
Nutritional, Antinutritional Compounds and Nutraceutical Significance of Native Bean Species (Phaseolus spp.) of Mexican Cultivars

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
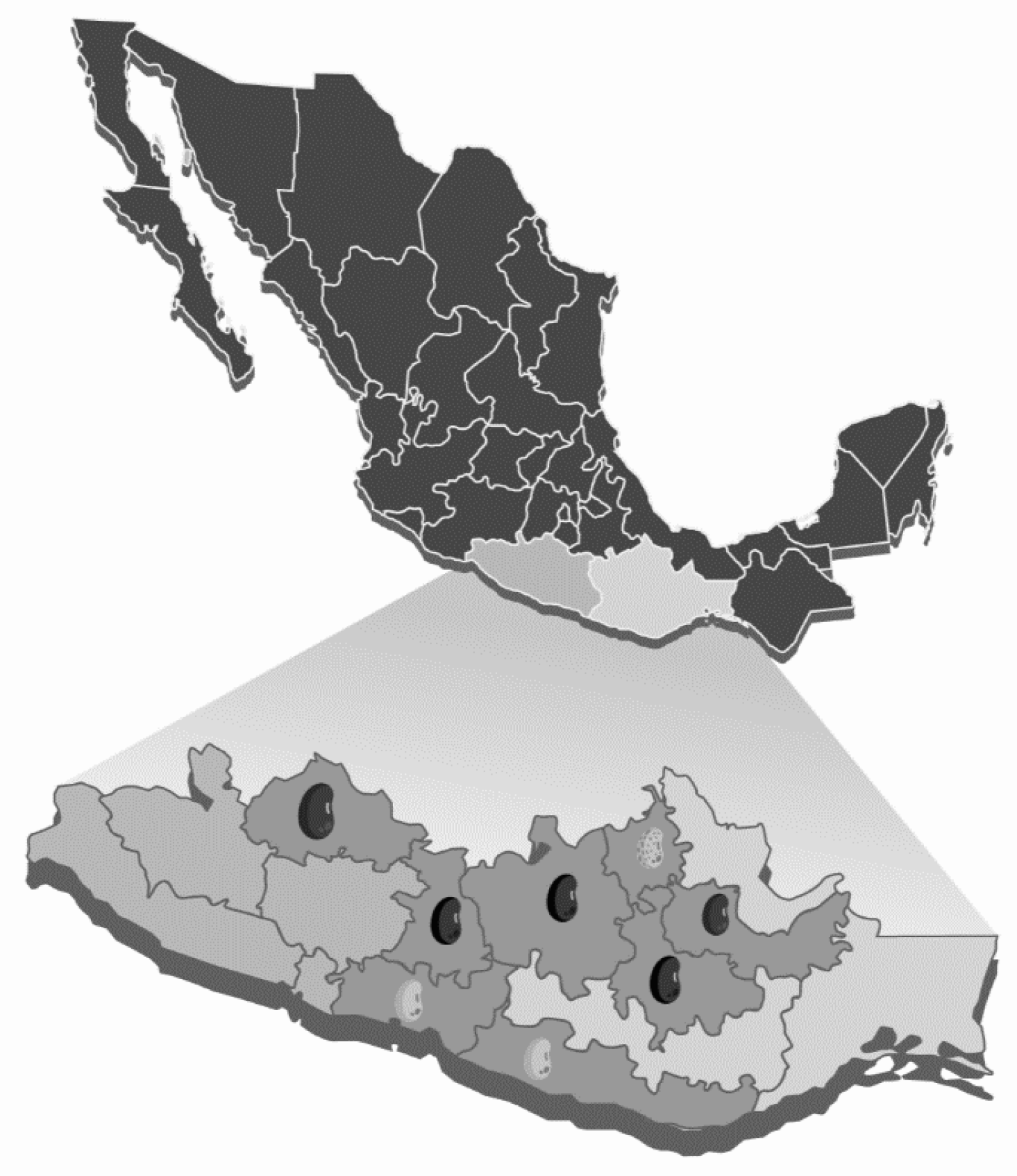
2.1. Plant Material
2.2. Physicochemical Analysis
2.3. Phenolic Compaounds
2.4. Antioxidant Activity
2.5. Antinutritonal Compounds
2.6. Statistical Analysis
3. Results
3.1. Physical Analysis
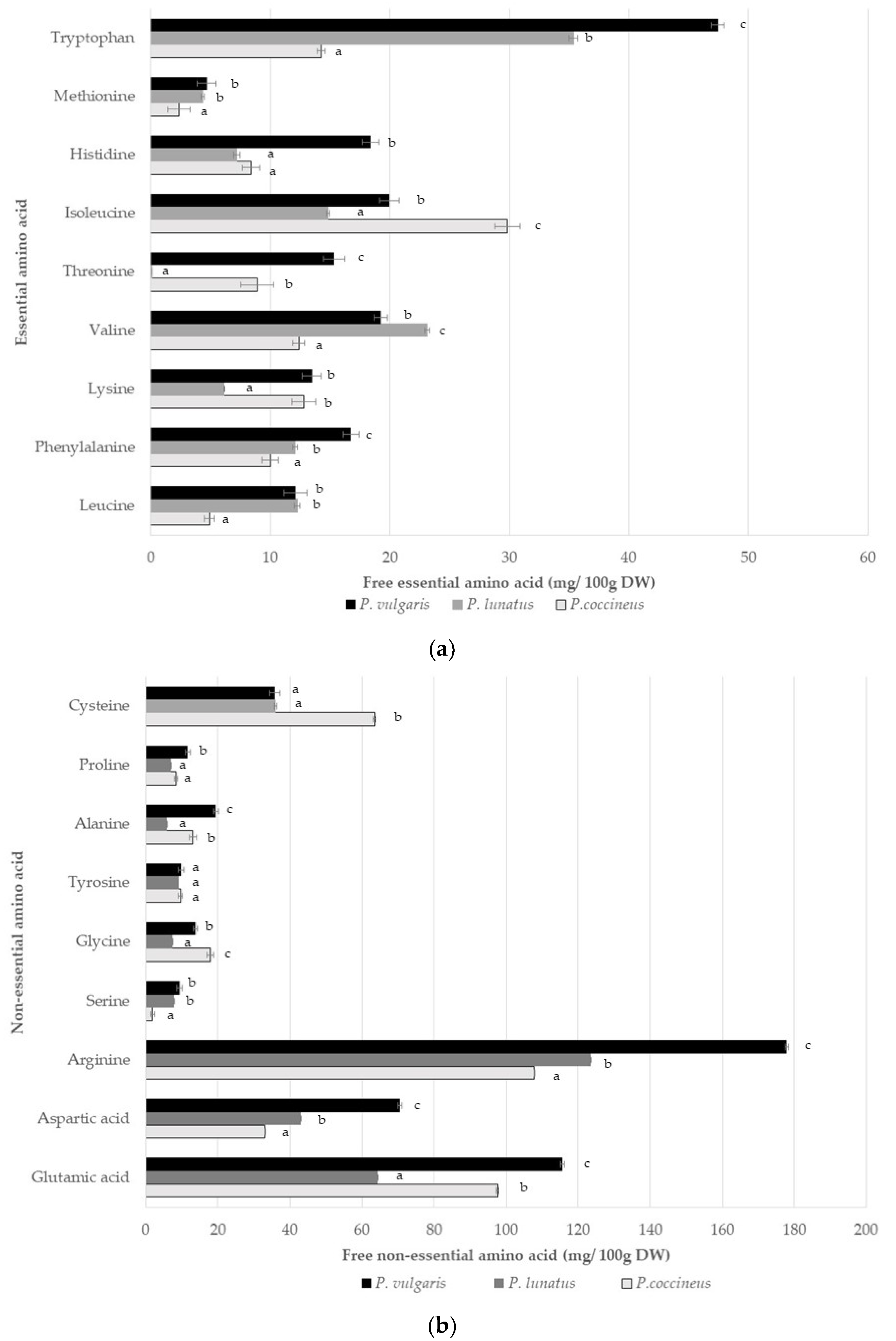
3.2. Nutritional Content
3.3. Phenolic Compounds, Antioxidant Activity, and Anti-Nutritional Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Akillioglu, H.G.; Karakaya, S. Changes in total phenol, total flavonoids, and antioxidant activities of common bean and pinto beans after soaking, cooking, and in vitro digestion process. Food Sci. Biotechnol. 2010, 19, 633–639. [Google Scholar] [CrossRef]
- Ignat, I.; Volf, I.; Popa, V.I. A critical review of methods for the characterization of polyphenolic compounds in fruits and vegetables. Food Chem. 2011, 126, 1821–1835. [Google Scholar] [CrossRef] [PubMed]
- Carbas, B.; Machado, N.; Oppolzer, D.; Ferreira, L.; Brites, C.; Rosa, A.S.E.; Barros, A. Comparison of near infrared (NIR) and mid-infrared (MIR) spectroscopy for the determination of nutritional and antinutritional parameters in common beans. Food Chem. 2020, 306, 125509. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Maldonado, S.H.; Acosta-Gallegos, J.A.; Álvarez-Muños, M.A.; García-Delgado, S.; Loarca-Piña, G. Food quality and nutraceutical potential of common bean (Phaseolus vulgaris L.). Agric. Técnica En México 2002, 28, 159–173. [Google Scholar]
- Hayat, I.; Ahmad, A.; Masud, T.; Ahmed, A.; Bashir, S. Nutritional and health perspectives of beans (Phaseolus vulgaris L.): An overview. Crit. Rev. Food Sci. Nutr. 2014, 54, 580–592. [Google Scholar] [CrossRef]
- Sinkovic, L.; Pipan, B.; Sinkovic, E.; Meglic, V. Morphological seed characterization of common (Phaseolus vulgaris L.) and runner (Phaseolus coccineus L.) bean germplasm: A Slovenian gene bank example. Biomed. Res. Int. 2019, 6376948. [Google Scholar] [CrossRef] [Green Version]
- Capistrán-Carabarin, A.; Aquino-Bolaños, E.N.; Garcia-Díaz, Y.D.; Chávez-Sevia, J.L.; Vera-Guzmán, A.M.; Carrillo-Rodríguez, J.C. Complementary in phenolic compounds and the antioxidant activities of Phaseolus coccineus L. and P. vulgaris L. landraces. Foods 2019, 8, 295. [Google Scholar] [CrossRef] [Green Version]
- Alcázar-Valle, M.; Lugo-Cervantes, E.; Mojica, L.; Morales-Hernández, N.; Reyes-Ramírez, H.; Enríquez-Vara, J.N.; García-Morales, S. Bioactive compounds, antioxidant activity, and antinutritional content of legumes: A comparison between four Phaseolus species. Molecules 2020, 25, 3528. [Google Scholar] [CrossRef]
- Alvarado-López, A.N.; Gómez-Oliván, L.M.; Heredia, J.B.; Baeza-Jiménez, R.; Garcia-Galindo, H.S.; Lopez-Martinez, L.X. Nutritional and bioactive characteristics of Ayocote bean (Phaseolus coccineus L.): An underutilized legume harvested in Mexico. CYTA-J. Food 2019, 17, 199–206. [Google Scholar] [CrossRef] [Green Version]
- Farinde, E.O.; Olanipeku, O.T.; Olasupo, R.B. Nutritional composition and antinutrients content of raw and processed lima bean (Phaseolus lunatus). Annals Food Sci. Technol. 2018, 19, 250–264. [Google Scholar]
- Flores-Sosa, A.R.; Aquino-Bolaños, E.N.; Cardador-Martínez, A.; Chávez-Servia, J.S.; Vera-Guzmán, A.M.; Carrillo-Rodríguez, J.C.; Jiménez, J.E. Variation in protein and amino acids content among landraces of common bean (Phaseolus vulgaris L.). Emir. J. Food Agric. 2020, 32, 750–760. [Google Scholar] [CrossRef]
- Kahsay, B.N.; Ziegler, J.; Imming, P.; Gebre-Mariam, T.; Neubert, R.H.H.; Moeller, L. Free amino acid contents of selected Ethiopian plant and fungi species: A search for alternative natural free amino acid sources for cosmeceutical applications. Amino acids 2021, 53, 1105–1122. [Google Scholar] [CrossRef] [PubMed]
- Barreto-Peixoto, J.A.; Álvarez-Rivera, G.; Alves, R.C.; Costa, A.S.G.; Machado, S.; Cifuentes, A.; Ibáñez, E.; Oliveira, M.B.P.P. Comprehensive phenolic and acid free amino acid analysis of rosemary infusions: Influence on the antioxidant potential. Antioxidants 2021, 10, 500. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Gepts, P.; Debouck, D.G. Races of common bean (Phaseolus vulgaris, Fabaceae). Econ. Bot. 1991, 45, 379–396. [Google Scholar] [CrossRef]
- McGuire, R.G. Reporting of objective color measurements. HortSci 1992, 27, 1254–1255. [Google Scholar] [CrossRef] [Green Version]
- AOAC, Association of Analytical Communities. Official Methods of Analysis of AOAC International, 16th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 1995. [Google Scholar]
- AOAC, Association of Analytical Communities. Official Methods Amino Acids Analysis, 17th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2006. [Google Scholar]
- Xu, B.J.; Chang, S.K.C. A comparative study on phenolic profiles and antioxidant activities of legumes as affected by extraction solvents. J. Food Sci. 2007, 72, 159–166. [Google Scholar] [CrossRef]
- Agostini-Costa, T.S.; Teodoro, A.F.P.; Alves, R.B.N.; Braga, L.R.; Ribeiro, I.F.; Silva, J.P.; Quintana, L.P.; Burle, M.L. Total phenolics, flavonoids, and antioxidant activity of lima beans conserved in a Brazilian genebank. Cienc. Rural. St. Maria 2015, 45, 335–341. [Google Scholar] [CrossRef] [Green Version]
- Del-Toro-Sánchez, C.L.; Gutiérrez-Lomeli, M.; Lugo-Cervantes, E.; Zurita, F.; Robles-García, M.A.; Ruíz-Cruz, S.; Aguilar, J.A.; Morales del Río, J.A.; Guerrero-Medina, P.J. Storage effect on phenols and the antioxidant activity of extracts from Anemopsis californica and inhibition of eleastase enzyme. J. Chem 2015, 602136. [Google Scholar] [CrossRef] [Green Version]
- Broadhurst, R.B.; Jones, W.T. Analysis of condensed tannins using acidified vanillin. J. Sci. Food Agric. 1978, 29, 788–794. [Google Scholar] [CrossRef]
- Sun, B.; Ricardo-da-Silva, J.M.; Spranger, I. Critical factors of vanillin assay for catechins and proanthocyanidins. J. Agric. Food Chem. 1998, 46, 4267–4274. [Google Scholar] [CrossRef]
- Mojica, L.; Meyer, A.; Berhow, M.A.; Gónzalez de Mejia, E. Bean cultivars (Phaseolus vulgaris L.) have similar high antioxidant capacity, in vitro inhibition of α-amylase and α-glucosidase while diverse phenolic composition and concentration. Food Res. Int. 2015, 69, 38–48. [Google Scholar] [CrossRef]
- Tovar-Pérez EGGuerrero-Becerra, L.; Lugo-Cervantes, E. Antioxidant activity of hydrolysates and peptide fractions of glutelin from cocoa (Theobroma cacao L.) seed. CyTA—J. Food 2017, 15, 489–496. [Google Scholar] [CrossRef] [Green Version]
- De-Mejia, E.G.; Hankins, C.N.; Paredes-Lopez, O.; Shannon, L.M. The lectins and lectin-like proteins of tepary beans (Phaseolus acutifolius) and tepary-common bean (Phaseolus vulgaris) hybrids. J. Food Biochem. 1990, 14, 117–126. [Google Scholar] [CrossRef]
- Kakade, M.L.; Rackis, J.J.; McGhee, J.E.; Puski, G. Determination of trypsin inhibitor activity of soy products: A collaborative analysis of an improved procedure. Cereal Chem. 1974, 51, 376–382. [Google Scholar]
- Espinosa-Pérez, E.N.; Ramírez-Vallejo, P.; Crosby-Galván, M.M.; Estrada-Gómez, J.A.; Lucas-Florentino, B.; Chávez-Servia, J.L. Classification of common dry bean landraces from the south-center of Mexico by seed morphology. Rev. Fitotec. Mex. 2015, 29, 29–38. [Google Scholar]
- Aquino-Bolanos, E.N.; Garcia-Diaz, Y.D.; Chavez-Servia, J.L.; Carrillo-Rodríguez, J.C.; Vera-Guzmán, A.M.; Herendia-García, E. Anthocyanin, polyphenol, and flavonoid contents and antioxidant activity in Mexican common bean (Phaseolus vulgaris L.) landraces. Emir. J. Food Agric. 2016, 28, 581–588. [Google Scholar] [CrossRef] [Green Version]
- López-Alcocer, J.J.; Lépiz-Ildefonso RGónzalez-Eguiarte, D.R.; Rodríguez-Macias, R.; López-Alcocer, E. Morphological variability of wild Phaseolus lunatus L. from the western region of Mexico. Rev. Fitotec. Mex. 2016, 39, 49–58. [Google Scholar]
- Solano-Cervantes, F.; Diaz-Ruiz, R.; Jacinto-Hernández, C.; Aguirre-Álvarez, L.; Huerta de la Peña, A. Agricultural practices, morphologic, proteinic and culinary description of the grain of bean cultivars showed in the regions of Tlatzala Guerrero. RaXimahai 2009, 5, 187–199. Available online: http://www.redaluc.org/articulo.oa?id=46111507005 (accessed on 11 May 2021).
- Castillo-Mendoza, M.; Ramírez-Vallejo, P.; Catillo-González, F.; Miranda-Colin, S. Morphological diversity in landraces of common and runner beans from the eastern Mexico state. Rev. Fitote. Mex. 2006, 29, 111–119. [Google Scholar]
- Pastor-Cavada, E.; Juan, R.; Pastor, J.E.; Alaiz, M.; Vioque, J. Protein and amino acid composition of select wild legume species of tribe Fabaceae. Food Chem. 2014, 163, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Baptista, A.; Pinho, O.; Pinto, E.; Casal, S.; Mota, C.; Ferreira, I.M.P.L.V.O. Characterization of protein and fat composition of seeds from common beans (Phaseolus vulgaris L.), cowpea (Vigna unguiculata L. Walp), and Babara froundnuts (Vigna subterranean L. Verdc) from Mozambique. Food Meas. 2017, 11, 442–450. [Google Scholar] [CrossRef]
- Grela, E.R.; Kiczorowsha, B.; Samolinska, W.; Metras, J.; Kiczorowski, P.; Rybinski, W.; Hanczakowska, E. Chemical composition of leguminous seeds: Part I-content of basic nutrients, amino acid, phytochemical compounds, and antioxidant activity. Eur. Food Res. Technol. 2017, 243, 1385–1395. [Google Scholar] [CrossRef]
- Kan, L.; Nie, S.; Hu, J.; Wang, S.; Cui, S.W.; Li, Y.; Xu, S.; Wu, Y.; Wang, J.; Bai, Z.; et al. Nutrients, phytochemicals and antioxidant activities of 26 kidney bean cultivars. Food Chem. Toxicol. 2017, 108, 467–477. [Google Scholar] [CrossRef]
- Mecha, E.; Natalello, S.; Carbas, B.; da Silva, A.B.; Leitão, S.T.; Brites, C.; Veloso, M.M.; Rubiales, D.; Costa, J.; Cabral, M.; et al. Bronze, M.R. Disclosing the Nutritional Quality Diversity of Portuguese Common Beans—The Missing Link for Their Effective Use in Protein Quality Breeding Programs. Agronomy 2021, 11, 221. [Google Scholar] [CrossRef]
- Ito, H.; Ueno, H.; Kikuzaki, H. Free Amino Acid Compositions for fruits. J. Nutr. Diet. Pract. 2017, 1, 001–005. [Google Scholar]
- Brestensky, M.; Nitrayova, S.; Patras, P.; Heger, J.; Nitray, J. Branched-chain amino acids and their importance in nutrition. J. Microbiol. Biotechnol. Food Sci. 2015, 5, 197–202. [Google Scholar] [CrossRef]
- Friedman, M. Analysis, nutrition, and health benefits of tryptophan. Int. J. Try Res. 2018, 11, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Sharma, A.; Kaur, R.; Thukral, A.K.; Bhardwaj, R.; Ahmad, P. Differential distribution of amino acids in plants. Amino Acids. 2017, 49, 821–869. [Google Scholar] [CrossRef] [PubMed]
- Diaz, A.M.; Caldas, G.V.; Blair, M.V. Concentration of condensed tannins and anthocyanins in common bean seed coats. Food Res. Int. 2010, 43, 595–601. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | *L | H | c | Length (mm) | Width (mm) | Thickness (mm) | Weight (g) | Color |
|---|---|---|---|---|---|---|---|---|
| GR-01 P.l. | 24.24 ± 0.14 i–k | 1.27 ± 0.02 d–i | 1.05 ± 0.03 a–c | 15.22 ± 1.25 t | 11.56 ± 0.89 v | 4.38 ± 0.31 d–g | 57.31 | Black–purple |
| GR-02 P.v. | 39.46 ± 0.78 p | 0.89 ± 0.00 b–i | 22.38 ± 0.73 q | 8.58 ± 065 bc | 6.44 ± 0.46 j–m | 5.05 ± 0.38 m–o | 17.13 | Brown |
| GR-03 P.v. | 68.64 ± 0.22 w | 1.44 ± 0.00 hi | 14.00 ± 0.10 h | 8.50 ± 0.75 b | 5.86 ± 0.38 m–o | 5.10 ± 0.42 no | 20.47 | White |
| GR-04 P.v. | 21.31 ± 0.13 ef | 1.45 ± 0.02 hi | 1.39 ± 0.01 b–d | 9.91 ± 0.77 il | 5.94 ± 0.54 d–f | 4.06 ± 0.40 bc | 19.38 | Black–purple |
| GR-05 P.v. | 22.03 ± 0.21fg | 1.50 ± 0.01 i | 2.23 ± 0.17 de | 10.63 ± 0.69 no | 6.46 ± 0.32 k–m | 4.32 ± 0.29 c–f | 22.74 | Black–purple |
| GR-06 P.v. | 68.64 ± 0.20 w | 1.33 ± 0.00 e–i | 19.60 ± 0.03 n | 9.19 ± 0.54 d-g | 6.38 ± 0.54 h–l | 4.43 ± 0.29 e–g | 18.73 | Light brown |
| GR-07 P.v. | 61.56 ± 0.25 u | 1.25 ± 0.00 c–i | 16.53 ± 0.09 kl | 12.40 ± 0.98 qr | 7.21 ± 0.58 q | 4.72 ± 0.47 i–l | 31.63 | Light brown |
| GR-08 P.l. | 57.20 ± 0.25 t | 1.24 ± 0.01 c–i | 10.73 ± 0.25 g | 15.13 ± 1.18 t | 10.66 ± 0.79 tu | 4.13 ± 0.33 d–f | 50.91 | Light brown |
| GR-09 P.l. | 65.85 ± 0.13 v | 1.31 ± 0.01 d-i | 13.88 ± 0.08 h | 15.59 ± 1.50 t | 10.91 ± 0.98 u | 4.33 ± 0.70 d–f | 56.40 | White |
| GR-10 P.v. | 22.10 ± 0.31 fg | 1.44 ± 0.01 hi | 1.56 ± 0.05 cd | 8.70 ± 0.76 b–d | 5.84 ± 0.58 de | 4.01 ± 0.39 b | 16.26 | Black–purple |
| GR-11 P.v. | 32.31 ± 0.47 n | 0.61 ± 0.04 b–i | 20.06 ± 0.16 no | 10.27 ± 0.61 k–n | 6.89 ± 0.51 op | 4.63 ± 050 g–k | 26.92 | Red |
| GR-12 P.v. | 73.95 ± 0.13 y | 1.48 ± 0.00 hi | 10.85 ± 0.03 g | 8.83 ± 0.74 b–d | 6.21 ± 0.46 f–j | 5.11 ± 0.42 no | 21.58 | White |
| GR-13 P.v. | 29.10 ± 0.39 m | 0.33 ± 0.03 c | 17.30 ± 0.47 lm | 10.84 ± 0.84 op | 5.70 ± 0.39 b–d | 4.61 ± 0.43 g–j | 21.47 | Red |
| GR-14 P.v. | 18.47 ± 0.07 bc | 1.50 ± 0.01 i | 0.84 ± 0.04 a-c | 10.20 ± 0.91 k–n | 6.28 ± 0.61 g–k | 4.70 ± 0.44 h–l | 23.75 | Black–purple |
| GR-15 P.c. | 37.05 ± 0.68 o | 0.73 ± 0.02 b–i | 11.65 ± 0.47 g | 12.71 ± 1.28 r | 9.60 ± 0.91 t | 6.13 ± 0.75 p | 53.96 | Purple |
| GR-16 P.v. | 42.74 ± 0.29 q | 0.94 ± 0.01 b–i | 13.37 ± 0.13 h | 10.51 ± 1.21 m–o | 6.67 ± 0.65 m–o | 4.88 ± 0.53 k–n | 27.00 | Purple |
| GR-17 P.v. | 23.18 ± 0.17 g–i | 0.38 ± 0.01 b–d | 20.05 ± 0.24 no | 11.95 ± 1.28 q | 7.89 ± 0.82 r | 5.94 ± 0.79 p | 39.63 | Red |
| GR-18 P.v. | 55.06 ± 0.82 s | 1.06 ± 0.01 b–i | 17.63 ± 0.10 m | 14.17 ± 1.16 s | 8.93 ± 0.70 s | 6.63 ± 0.70 q | 57.95 | Light brown |
| OX-01 P.v. | 19.82 ± 0.04 d | −1.11 ± 0.09 a | 1.27 ± 0.05 ab | 10.30 ± 0.74 k–n | 6.43 ± 0.42 j–m | 4.54 ± 0.35 f–i | 22.92 | Black–blue |
| OX-02 P.v. | 17.47 ± 0.27 ab | −1.50 ± 0.07 a | 0.34 ± 0.03 ab | 9.48 ± 0.68 e–i | 6.10 ± 0.38 e–h | 4.15 ± 0.35 b–d | 18.96 | Black–blue |
| OX-03 P.v. | 19.22 ± 0.09 cd | 1.44 ± 0.09 hi | 0.23 ± 0.01 a | 9.53 ± 0.72 f–i | 5.83 ± 0.43 d–e | 4.21 ± 0.38 b–e | 18.40 | Black–purple |
| OX-04 P.v. | 17.05 ± 0.09 a | −1.45 ± 0.06 a | 0.40 ± 0.15 ab | 9.56 ± 0.74 g–i | 6.14 ± 0.44 e–i | 4.58 ± 0.37 f–i | 20.92 | Black–blue |
| OX-05 P.v. | 18.93 ± 0.25 cd | 1.37 ± 0.02 f–i | 1.48 ± 0.13 cd | 8.82 ± 0.90 b–d | 5.23 ± 0.46 a | 3.67 ± 0.44 a | 12.23 | Black–purple |
| OX-06 P.v. | 66.08 ± 0.29 v | 1.32 ± 0.00 e–i | 21.79 ± 0.79 pq | 10.40 ± 0.88 l–o | 7.11 ± 0.70 pq | 5.26 ± 0.36 o | 27.67 | Light brown |
| OX-07 P.v. | 69.23 ± 0.16 w | 1.43 ± 0.00 hi | 14.37 ± 0.26 hi | 9.39 ± 064 e–h | 5.97 ± 0.33 d–g | 4.97 ± 0.30 l–n | 21.44 | White |
| OX-08 P.v. | 49.47 ± 0.84 r | 1.03 ± 0.01 b–i | 21.01 ± 0.18 op | 9.70 ± 086 h–j | 5.66 ± 0.45 b–d | 4.33 ± 0.57 d–f | 17.93 | Light brown |
| OX-09 P.v. | 20.10 ± 0.47 de | 1.36 ± 0.01 f–i | 1.25 ± 0.11 a-d | 9.58 ± 0.70 g–i | 6.58 ± 0.46 l–o | 4.85 ± 0.36 j–n | 21.87 | Black–purple |
| OX-10 P.v. | 28.13 ± 0.44 m | 0.55 ± 0.02 b-h | 19.57 ± 0.26 n | 10.53 ± 0.59 m–o | 5.77 ± 0.50 cd | 4.79 ± 0.38 i–m | 22.39 | Red |
| OX-11 P.v. | 71.60 ± 0.03 x | 1.49 ± 0.00 i | 17.79 ± 0.04 m | 9.83 ± 0.63 h–k | 5.32 ± 0.37 a | 4.13 ± 0.31 b–d | 17.31 | White |
| OX-12 P.v. | 73.03 ± 0.11 y | 1.46 ± 0.01 hi | 16.18 ± 0.26 jk | 10.47 ± 0.76 m–o | 6.56 ± 0.45 l–n | 4.80 ± 0.46 i–m | 25.64 | White |
| OX-13 P.v. | 22.87 ± 0.39 gh | 0.46 ± 0.01 b–g | 15.61 ± 0.22 jk | 10.11 ± 0.52 j–m | 6.54 ± 0.35 l–n | 4.71 ± 0.39 h–l | 23.25 | Red |
| OX-14 P.v. | 71.50 ± 0.13 x | 1.38 ± 0.00 g–i | 15.39 ± 0.09 ij | 7.83 ± 0.55 a | 5.46 ± 0.37 a–c | 4.43 ± 0.45 e–g | 14.89 | White |
| OX-15 P.v. | 22.51 ± 0.01 f–h | 0.44 ± 1.73 b–f | 1.50 ± 0.14 cd | 8.75 ± 0.87 b–d | 5.73 ± 0.45 cd | 4.04 ± 0.40 b | 17.86 | Black |
| OX-16 P.v. | 24.90 ± 0.23 jk | 0.21 ± 0.03 b | 2.15 ± 0.07 de | 8.99 ± 0.78 b–e | 5.39 ± 0.38 ab | 4.44 ± 0.51 e–h | 15.19 | Black |
| OX-17 P.v. | 25.31 ± 0.20 kl | 1.44 ± 0.04 hi | 2.78 ± 0.24 e | 9.05 ± 0.78 c–f | 5.33 ± 0.41 a | 4.05 ± 0.39 bc | 16.31 | Black–purple |
| OX-18 P.v. | 23.60 ± 0.23 h–j | 0.17 ± 0.14 b | 2.23 ± 0.52 de | 11.22 ± 1.21 p | 6.80 ± 0.57 n–p | 5.25 ± 0.79 o | 29.07 | Black |
| OX-19 P.v. | 28.68 ± 0.49 m | 0.77 ± 0.14 b | 13.83 ± 0.39 f | 9.90 ± 0.86 i–l | 5.74 ± 0.52 cd | 4.80 ± 0.63 i–m | 20.97 | Dark brown |
| OX-20 P.c. | 26.25 ± 0.58 l | 0.41 ± 0.10 b–e | 3.99 ± 0.39 f | 14.09 ± 1.59 s | 10.37 ± 1.12 t | 6.67 ± 0.96 q | 67.29 | Black |
| Species | Moisture | Ashes | Fat | Protein | Carbohydrates | Fiber | Food Energy (Kcal/100 g) |
|---|---|---|---|---|---|---|---|
| P. coccineus | 11.20 ± 0.74 b | 4.27 ± 0.50 c | 1.87 ± 0.44 ab | 17.28 ± 2.06 a | 65.39 ± 1.39 b | 4.18 ± 0.33 a | 424.54 ± 9.96 a |
| P. lunatus | 9.62 ± 0.77 a | 3.48 ± 0.19 a | 1.60 ± 0.08 a | 19.68 ± 0.58 b | 65.62 ± 0.85 b | 5.21 ± 0.20 a | 445.98 ± 8.13 b |
| P. vulgaris | 10.16 ± 1.49 b | 3.78 ± 0.25 b | 2.22 ± 0.38 b | 20.48 ± 2.78 b | 62.57 ± 0.43 a | 5.02 ± 1.76 a | 444.78 ± 2.45 b |
| Variables Evaluated | P. coccineus | P. lunatus | P. vulgaris |
|---|---|---|---|
| Total phenolic content (mg GAE/g) | 472.03 ± 0.17 a | 604.85 ± 0.35 b | 440.40 ± 0.61 a |
| Flavonoid content (mg QE/g) | 12.38 ± 0.45 c | 4.99 ± 0.94 a | 8.82 ± 0.77 b |
| Anthocyanin content (mg C-3G/g seed coat) | 0.53 ± 0.06 a | 1.71 ± 1.32 b | 2.69 ± 1.24 c |
| Tannin content (mg CAE/g) | 1.01 ± 0.65 ab | 0.59 ± 0.70 a | 1.37 ± 0.98 b |
| Phytic acid content (mg/100 g bean flour) | 0.58 ± 0.22 b | 0.43 ± 0.32 ab | 0.39 ± 0.56 a |
| Oligosaccharid content (mg/100 g bean flour) | 6.44 ± 0.04 b | 3.68 ± 0.11 a | 3.92 ± 0.27 a |
| Lectin (HAU/mg protein) | 1.33 ± 0.12 a | 1.52 ± 0.03 ab | 1.63 ± 0.18 b |
| Trypsin inhibitory activity (TIU/g bean flour) | 15.97 ± 0.03 ab | 18.05 ± 0.01 b | 13.19 ± 0.32 a |
| Radical scavenging activity (% ARSA) | 80.57 ± 0.08 a | 84.18 ± 0.01 ab | 86.43 ± 0.06 b |
| Radical scavenging activity (% DRSA) | 56.47 ± 0.08 a | 88.08 ± 0.02 ab | 89.28 ± 0.04 b |
| Correlation | r | p |
|---|---|---|
| Luminosity and hue | 0.47 | 0.003 |
| Luminosity and chromaticity | 0.64 | 0.000 |
| Luminosity and phenols | −0.53 | 0.001 |
| Luminosity and flavonoids | −0.72 | 0.000 |
| Luminosity and tannins | −0.67 | 0.000 |
| Luminosity and DPPH | −0.49 | 0.000 |
| Luminosity and anthocyanins | −0.66 | 0.000 |
| Hue and flavonoids | −0.39 | 0.017 |
| Hue and tannins | −0.56 | 0.000 |
| Chromaticity and tannins | −0.49 | 0.002 |
| Chromaticity and DPPH | −0.59 | 0.000 |
| Chromaticity and anthocyanins | −0.80 | 0.000 |
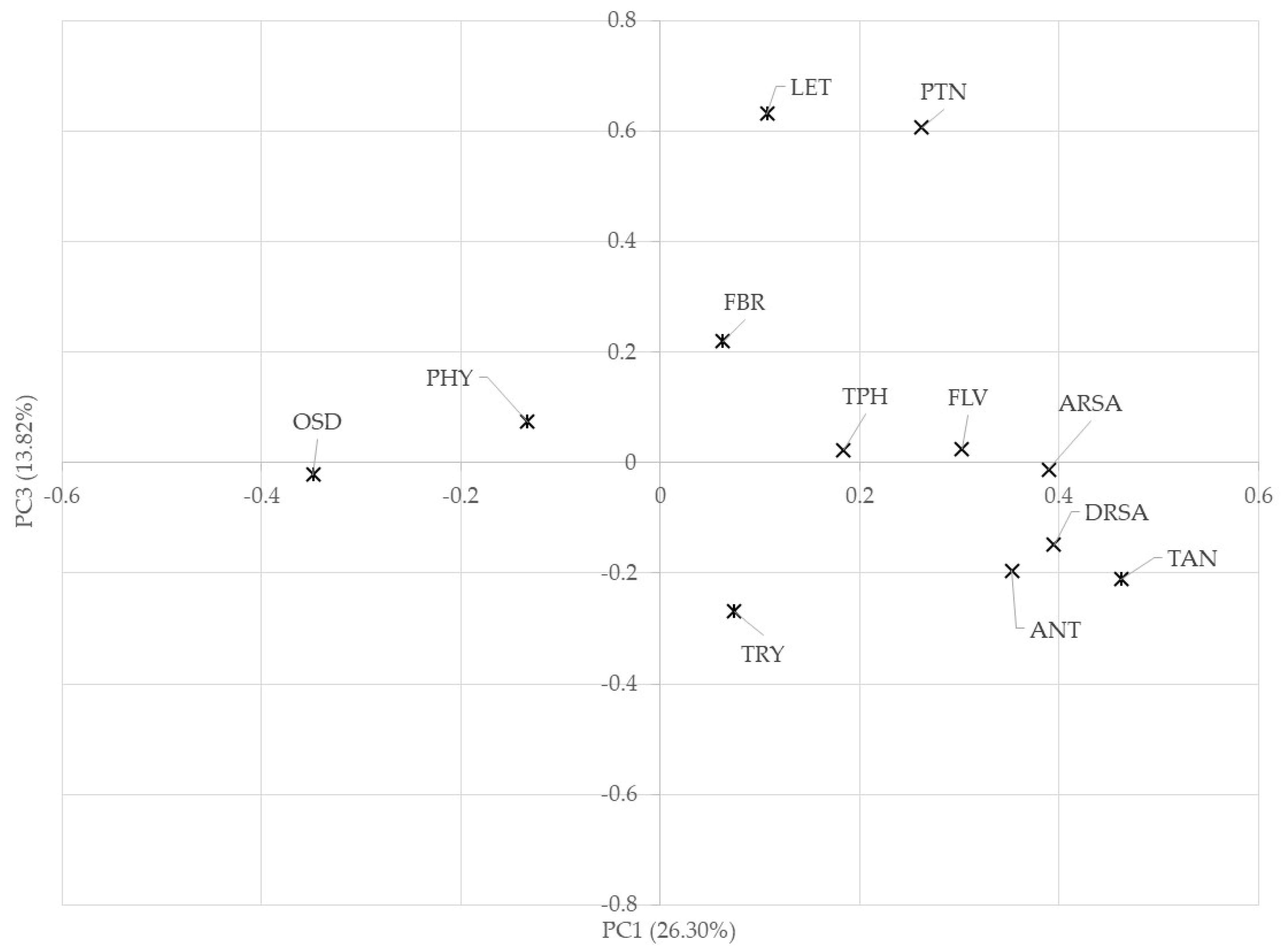
| Phenolic compounds and antioxidant activity | Sample | Specie | PCA1 | PCA3 |  |
| OX-01 | P. vulgaris | 1.83 | −1.79 | ||
| OX-02 | P. vulgaris | 1.47 | −1.72 | ||
| OX-03 | P. vulgaris | 1.63 | −1.40 | ||
| OX-04 | P. vulgaris | 4.52 | 0.76 | ||
| OX-05 | P. vulgaris | 2.54 | −0.94 | ||
| OX-09 | P. vulgaris | 2.51 | 0.06 | ||
| OX-10 | P. vulgaris | 3.05 | −1.22 | ||
| OX-13 | P. vulgaris | 1.90 | 1.30 | ||
| OX-16 | P. vulgaris | 1.31 | 0.85 | ||
| GR-01 | P. lunatus | 0.50 | −0.46 | ||
| GR-04 | P. vulgaris | 1.63 | 1.47 | ||
| GR-05 | P. vulgaris | 1.74 | 1.43 | ||
| GR-10 | P. vulgaris | 0.98 | 0.33 | ||
| GR-14 | P. vulgaris | 1.45 | −0.09 | ||
| Oligosaccharides and phytic acid | OX-06 | P. vulgaris | −1.31 | 1.22 |  |
| OX-07 | P. vulgaris | −0.91 | 0.11 | ||
| OX-08 | P. vulgaris | −1.76 | 0.86 | ||
| OX-12 | P. vulgaris | −1.12 | −0.02 | ||
| OX-14 | P. vulgaris | −2.39 | 0.33 | ||
| OX-19 | P. vulgaris | −1.45 | 0.20 | ||
| OX-20 | P. coccineus | −1.45 | −0.88 | ||
| GR-03 | P. vulgaris | −1.78 | −0.61 | ||
| GR-06 | P. vulgaris | −2.36 | 1.61 | ||
| GR-07 | P. vulgaris | −1.88 | −0.98 | ||
| GR-08 | P. lunatus | −1.06 | −0.51 | ||
| GR-09 | P. lunatus | −1.20 | 0.03 | ||
| GR-15 | P. coccineus | −2.93 | −1.75 | ||
| GR-17 | P. vulgaris | −0.70 | −0.01 | ||
| GR-18 | P. vulgaris | −1.98 | −1.49 | ||
| Protein, fiber, lectin | OX-11 | P. vulgaris | −0.54 | 0.80 |  |
| GR-02 | P. vulgaris | −0.98 | 2.52 | ||
| GR-12 | P. vulgaris | −1.32 | 2.98 | ||
| GR-13 | P. vulgaris | 1.11 | 2.61 | ||
| Inhibitory trypsin activity | OX-15 | P. vulgaris | 0.90 | −1.63 |  |
| OX-17 | P. vulgaris | −0.10 | −0.85 | ||
| OX-18 | P. vulgaris | 0.00 | −0.05 | ||
| GR-11 | P. vulgaris | −0.47 | −1.52 | ||
| GR-16 | P. vulgaris | −1.38 | −1.52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alcázar-Valle, M.; García-Morales, S.; Mojica, L.; Morales-Hernández, N.; Sánchez-Osorio, E.; Flores-López, L.; Enríquez-Vara, J.N.; Lugo-Cervantes, E. Nutritional, Antinutritional Compounds and Nutraceutical Significance of Native Bean Species (Phaseolus spp.) of Mexican Cultivars. Agriculture 2021, 11, 1031. https://doi.org/10.3390/agriculture11111031
Alcázar-Valle M, García-Morales S, Mojica L, Morales-Hernández N, Sánchez-Osorio E, Flores-López L, Enríquez-Vara JN, Lugo-Cervantes E. Nutritional, Antinutritional Compounds and Nutraceutical Significance of Native Bean Species (Phaseolus spp.) of Mexican Cultivars. Agriculture. 2021; 11(11):1031. https://doi.org/10.3390/agriculture11111031
Chicago/Turabian StyleAlcázar-Valle, Montserrat, Soledad García-Morales, Luis Mojica, Norma Morales-Hernández, Ever Sánchez-Osorio, Lourdes Flores-López, Jhony Navat Enríquez-Vara, and Eugenia Lugo-Cervantes. 2021. "Nutritional, Antinutritional Compounds and Nutraceutical Significance of Native Bean Species (Phaseolus spp.) of Mexican Cultivars" Agriculture 11, no. 11: 1031. https://doi.org/10.3390/agriculture11111031