig. Sci. 2012, 30, 433–447. [Google Scholar] [CrossRef]
- Fraga, H.; Santos, J.A.; Moutinho-Pereira, J.; Carlos, C.; Silvestre, J.; Eiras-Dias, J.; Mota, T.; Malheiro, A.C. Statistical modelling of grapevine phenology in Portuguese wine regions: Observed trends and climate change projections. J. Agric. Sci. 2016, 154, 795–811. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.A.; Fraga, H.; Malheiro, A.C.; Moutinho-Pereira, J.; Dinis, L.T.; Correia, C.; Moriondo, M.; Leolini, L.; Dibari, C.; Costafreda-Aumedes, S.; et al. A review of the potential climate change impacts and adaptation options for European viticulture. Appl. Sci. 2020, 10, 3092. [Google Scholar] [CrossRef]
- Conejero, W.; Alarcón, J.J.; García-Orellana, Y.; Nicolás, E.; Torrecillas, A. Evaluation of sap flow and trunk diameter sensors for irrigation scheduling in early maturing peach trees. Tree Physiol. 2007, 27, 1753–1759. [Google Scholar] [CrossRef] [Green Version]
- Montoro, A.; Fereres, E.; López-Urrea, R.; Mañas, F.; López-Fuster, P. Sensitivity of trunk diameter fluctuations in Vitis vinifera L. Tempranillo and Cabernet Sauvignon cultivars. Am. J. Enol. Vitic. 2012, 63, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Lovisolo, C.; Lavoie-Lamoureux, A.; Tramontini, S.; Ferrandino, A. Grapevine adaptations to water stress: New perspectives about soil/plant interactions. Theor. Exp. Plant Physiol. 2016, 28, 53–66. [Google Scholar] [CrossRef]
- Malheiro, A.C.; Ferreira, M.I.; Conceição, N.; Green, S. Root and stem water dynamics of rainfed grapevines in the Douro region. Acta Hortic. 2016, 1136, 221–228. [Google Scholar] [CrossRef]
- Cifre, J.; Bota, J.; Escalona, J.M.; Medrano, H.; Flexas, J. Physiological tools for irrigation scheduling in grapevine (Vitis vinifera L.): An open gate to improve water-use efficiency? Agric. Ecosyst. Environ. 2005, 106, 159–170. [Google Scholar] [CrossRef]
- Simonneau, T.; Lebon, E.; Coupel-Ledru, A.; Marguerit, E.; Rossdeutsch, L.; Ollat, N. Adapting plant material to face water stress in vineyards: Which physiological targets for an optimal control of plant water status? OENO One 2017, 51, 167–179. [Google Scholar] [CrossRef]
- Smart, R.E. Principles of Grapevine Canopy Microclimate Manipulation with Implications for Yield and Quality. A Review. Am. J. Enol. Vitic. 1985, 36, 230–239. [Google Scholar]
- Van Leeuwen, C.; Tregoat, O.; Choné, X.; Bois, B.; Pernet, D.; Gaudillére, J.P. Vine water status is a key factor in grape ripening and vintage quality for red bordeaux wine. How can it be assessed for vineyard management purposes? J. Int. Sci. Vigne Vin 2009, 43, 121–134. [Google Scholar] [CrossRef]
- Reynolds, A.G.; Vanden Heuvel, J.E. Influence of grapevine training systems on vine growth and fruit composition: A review. Am. J. Enol. Vitic. 2009, 60, 251–268. [Google Scholar]
- Fragasso, M.; Antonacci, D.; Pati, S.; Tufariello, M.; Baiano, A.; Forleo, L.R.; Caputo, A.R.; La Notte, E. Influence of training system on volatile and sensory profiles of primitivo grapes and wines. Am. J. Enol. Vitic. 2012, 63, 477–486. [Google Scholar] [CrossRef]
- Ferree, D.; Steiner, T.; Gallander, J.; Scurlock, D.; Johns, G.; Riesen, R. Performance of “Seyval Blanc” grape in four training systems over five years. HortScience 2002, 37, 1023–1027. [Google Scholar] [CrossRef] [Green Version]
- Cavallo, P.; Poni, S.; Rotundo, A. Ecophysiology and vine performance of cv. “Aglianico” under various training systems. Sci. Hortic. 2001, 87, 21–32. [Google Scholar] [CrossRef]
- IVV. Vinhos e Aguardentes de Portugal, Anuário 2018; Ministério da Agricultura, do Desenvolvimento Rural e das Pescas, Instituto da Vinha e do Vinho: Lisboa, Portugal, 2018. [Google Scholar]
- Magalhães, N. Tratado de Viticultura: A Videira, a Vinha e o “Terroir”, 1st ed.; Esfera Poética: Lisboa, Portugal, 2015; 608p, ISBN 978-972-8987-15-2. [Google Scholar]
- Fernández, J.E. Plant-based methods for irrigation scheduling of woody crops. Horticulturae 2017, 3, 35. [Google Scholar] [CrossRef] [Green Version]
- Meng, Z.; Duan, A.; Chen, D.; Dassanayake, K.B.; Wang, X.; Liu, Z.; Liu, H.; Gao, S. Suitable indicators using stem diameter variation-derived indices to monitor the water status of greenhouse tomato plants. PLoS ONE 2017, 12, e0171423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escalona, J.; Flexas, J.; Medrano, H. Drought effects on water flow, photosynthesis and growth of potted grapevines. Vitis 2002, 41, 57–62. [Google Scholar]
- Daudet, F.A.; Améglio, T.; Cochard, H.; Archilla, O.; Lacointe, A. Experimental analysis of the role of water and carbon in tree stem diameter variations. J. Exp. Bot. 2005, 56, 135–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velez, J.E.; Intrigliolo, D.S.; Castel, J.R. Scheduling deficit irrigation of citrus trees with maximum daily trunk shrinkage. Agric. Water Manag. 2007, 90, 197–204. [Google Scholar] [CrossRef]
- Moriana, A.; Girón, I.F.; Martín-Palomo, M.J.; Conejero, W.; Ortuño, M.F.; Torrecillas, A.; Moreno, F. New approach for olive trees irrigation scheduling using trunk diameter sensors. Agric. Water Manag. 2010, 97, 1822–1828. [Google Scholar] [CrossRef]
- de la Rosa, J.M.; Conesa, M.R.; Domingo, R.; Torres, R.; Pérez-Pastor, A. Feasibility of using trunk diameter fluctuation and stem water potential reference lines for irrigation scheduling of early nectarine trees. Agric. Water Manag. 2013, 126, 133–141. [Google Scholar] [CrossRef]
- Granier, A. Une nouvelle méthode pour la mesure du flux de sève brute dans le tronc des arbres. Ann. Sci. For. 1985, 42, 193–200. [Google Scholar] [CrossRef]
- de Swaef, T.; Steppe, K. Linking stem diameter variations to sap flow, turgor and water potential in tomato. Funct. Plant Biol. 2010, 37, 429–438. [Google Scholar] [CrossRef]
- Ortuño, M.F.; García-Orellana, Y.; Conejero, W.; Ruiz-Sánchez, M.C.; Mounzer, O.; Alarcón, J.J.; Torrecillas, A. Relationships between climatic variables and sap flow, stem water potential and maximum daily trunk shrinkage in lemon trees. Plant Soil 2006, 279, 229–242. [Google Scholar] [CrossRef]
- Ortuño, M.F.; García-Orellana, Y.; Conejero, W.; Ruiz-Sánchez, M.C.; Alarcón, J.J.; Torrecillas, A. Stem and leaf water potentials, gas exchange, sap flow, and trunk diameter fluctuations for detecting water stress in lemon trees. Trees Struct. Funct. 2006, 20, 1–8. [Google Scholar] [CrossRef]
- Coehn, M.; Goldhamer, D.A.; Fereres, E.; Girona, J.; Mata, M. Assessment of peach tree responses to irrigation water ficits by continuous monitoring of trunk diameter changes. J. Hortic. Sci. Biotechnol. 2001, 76, 55–60. [Google Scholar] [CrossRef]
- Conejero, W.; Alarcón, J.J.; García-Orellana, Y.; Abrisqueta, J.M.; Torrecillas, A. Daily sap flow and maximum daily trunk shrinkage measurements for diagnosing water stress in early maturing peach trees during the post-harvest period. Tree Physiol. 2007, 27, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, J.E.; Moreno, F.; Martín-Palomo, M.J.; Cuevas, M.V.; Torres-Ruiz, J.M.; Moriana, A. Combining sap flow and trunk diameter measurements to assess water needs in mature olive orchards. Environ. Exp. Bot. 2011, 72, 330–338. [Google Scholar] [CrossRef]
- Cuevas, M.V.; Torres-Ruiz, J.M.; Álvarez, R.; Jiménez, M.D.; Cuerva, J.; Fernández, J.E. Assessment of trunk diameter variation derived indices as water stress indicators in mature olive trees. Agric. Water Manag. 2010, 97, 1293–1302. [Google Scholar] [CrossRef]
- Intrigliolo, D.S.; Castel, J.R. Evaluation of grapevine water status from trunk diameter variations. Irrig. Sci. 2007, 26, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Baert, A.; Villez, K.; Steppe, K. Automatic drought stress detection in grapevines without using conventional threshold values. Plant Soil 2013, 369, 439–452. [Google Scholar] [CrossRef]
- Oliveira, M.T. Dependence of Sap Flow and Stem Diameter Fluctuation of Grapevines on Reference Evapotranspiration: An Event Coincidence Analysis. Adv. Plants Agric. Res. 2017, 7, 433–439. [Google Scholar] [CrossRef] [Green Version]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classification update. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Fonseca, A.R.; Santos, J.A. High-resolution temperature datasets in Portugal from a geostatistical approach: Variability and extremes. J. Appl. Meteorol. Climatol. 2018, 57, 627–644. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration: Guidelines for Computing Crop Water Requirements; FAO Irrigation and Drainage Paper 56; Food and Agriculture Organization of the United Nations: Rome, Italy, 1998; ISBN 9251042195. [Google Scholar]
- Agroconsultores and Coba. Carta de Solos, Carta de Uso Actual da Terra e Carta de Aptidão da Terra, do Nordeste de Portugal; Universidade de Trás-os-Montes e Alto Douro (UTAD): Vila Real, Portugal, 1991. [Google Scholar]
- Granier, A.; Gross, P. Mesure du flux de sève brute dans le tronc du Douglas par une nouvelle méthode thermique. Ann. Sci. For. 1987, 44, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Katerji, N.; Tardieu, F.; Bethenod, O.; Quetin, P. Behavior of Maize Stem Diameter during Drying Cycles: Comparison of Two Methods for Detecting Water Stress. Crop Sci. 1994, 34, 165–169. [Google Scholar] [CrossRef]
- Zweifel, R.; Item, H.; Häsler, R. Stem radius changes and their relation to stored water in stems of young Norway spruce trees. Trees Struct. Funct. 2000, 15, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Fernández, J.E.; Cuevas, M.V. Irrigation scheduling from stem diameter variations: A review. Agric. For. Meteorol. 2010, 150, 135–151. [Google Scholar] [CrossRef]
- Scholander, P.F.; Hammel, H.J.; Bradstreet, A.; Hemmingsen, E.A. Sap Pressure in Vascular Plants. Science 1965, 148, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.C. Measurement of plant water status by the pressure chamber technique. Irrig. Sci. 1988, 9, 289–308. [Google Scholar] [CrossRef]
- López-Lozano, R.; Casterad, M.A. Comparison of different protocols for indirect measurement of leaf area index with ceptometers in vertically trained vineyards. Aust. J. Grape Wine Res. 2013, 19, 116–122. [Google Scholar] [CrossRef] [Green Version]
- Herzog, K.M.; Häsler, R.; Thum, R. Diurnal changes in the radius of a subalpine Norway spruce stem: Their relation to the sap flow and their use to estimate transpiration. Trees 1995, 10, 94–101. [Google Scholar] [CrossRef]
- Malheiro, A.C. Microclimate, Yield and Water-Use of Vineyards in the Douro Region, Portugal. Ph.D. Thesis, Cranfield University, Bedfordshire, UK, 2005. [Google Scholar]
- Winkel, T.; Rambal, S. Influence of Water Stress on Grapevines Growing in the Field: From Leaf to Whole-Plant Response. Aust. J. Plant Physiol. 1993, 20, 143–157. [Google Scholar] [CrossRef]
- Ojeda, H.; Saurin, N. L’irrigation de précision de la vigne: Méthodes, outils et stratégies pour maximiser la qualité et les rendements de la vendange en économisant de l’eau. Innov. Agron. 2014, 38, 97–108. [Google Scholar]
- Santesteban, L.G.; Palacios, I.; Miranda, C.; Iriarte, J.C.; Royo, J.B.; Gonzalo, R. Terahertz time domain spectroscopy allows contactless monitoring of grapevine water status. Front. Plant Sci. 2015, 6, 404. [Google Scholar] [CrossRef] [Green Version]
- Kramer, P.J. Water Relations Plants; Academic Press: New York, NY, USA, 1983. [Google Scholar]
- Pereira, A.R.; Green, S.; Villa Nova, N.A. Penman-Monteith reference evapotranspiration adapted to estimate irrigated tree transpiration. Agric. Water Manag. 2006, 83, 153–161. [Google Scholar] [CrossRef]
- Pearce, I.; Coombe, B.G. Grapevine phenology. In Viticulture; Dry, P.R., Coombe, B.G., Eds.; Winetitles: Broadview, Australia, 2005. [Google Scholar]
- Ambrose, A.R.; Sillett, S.C.; Koch, G.W.; Van Pelt, R.; Antoine, M.E.; Dawson, T.E. Effects of height on treetop transpiration and stomatal conductance in coast redwood (Sequoia sempervirens). Tree Physiol. 2010, 30, 1260–1272. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.G.; Phillips, N.; Bond, B.J. The hydraulic limitation hypothesis revisited. Plant Cell Environ. 2006, 29, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Favero, A.C.; Amorim, D.A.D.; Mota, R.V.D.; Souza, C.R.D.; Regina, M.D.A. Physiological responses and production of “Syrah” vines as a function of training systems. Sci. Agric. 2010, 67, 267–273. [Google Scholar] [CrossRef] [Green Version]
- Schubert, A.; Lovisolo, C.; Peterlunger, E. Shoot orientation affects vessel size, shoot hydraulic conductivity and shoot growth rate in Vitis vinifera L. Plant Cell Environ. 1999, 22, 197–204. [Google Scholar] [CrossRef]
- Zweifel, R. Radial stem variations—A source of tree physiological information not fully exploited yet. Plant Cell Environ. 2016, 39, 231–232. [Google Scholar] [CrossRef]
- Zweifel, R.; Haeni, M.; Buchmann, N.; Eugster, W. Are trees able to grow in periods of stem shrinkage? New Phytol. 2016, 211, 839–849. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Kang, S.; Li, F.; Li, S.; Du, T.; Tong, L. Relationship between environmental factor and maximum daily stem shrinkage in apple tree in arid region of northwest China. Sci. Hortic. 2011, 130, 118–125. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DOY | 185 | 199 | 213 | 220 | 234 | |
|---|---|---|---|---|---|---|
| LAI | Cordon | 0.94 | 1.17 | 0.84 | 0.81 | 0.79 |
| Guyot | 0.91 | 1.12 | 0.79 | 0.76 | 0.74 | |
| p-value | 0.378 | 0.476 | 0.421 | 0.261 | 0.193 | |
| SF/LAI | Cordon | 1.27 | 1.05 | 1.25 | 1.08 | 0.96 |
| Guyot | 1.73 | 1.47 | 1.67 | 1.51 | 1.35 | |
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.005 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malheiro, A.C.; Pires, M.; Conceição, N.; Claro, A.M.; Dinis, L.-T.; Moutinho-Pereira, J. Linking Sap Flow and Trunk Diameter Measurements to Assess Water Dynamics of Touriga-Nacional Grapevines Trained in Cordon and Guyot Systems. Agriculture 2020, 10, 315. https://doi.org/10.3390/agriculture10080315
Malheiro AC, Pires M, Conceição N, Claro AM, Dinis L-T, Moutinho-Pereira J. Linking Sap Flow and Trunk Diameter Measurements to Assess Water Dynamics of Touriga-Nacional Grapevines Trained in Cordon and Guyot Systems. Agriculture. 2020; 10(8):315. https://doi.org/10.3390/agriculture10080315
Chicago/Turabian StyleMalheiro, Aureliano C., Mafalda Pires, Nuno Conceição, Ana M. Claro, Lia-Tânia Dinis, and José Moutinho-Pereira. 2020. "Linking Sap Flow and Trunk Diameter Measurements to Assess Water Dynamics of Touriga-Nacional Grapevines Trained in Cordon and Guyot Systems" Agriculture 10, no. 8: 315. https://doi.org/10.3390/agriculture10080315