
Role of Halotolerant Plant Growth-Promoting Rhizobacteria in Mitigating Salinity Stress: Recent Advances and Possibilities
 and
and 
Abstract
:1. Introduction
2. HT-PGPR: Diversity and Their Effect on Crop Production
3. HT-PGPR and Their Effects in Mitigating Salt Stress in Crops
4. Plant Growth-Promoting Mechanisms by HT-PGPR
4.1. HT-PGPR Mediated Increased Availability of Soil Nutrients
4.2. HT-PGPR Mediation Increase Availability of Indole-3-Acetic Acid
4.3. HT-PGPR Modulations of Ethylene
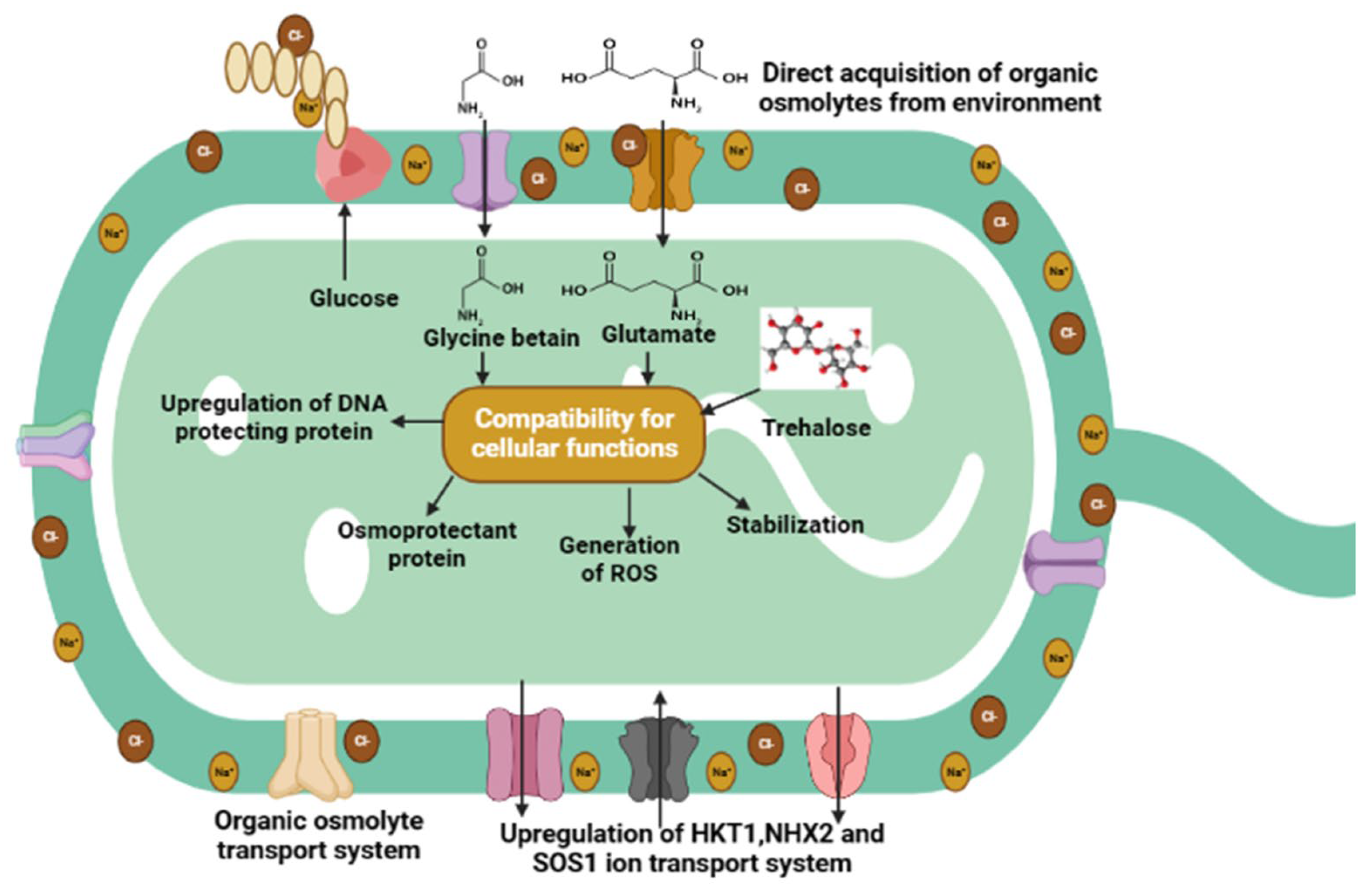
4.4. HT-PGPR Improves Osmoprotectants, Compatible Solutes, and Ion Homeostasis
4.5. Production of Exopolysaccharides and Volatile Organic Compounds by HT-PGPR
5. Future Prospects
6. Conclusions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Myo, E.M.; Ge, B.; Ma, J.; Cui, H.; Liu, B.; Shi, L.; Jiang, M.; Zhang, K. Indole-3-Acetic Acid Production by Streptomyces Fradiae NKZ-259 and Its Formulation to Enhance Plant Growth. BMC Microbiol. 2019, 19, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharti, N.; Barnawal, D.; Awasthi, A.; Yadav, A.; Kalra, A. Plant Growth Promoting Rhizobacteria Alleviate Salinity Induced Negative Effects on Growth, Oil Content and Physiological Status in Mentha Arvensis. Acta Physiol. Plant 2014, 36, 45–60. [Google Scholar] [CrossRef]
- Morris, J.; González, J.E. The Novel Genes EmmABC Are Associated with Exopolysaccharide Production, Motility, Stress Adaptation, and Symbiosis in Sinorhizobium Meliloti. J. Bacteriol. 2009, 191, 5890–5900. [Google Scholar] [CrossRef] [Green Version]
- Ladeiro, B. Saline Agriculture in the 21st Century: Using Salt Contaminated Resources to Cope Food Requirements. J. Bot. 2012, 2012, 310705. [Google Scholar] [CrossRef]
- Shahid, S.A.; Zaman, M.; Heng, L. Soil Salinity: Historical Perspectives and a World Overview of the Problem. In Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Zaman, M., Shahid, S.A., Heng, L., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 43–53. ISBN 978-3-319-96190-3. [Google Scholar]
- Hernandez Fernandez, M.T.; Mataix-Solera, J.; Lichner, L.; Štekaurová, V.; Zaujec, A.; Garcia Izquierdo, C. Assessing the Microbiological, Biochemical, Soil-Physical and Hydrological Effects of Amelioration of Degraded Soils in Semiarid Spain. Biologia 2007, 62, 542–546. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Singh, J.S.; Saxena, A.K.; Singh, D.P. Impact of PGPR Inoculation on Growth and Antioxidant Status of Wheat under Saline Conditions. Plant Biol. 2012, 14, 605–611. [Google Scholar] [CrossRef]
- Orhan, F. Alleviation of Salt Stress by Halotolerant and Halophilic Plant Growth-Promoting Bacteria in Wheat (Triticum aestivum). Braz. J. Microbiol. 2016, 47, 621. [Google Scholar] [CrossRef] [Green Version]
- Al-Yassin, A. Adverse Effects of Salinity on Citrus. A review paper. J. Cent Eur. Agric. 2004, 4, 263–272. [Google Scholar]
- Egamberdieva, D.; Wirth, S.; Bellingrath-Kimura, S.D.; Mishra, J.; Arora, N.K. Salt-Tolerant Plant Growth Promoting Rhizobacteria for Enhancing Crop Productivity of Saline Soils. Front. Microbiol. 2019, 10, 2791. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Tanna, B. Halophytes: Potential Resources for Salt Stress Tolerance Genes and Promoters. Front. Plant Sci. 2017, 8, 829. [Google Scholar] [CrossRef]
- Rahman, M.M.; Mostofa, M.G.; Keya, S.S.; Siddiqui, M.N.; Ansary, M.M.U.; Das, A.K.; Rahman, M.A.; Tran, L.S.-P. Adaptive Mechanisms of Halophytes and Their Potential in Improving Salinity Tolerance in Plants. Int. J. Mol. Sci. 2021, 22, 10733. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.M.; Sako, K.; Matsui, A.; Suzuki, Y.; Mostofa, M.G.; Ha, C.V.; Tanaka, M.; Tran, L.-S.P.; Habu, Y.; Seki, M. Ethanol Enhances High-Salinity Stress Tolerance by Detoxifying Reactive Oxygen Species in Arabidopsis thaliana and Rice. Front. Plant Sci. 2017, 8, 1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, O.X.; Ronald, P.C. Genetic Engineering for Disease Resistance in Plants: Recent Progress and Future Perspectives. Plant Physiol. 2019, 180, 26–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Singh, P.; Sharma, R. Microorganism as a Tool of Bioremediation Technology for Cleaning Environment: A Review. Proc. Int. Acad. Ecol. Environ. Sci. 2014, 4, 1–6. [Google Scholar]
- Gopalakrishnan, S.; Sathya, A.; Vijayabharathi, R.; Varshney, R.K.; Gowda, C.L.L.; Krishnamurthy, L. Plant Growth Promoting Rhizobia: Challenges and Opportunities. 3 Biotech 2015, 5, 355–377. [Google Scholar] [CrossRef] [Green Version]
- Abbas, R.; Rasul, S.; Aslam, K.; Baber, M.; Shahid, M.; Mubeen, F.; Naqqash, T. Halotolerant PGPR: A Hope for Cultivation of Saline Soils. J. King Saud Univ. Sci. 2019, 31, 1195–1201. [Google Scholar] [CrossRef]
- Sharma, K.; Sharma, S.; Vaishnav, A.; Jain, R.; Singh, D.; Singh, H.B.; Goel, A.; Singh, S. Salt-Tolerant PGPR Strain Priestia Endophytica SK1 Promotes Fenugreek Growth under Salt Stress by Inducing Nitrogen Assimilation and Secondary Metabolites. J. Appl. Microbiol. 2022, 133, 2802–2813. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd_Allah, E.F.; Hashem, A. Phytohormones and Beneficial Microbes: Essential Components for Plants to Balance Stress and Fitness. Front. Microbiol. 2017, 8, 2104. [Google Scholar] [CrossRef]
- Sunita, K.; Mishra, I.; Mishra, J.; Prakash, J.; Arora, N.K. Secondary Metabolites From Halotolerant Plant Growth Promoting Rhizobacteria for Ameliorating Salinity Stress in Plants. Front. Microbiol. 2020, 11, 567768. [Google Scholar] [CrossRef]
- Hernández-Canseco, J.; Bautista-Cruz, A.; Sánchez-Mendoza, S.; Aquino-Bolaños, T.; Sánchez-Medina, P.S. Plant Growth-Promoting Halobacteria and Their Ability to Protect Crops from Abiotic Stress: An Eco-Friendly Alternative for Saline Soils. Agronomy 2022, 12, 804. [Google Scholar] [CrossRef]
- Ruginescu, R.; Gomoiu, I.; Popescu, O.; Cojoc, R.; Neagu, S.; Lucaci, I.; Batrinescu-Moteau, C.; Enache, M. Bioprospecting for Novel Halophilic and Halotolerant Sources of Hydrolytic Enzymes in Brackish, Saline and Hypersaline Lakes of Romania. Microorganisms 2020, 8, 1903. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Kim, M.-S.; Sun, Y.; Dowd, S.E.; Shi, H.; Paré, P.W. Soil Bacteria Confer Plant Salt Tolerance by Tissue-Specific Regulation of the Sodium Transporter HKT1. Mol. Plant Microbe Interact. 2008, 21, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The Role of Na+ and K+ Transporters in Salt Stress Adaptation in Glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.; Peng, F.; Xue, X.; You, Q.; Zhang, W.; Wang, T.; Huang, C. The Growth Promotion of Two Salt-Tolerant Plant Groups with PGPR Inoculation: A Meta-Analysis. Sustainability 2019, 11, 378. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.-F.; Chaparro, J.M.; Reardon, K.F.; Zhang, R.; Shen, Q.; Vivanco, J.M. Rhizosphere Interactions: Root Exudates, Microbes, and Microbial Communities. Botany 2014, 92, 267–275. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Zahir, Z.A.; Naveed, M.; Nawaz, S. Mitigation of Salinity-Induced Negative Impact on the Growth and Yield of Wheat by Plant Growth-Promoting Rhizobacteria in Naturally Saline Conditions. Ann. Microbiol. 2013, 63, 225–232. [Google Scholar] [CrossRef]
- Phour, M.; Sindhu, S.S. Amelioration of Salinity Stress and Growth Stimulation of Mustard (Brassica juncea L.) by Salt-Tolerant Pseudomonas Species. Appl. Soil Ecol. 2020, 149, 103518. [Google Scholar] [CrossRef]
- Aslam, F.; Ali, B. Halotolerant Bacterial Diversity Associated with Suaeda fruticosa (L.) Forssk. Improved Growth of Maize under Salinity Stress. Agronomy 2018, 8, 131. [Google Scholar] [CrossRef] [Green Version]
- Zahir, Z.A.; Ghani, U.; Naveed, M.; Nadeem, S.M.; Asghar, H.N. Comparative Effectiveness of Pseudomonas and Serratia Sp. Containing ACC-Deaminase for Improving Growth and Yield of Wheat (Triticum aestivum L.) under Salt-Stressed Conditions. Arch. Microbiol. 2009, 191, 415–424. [Google Scholar] [CrossRef]
- Kasotia, A.; Varma, A.; Tuteja, N.; Choudhary, D.K. Amelioration of Soybean Plant from Saline-Induced Condition by Exopolysaccharide Producing Pseudomonas-Mediated Expression of High Affinity K + -Transporter (HKT1) Gene. Curr. Sci. 2016, 111, 1961–1967. [Google Scholar] [CrossRef]
- Kwon, S.-W.; Park, J.-Y.; Kim, J.-S.; Kang, J.-W.; Cho, Y.-H.; Lim, C.-K.; Parker, M.A.; Lee, G.-B. Phylogenetic Analysis of the Genera Bradyrhizobium, Mesorhizobium, Rhizobium and Sinorhizobium on the Basis of 16S RRNA Gene and Internally Transcribed Spacer Region Sequences. Int. J. Syst. Evol. Microbiol. 2005, 55, 263–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, Y.S.; Ryu, C.-M.; Lee, S.; Park, H.B.; Han, K.S.; Lee, J.H.; Lee, K.; Chung, W.S.; Jeong, M.-J.; Kim, H.K.; et al. Proteome Analysis of Arabidopsis Seedlings Exposed to Bacterial Volatiles. Planta 2010, 232, 1355–1370. [Google Scholar] [CrossRef] [PubMed]
- Baek, D.; Rokibuzzaman, M.; Khan, A.; Kim, M.C.; Park, H.J.; Yun, D.; Chung, Y.R. Plant-Growth Promoting Bacillus Oryzicola YC7007 Modulates Stress-Response Gene Expression and Provides Protection From Salt Stress. Front. Plant Sci. 2020, 10, 1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, T.N.; Tran, B.T.H.; Van Bui, L.; Hoang, M.T.T. Plant Growth-Promoting Rhizobacterium Pseudomonas PS01 Induces Salt Tolerance in Arabidopsis thaliana. BMC Res. Notes 2019, 12, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabhi, N.E.H.; Silini, A.; Cherif-Silini, H.; Yahiaoui, B.; Lekired, A.; Robineau, M.; Esmaeel, Q.; Jacquard, C.; Vaillant-Gaveau, N.; Clément, C.; et al. Pseudomonas Knackmussii MLR6, a Rhizospheric Strain Isolated from Halophyte, Enhances Salt Tolerance in Arabidopsis thaliana. J. Appl. Microbiol. 2018, 125, 1836–1851. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, Y.; Wu, G.; Veronican Njeri, K.; Shen, Q.; Zhang, N.; Zhang, R. Induced Maize Salt Tolerance by Rhizosphere Inoculation of Bacillus Amyloliquefaciens SQR9. Physiol. Plant 2016, 158, 34–44. [Google Scholar] [CrossRef]
- Pinedo, I.; Ledger, T.; Greve, M.; Poupin, M.J. Burkholderia Phytofirmans PsJN Induces Long-Term Metabolic and Transcriptional Changes Involved in Arabidopsis thaliana Salt Tolerance. Front. Plant Sci. 2015, 6, 466. [Google Scholar] [CrossRef] [Green Version]
- Sukweenadhi, J.; Kim, Y.-J.; Choi, E.-S.; Koh, S.-C.; Lee, S.-W.; Kim, Y.-J.; Yang, D.C. Paenibacillus Yonginensis DCY84T Induces Changes in Arabidopsis thaliana Gene Expression against Aluminum, Drought, and Salt Stress. Microbiol. Res. 2015, 172, 7–15. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Sukweenadhi, J.; Seok, J.W.; Kang, C.H.; Choi, E.-S.; Subramaniyam, S.; Yang, D.C. Complete Genome Sequence of Paenibacillus Yonginensis DCY84T, a Novel Plant Symbiont That Promotes Growth via Induced Systemic Resistance. Stand. Genom. Sci. 2017, 12, 63. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Jang, Y.-J.; Lee, S.-M.; Oh, B.-T.; Chae, J.-C.; Lee, K.-J. Alleviation of Salt Stress by Enterobacter Sp. EJ01 in Tomato and Arabidopsis Is Accompanied by Up-Regulation of Conserved Salinity Responsive Factors in Plants. Mol. Cells 2014, 37, 109–117. [Google Scholar] [CrossRef]
- Qurashi, A.W.; Sabri, A.N. Bacterial Exopolysaccharide and Biofilm Formation Stimulate Chickpea Growth and Soil Aggregation under Salt Stress. Braz. J. Microbiol. 2012, 43, 1183–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Jan, R.; Kang, S.-M.; Kim, K.-M.; Lee, I.-J. Rhizobacteria AK1 Remediates the Toxic Effects of Salinity Stress via Regulation of Endogenous Phytohormones and Gene Expression in Soybean. Biochem. J. 2019, 476, 2393–2409. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Adhikari, A.; Jan, R.; Ali, S.; Imran, M.; Kim, K.-M.; Lee, I.-J. Halotolerant Rhizobacterial Strains Mitigate the Adverse Effects of NaCl Stress in Soybean Seedlings. Biomed. Res. Int. 2019, 2019, 9530963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaishnav, A.; Kumari, S.; Jain, S.; Varma, A.; Tuteja, N.; Choudhary, D.K. PGPR-Mediated Expression of Salt Tolerance Gene in Soybean through Volatiles under Sodium Nitroprusside: PGPR-Mediated Amelioration of Soybean under Salt Stress. J. Basic Microbiol. 2016, 56, 1274–1288. [Google Scholar] [CrossRef] [PubMed]
- Vaishnav, A.; Kumari, S.; Jain, S.; Varma, A.; Choudhary, D.K. Putative Bacterial Volatile-Mediated Growth in Soybean (Glycine max L. Merrill) and Expression of Induced Proteins under Salt Stress. J. Appl. Microbiol. 2015, 119, 539–551. [Google Scholar] [CrossRef]
- Ma, Y.; Rajkumar, M.; Oliveira, R.S.; Zhang, C.; Freitas, H. Potential of Plant Beneficial Bacteria and Arbuscular Mycorrhizal Fungi in Phytoremediation of Metal-Contaminated Saline Soils. J. Hazard. Mater. 2019, 379, 120813. [Google Scholar] [CrossRef]
- Kiani, M.Z.; Ali, A.; Sultan, T.; Ahmad, R.; Hydar, S.I. Plant Growth Promoting Rhizobacteria Having 1-Aminocyclopropane-1-Carboxylic Acid Deaminase to Induce Salt Tolerance in Sunflower (Helianthus annus L.). Nat. Resour. 2015, 6, 391–397. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, M.B.; Hidri, R.; Talbi-Zribi, O.; Taamalli, W.; Abdelly, C.; Djébali, N. Auxin and Proline Producing Rhizobacteria Mitigate Salt-Induced Growth Inhibition of Barley Plants by Enhancing Water and Nutrient Status. S. Afr. J. Bot. 2020, 128, 209–217. [Google Scholar] [CrossRef]
- Sultana, S.; Paul, S.C.; Parveen, S.; Alam, S.; Rahman, N.; Jannat, B.; Hoque, S.; Rahman, M.T.; Karim, M.M. Isolation and Identification of Salt-Tolerant Plant-Growth-Promoting Rhizobacteria and Their Application for Rice Cultivation under Salt Stress. Can. J. Microbiol. 2020, 66, 144–160. [Google Scholar] [CrossRef]
- Chauhan, P.S.; Lata, C.; Tiwari, S.; Chauhan, A.S.; Mishra, S.K.; Agrawal, L.; Chakrabarty, D.; Nautiyal, C.S. Transcriptional Alterations Reveal Bacillus Amyloliquefaciens-Rice Cooperation under Salt Stress. Sci. Rep. 2019, 9, 11912. [Google Scholar] [CrossRef] [Green Version]
- Nghia, N.K.; Tien, T.T.M.; Oanh, N.T.K.; Nuong, N.H.K. Isolation and Characterization of Indole Acetic Acid Producing Halophilic Bacteria from Salt Affected Soil of Rice–Shrimp Farming System in the Mekong Delta, Vietnam. Agric. For. Fish. 2017, 6, 69. [Google Scholar] [CrossRef] [Green Version]
- Jha, Y.; Subramanian, R.B.; Patel, S. Combination of Endophytic and Rhizospheric Plant Growth Promoting Rhizobacteria in Oryza Sativa Shows Higher Accumulation of Osmoprotectant against Saline Stress. Acta Physiol. Plant 2011, 33, 797–802. [Google Scholar] [CrossRef]
- Niu, S.-Q.; Li, H.-R.; Paré, P.W.; Aziz, M.; Wang, S.-M.; Shi, H.; Li, J.; Han, Q.-Q.; Guo, S.-Q.; Li, J.; et al. Induced Growth Promotion and Higher Salt Tolerance in the Halophyte Grass Puccinellia Tenuiflora by Beneficial Rhizobacteria. Plant Soil 2016, 407, 217–230. [Google Scholar] [CrossRef]
- Kang, S.-M.; Shahzad, R.; Bilal, S.; Khan, A.L.; Park, Y.-G.; Lee, K.-E.; Asaf, S.; Khan, M.A.; Lee, I.-J. Indole-3-Acetic-Acid and ACC Deaminase Producing Leclercia Adecarboxylata MO1 Improves Solanum lycopersicum L. Growth and Salinity Stress Tolerance by Endogenous Secondary Metabolites Regulation. BMC Microbiol. 2019, 19, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaishnav, A.; Singh, J.; Singh, P.; Rajput, R.S.; Singh, H.B.; Sarma, B.K. Sphingobacterium Sp. BHU-AV3 Induces Salt Tolerance in Tomato by Enhancing Antioxidant Activities and Energy Metabolism. Front. Microbiol. 2020, 11, 443. [Google Scholar] [CrossRef]
- Yan, J.; Smith, M.D.; Glick, B.R.; Liang, Y. Effects of ACC Deaminase Containing Rhizobacteria on Plant Growth and Expression of Toc GTPases in Tomato (Solanum lycopersicum) under Salt Stress. Botany 2014, 92, 775–781. [Google Scholar] [CrossRef]
- Han, Q.-Q.; Lü, X.-P.; Bai, J.-P.; Qiao, Y.; Paré, P.W.; Wang, S.-M.; Zhang, J.-L.; Wu, Y.-N.; Pang, X.-P.; Xu, W.-B.; et al. Beneficial Soil Bacterium Bacillus Subtilis (GB03) Augments Salt Tolerance of White Clover. Front. Plant Sci. 2014, 5, 525. [Google Scholar] [CrossRef]
- Shah, D.; Khan, M.S.; Aziz, S.; Ali, H.; Pecoraro, L. Molecular and Biochemical Characterization, Antimicrobial Activity, Stress Tolerance, and Plant Growth-Promoting Effect of Endophytic Bacteria Isolated from Wheat Varieties. Microorganisms 2021, 10, 21. [Google Scholar] [CrossRef]
- Vaishnav, A.; Kumar, R.; Singh, H.B.; Sarma, B.K. Extending the Benefits of PGPR to Bioremediation of Nitrile Pollution in Crop Lands for Enhancing Crop Productivity. Sci. Total Environ. 2022, 826, 154170. [Google Scholar] [CrossRef] [PubMed]
- Safdarian, M.; Askari, H.; Shariati, J.V.; Nematzadeh, G. Transcriptional Responses of Wheat Roots Inoculated with Arthrobacter Nitroguajacolicus to Salt Stress. Sci. Rep. 2019, 9, 1792. [Google Scholar] [CrossRef]
- Singh, R.P.; Jha, P.N. The Multifarious PGPR Serratia Marcescens CDP-13 Augments Induced Systemic Resistance and Enhanced Salinity Tolerance of Wheat (Triticum aestivum L.). PLoS ONE 2016, 11, e0155026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirhadi, M.; Enayatizamir, N.; Motamedi, H.; Sorkheh, K. Screening of Salt Tolerant Sugarcane Endophytic Bacteria with Potassium and Zinc for Their Solubilizing and Antifungal Activity. Biosci. Biotech. Res. Comm. 2016, 9, 530–538. [Google Scholar] [CrossRef]
- Ramadoss, D.; Lakkineni, V.K.; Bose, P.; Ali, S.; Annapurna, K. Mitigation of Salt Stress in Wheat Seedlings by Halotolerant Bacteria Isolated from Saline Habitats. Springerplus 2013, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alzahrani, S.M.; Ali, H.M.; Alayafi, A.A.; Ahmad, M. Serratia Liquefaciens KM4 Improves Salt Stress Tolerance in Maize by Regulating Redox Potential, Ion Homeostasis, Leaf Gas Exchange and Stress-Related Gene Expression. Int. J. Mol. Sci. 2018, 19, 3310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alla, M.M.N.; Hassan, N.M. A Possible Role for C4 Photosynthetic Enzymes in Tolerance of Zea mays to NaCl. Protoplasma 2012, 249, 1109–1117. [Google Scholar] [CrossRef] [PubMed]
- Habib, S.H.; Kausar, H.; Saud, H.M. Plant Growth-Promoting Rhizobacteria Enhance Salinity Stress Tolerance in Okra through ROS-Scavenging Enzymes. Biomed. Res. Int. 2016, 2016, 6284547. [Google Scholar] [CrossRef] [Green Version]
- Etesami, H.; Beattie, G.A. Mining Halophytes for Plant Growth-Promoting Halotolerant Bacteria to Enhance the Salinity Tolerance of Non-Halophytic Crops. Front. Microbiol. 2018, 9, 148. [Google Scholar] [CrossRef] [Green Version]
- Xin, C.-S.; Dong, H.-Z.; Luo, Z.; Tang, W.; Zhang, D.-M.; Li, W.-J.; Kong, X.-Q. Effects of N, P, and K Fertilizer Application on Cotton Growing in Saline Soil in Yellow River Delta. Acta Agron. Sin. 2010, 36, 1698–1706. [Google Scholar] [CrossRef]
- Thomine, S.; Lanquar, V. Iron Transport and Signaling in Plants. In Transporters and Pumps in Plant Signaling; Geisler, M., Venema, K., Eds.; Signaling and Communication in Plants; Springer: Berlin/Heidelberg, Germany, 2011; pp. 99–131. ISBN 978-3-642-14369-4. [Google Scholar]
- Fageria, N.; Gheyi, H.; Moreira, A. Nutrient Bioavailability in Salt Affected Soils. J. Plant Nutr. 2011, 34, 945–962. [Google Scholar] [CrossRef]
- Sharma, S.; Kulkarni, J.; Jha, B. Halotolerant Rhizobacteria Promote Growth and Enhance Salinity Tolerance in Peanut. Front. Microbiol. 2016, 7, 1600. [Google Scholar] [CrossRef] [Green Version]
- Teo, H.M.; Bhubalan, K.; Ci, M.S.; Ng, L.C. Setting a Plausible Route for Saline Soil-Based Crop Cultivations by Application of Beneficial Halophyte-Associated Bacteria: A Review. Microorganisms 2022, 10, 657. [Google Scholar] [CrossRef] [PubMed]
- Raymond, J.; Siefert, J.L.; Staples, C.R.; Blankenship, R.E. The Natural History of Nitrogen Fixation. Mol. Biol. Evol. 2004, 21, 541–554. [Google Scholar] [CrossRef] [PubMed]
- Hachiya, T.; Sakakibara, H. Interactions between Nitrate and Ammonium in Their Uptake, Allocation, Assimilation, and Signaling in Plants. J. Exp. Bot. 2017, 68, 2501–2512. [Google Scholar] [CrossRef] [PubMed]
- Parmar, P.; Sindhu, S.S. Potassium Solubilization by Rhizosphere Bacteria: Influence of Nutritional and Environmental Conditions. J. Microbiol. Res. 2013, 3, 25–31. [Google Scholar] [CrossRef]
- Yan, N.; Marschner, P.; Cao, W.; Zuo, C.; Qin, W. Influence of Salinity and Water Content on Soil Microorganisms. Int. Soil Water Conserv. Res. 2015, 3, 316–323. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.E.; Hocking, P.J.; Simpson, R.J.; George, T.S.; Richardson, A.E.; Hocking, P.J.; Simpson, R.J.; George, T.S. Plant Mechanisms to Optimise Access to Soil Phosphorus. Crop Pasture Sci. 2009, 60, 124–143. [Google Scholar] [CrossRef]
- Mukherjee, S.; Sen, S.K. Exploration of Novel Rhizospheric Yeast Isolate as Fertilizing Soil Inoculant for Improvement of Maize Cultivation. J. Sci. Food Agric. 2015, 95, 1491–1499. [Google Scholar] [CrossRef]
- Nutaratat, P.; Srisuk, N.; Arunrattiyakorn, P.; Limtong, S. Plant Growth-Promoting Traits of Epiphytic and Endophytic Yeasts Isolated from Rice and Sugar Cane Leaves in Thailand. Fungal Biol. 2014, 118, 683–694. [Google Scholar] [CrossRef]
- Fernandez-San Millan, A.; Farran, I.; Larraya, L.; Ancin, M.; Arregui, L.M.; Veramendi, J. Plant Growth-Promoting Traits of Yeasts Isolated from Spanish Vineyards: Benefits for Seedling Development. Microbiol. Res. 2020, 237, 126480. [Google Scholar] [CrossRef]
- Riggs, P.J.; Chelius, M.K.; Iniguez, A.L.; Kaeppler, S.M.; Triplett, E.W. Enhanced Maize Productivity by Inoculation with Diazotrophic Bacteria. Funct. Plant Biol. 2001, 28, 829–836. [Google Scholar] [CrossRef]
- Tairo, E.V.; Ndakidemi, P.A. Possible Benefits of Rhizobial Inoculation and Phosphorus Supplementation on Nutrition, Growth and Economic Sustainability in Grain Legumes. Am. J. Res. Commun. 2013, 1, 532–556. [Google Scholar]
- Oberson, A.; Frossard, E.; Bühlmann, C.; Mayer, J.; Mäder, P.; Lüscher, A. Nitrogen Fixation and Transfer in Grass-Clover Leys under Organic and Conventional Cropping Systems. Plant Soil 2013, 371, 237–255. [Google Scholar] [CrossRef] [Green Version]
- Kolodiazhnyi, O.I. Phosphorus Compounds of Natural Origin: Prebiotic, Stereochemistry, Application. Symmetry 2021, 13, 889. [Google Scholar] [CrossRef]
- Etesami, H.; Maheshwari, D.K. Use of Plant Growth Promoting Rhizobacteria (PGPRs) with Multiple Plant Growth Promoting Traits in Stress Agriculture: Action Mechanisms and Future Prospects. Ecotoxicol. Environ. Saf. 2018, 156, 225–246. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H. Enhanced Phosphorus Fertilizer Use Efficiency with Microorganisms. In Nutrient Dynamics for Sustainable Crop Production; Meena, R.S., Ed.; Springer: Singapore, 2020; pp. 215–245. ISBN 9789811386602. [Google Scholar]
- Timofeeva, A.; Galyamova, M.; Sedykh, S. Prospects for Using Phosphate-Solubilizing Microorganisms as Natural Fertilizers in Agriculture. Plants 2022, 11, 2119. [Google Scholar] [CrossRef] [PubMed]
- Goswami, D.; Pithwa, S.; Dhandhukia, P.; Thakker, J.N. Delineating Kocuria Turfanensis 2M4 as a Credible PGPR: A Novel IAA-Producing Bacteria Isolated from Saline Desert. J. Plant Interact. 2014, 9, 566–576. [Google Scholar] [CrossRef]
- Haas, D.; Défago, G. Biological Control of Soil-Borne Pathogens by Fluorescent Pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef]
- Saha, M.; Sarkar, S.; Sarkar, B.; Sharma, B.K.; Bhattacharjee, S.; Tribedi, P. Microbial Siderophores and Their Potential Applications: A Review. Environ. Sci. Pollut. Res. Int. 2016, 23, 3984–3999. [Google Scholar] [CrossRef]
- Rajkumar, M.; Ae, N.; Prasad, M.N.V.; Freitas, H. Potential of Siderophore-Producing Bacteria for Improving Heavy Metal Phytoextraction. Trends Biotechnol. 2010, 28, 142–149. [Google Scholar] [CrossRef]
- Latour, X.; Delorme, S.; Mirleau, P.; Lemanceau, P. Identification of Traits Implicated in the Rhizosphere Competence of Fluorescent Pseudomonads: Description of a Strategy Based on Population and Model Strain Studies. Agronomie 2003, 23, 397–405. [Google Scholar] [CrossRef]
- Gong, Q.; Li, Z.; Wang, L.; Dai, T.; Kang, Q.; Niu, D. Exogenous of Indole-3-Acetic Acid Application Alleviates Copper Toxicity in Spinach Seedlings by Enhancing Antioxidant Systems and Nitrogen Metabolism. Toxics 2019, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Zou, H.; Wang, B.; Yuan, F. Progress and Applications of Plant Growth-Promoting Bacteria in Salt Tolerance of Crops. Int. J. Mol. Sci. 2022, 23, 7036. [Google Scholar] [CrossRef] [PubMed]
- Jha, C.K.; Saraf, M. Plant Growth Promoting Rhizobacteria (PGPR): A Review. J. Agric. Res. Dev. 2015, 5, 108–119. [Google Scholar] [CrossRef]
- Ahmad, F.; Ahmad, I.; Khan, M. Indole Acetic Acid Production by the Indigenous Isolates of Azotobacter and Fluorescent Pseudomonas in the Presence and Absence of Tryptophan. Turk. J. Biol. 2005, 29, 29–34. [Google Scholar]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-Acetic Acid in Microbial and Microorganism-Plant Signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [Green Version]
- Mohite, B. Isolation and Characterization of Indole Acetic Acid (IAA) Producing Bacteria from Rhizospheric Soil and Its Effect on Plant Growth. J. Soil Sci. Plant Nutr. 2013, 13, 638–649. [Google Scholar] [CrossRef]
- Defez, R.; Andreozzi, A.; Romano, S.; Pocsfalvi, G.; Fiume, I.; Esposito, R.; Angelini, C.; Bianco, C. Bacterial IAA-Delivery into Medicago Root Nodules Triggers a Balanced Stimulation of C and N Metabolism Leading to a Biomass Increase. Microorganisms 2019, 7, 403. [Google Scholar] [CrossRef] [Green Version]
- Albacete, A.; Ghanem, M.E.; Martínez-Andújar, C.; Acosta, M.; Sánchez-Bravo, J.; Martínez, V.; Lutts, S.; Dodd, I.C.; Pérez-Alfocea, F. Hormonal Changes in Relation to Biomass Partitioning and Shoot Growth Impairment in Salinized Tomato (Solanum lycopersicum L.) Plants. J. Exp. Bot. 2008, 59, 4119–4131. [Google Scholar] [CrossRef]
- Ramos Solano, B.; Barriuso, J.; Gutiérrez Mañero, F.J. Physiological and Molecular Mechanisms of Plant Growth Promoting Rhizobacteria (PGPR). In Plant-Bacteria Interactions; John Wiley & Sons, Ltd.: New York, NY, USA, 2008; pp. 41–54. ISBN 978-3-527-62198-9. [Google Scholar]
- Etesami, H.; Alikhani, H.A. Co-Inoculation with Endophytic and Rhizosphere Bacteria Allows Reduced Application Rates of N-Fertilizer for Rice Plant. Rhizosphere 2016, 2, 5–12. [Google Scholar] [CrossRef]
- Kerbab, S.; Silini, A.; Chenari Bouket, A.; Cherif-Silini, H.; Eshelli, M.; El Houda Rabhi, N.; Belbahri, L. Mitigation of NaCl Stress in Wheat by Rhizosphere Engineering Using Salt Habitat Adapted PGPR Halotolerant Bacteria. Appl. Sci. 2021, 11, 1034. [Google Scholar] [CrossRef]
- Amini Hajiabadi, A.; Mosleh Arani, A.; Ghasemi, S.; Rad, M.H.; Etesami, H.; Shabazi Manshadi, S.; Dolati, A. Mining the Rhizosphere of Halophytic Rangeland Plants for Halotolerant Bacteria to Improve Growth and Yield of Salinity-Stressed Wheat. Plant Physiol. Biochem. 2021, 163, 139–153. [Google Scholar] [CrossRef]
- Pliego, C.; Cazorla, F.M.; González-Sánchez, M.A.; Pérez-Jiménez, R.M.; de Vicente, A.; Ramos, C. Selection for Biocontrol Bacteria Antagonistic toward Rosellinia Necatrix by Enrichment of Competitive Avocado Root Tip Colonizers. Res. Microbiol. 2007, 158, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Fatima, T.; Arora, N.K. Pseudomonas Entomophila PE3 and Its Exopolysaccharides as Biostimulants for Enhancing Growth, Yield and Tolerance Responses of Sunflower under Saline Conditions. Microbiol. Res. 2021, 244, 126671. [Google Scholar] [CrossRef]
- Li, M.; Guo, R.; Yu, F.; Chen, X.; Zhao, H.; Li, H.; Wu, J. Indole-3-Acetic Acid Biosynthesis Pathways in the Plant-Beneficial Bacterium Arthrobacter Pascens ZZ21. Int. J. Mol. Sci. 2018, 19, 443. [Google Scholar] [CrossRef] [PubMed]
- Ahemad, M.; Khan, M.S. Comparative Toxicity of Selected Insecticides to Pea Plants and Growth Promotion in Response to Insecticide-Tolerant and Plant Growth Promoting Rhizobium Leguminosarum. Crop Prot. 2010, 29, 325–329. [Google Scholar] [CrossRef]
- Yaish, M.W.; Antony, I.; Glick, B.R. Isolation and Characterization of Endophytic Plant Growth-Promoting Bacteria from Date Palm Tree (Phoenix dactylifera L.) and Their Potential Role in Salinity Tolerance. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2015, 107, 1519–1532. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Rodríguez, A.; Heydrich-Pérez, M.; Acebo-Guerrero, Y.; Velazquez-del Valle, M.G.; Hernández-Lauzardo, A.N. Antagonistic Activity of Cuban Native Rhizobacteria against Fusarium verticillioides (Sacc.) Nirenb. in Maize (Zea mays L.). Appl. Soil Ecol. 2008, 39, 180–186. [Google Scholar] [CrossRef]
- Shaharoona, B.; Naveed, M.; Arshad, M.; Zahir, Z.A. Fertilizer-Dependent Efficiency of Pseudomonads for Improving Growth, Yield, and Nutrient Use Efficiency of Wheat (Triticum aestivum L.). Appl. Microbiol. Biotechnol. 2008, 79, 147–155. [Google Scholar] [CrossRef]
- Barbaccia, P.; Gaglio, R.; Dazzi, C.; Miceli, C.; Bella, P.; Lo Papa, G.; Settanni, L. Plant Growth-Promoting Activities of Bacteria Isolated from an Anthropogenic Soil Located in Agrigento Province. Microorganisms 2022, 10, 2167. [Google Scholar] [CrossRef]
- Wani, P.A.; Khan, M.S. Bacillus Species Enhance Growth Parameters of Chickpea (Cicer arietinum L.) in Chromium Stressed Soils. Food Chem. Toxicol. 2010, 48, 3262–3267. [Google Scholar] [CrossRef]
- Jin, T.; Ren, J.; Li, Y.; Bai, B.; Liu, R.; Wang, Y. Plant Growth-Promoting Effect and Genomic Analysis of the P. Putida LWPZF Isolated from C. Japonicum Rhizosphere. AMB Express 2022, 12, 101. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, P.N.; Jha, D.K. Plant Growth-Promoting Rhizobacteria (PGPR): Emergence in Agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Komatsu, S. The Role of Phytohormones in Plant Response to Flooding. Int. J. Mol. Sci. 2022, 23, 6383. [Google Scholar] [CrossRef]
- Glick, B.R. Bacteria with ACC Deaminase Can Promote Plant Growth and Help to Feed the World. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Glick, B.R.; Cheng, Z.; Czarny, J.; Duan, J. Promotion of Plant Growth by ACC Deaminase-Producing Soil Bacteria. Eur. J. Plant Pathol. 2007, 119, 329–339. [Google Scholar] [CrossRef]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M.I.R. Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other Phytohormones. Front. Plant Sci. 2017, 8, 475. [Google Scholar] [CrossRef] [Green Version]
- Riyazuddin, R.; Verma, R.; Singh, K.; Nisha, N.; Keisham, M.; Bhati, K.K.; Kim, S.T.; Gupta, R. Ethylene: A Master Regulator of Salinity Stress Tolerance in Plants. Biomolecules 2020, 10, 959. [Google Scholar] [CrossRef]
- Carmen, B.; Roberto, D. Soil Bacteria Support and Protect Plants Against Abiotic Stresses; IntechOpen: Rijeka, Croatia, 2011; ISBN 978-953-307-394-1. [Google Scholar]
- Pérez-López, U.; Robredo, A.; Lacuesta, M.; Mena-Petite, A.; Muñoz-Rueda, A. Elevated CO2 Reduces Stomatal and Metabolic Limitations on Photosynthesis Caused by Salinity in Hordeum Vulgare. Photosynth. Res. 2012, 111, 269–283. [Google Scholar] [CrossRef]
- Munns, R. Comparative Physiology of Salt and Water Stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Dodd, I.C.; Pérez-Alfocea, F. Microbial Amelioration of Crop Salinity Stress. J. Exp. Bot. 2012, 63, 3415–3428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, D.; Lade, H. Plant-Growth-Promoting Rhizobacteria to Improve Crop Growth in Saline Soils: A Review. Agron. Sustain. Dev. 2014, 34, 737–752. [Google Scholar] [CrossRef]
- Morillon, R.; Chrispeels, M.J. The Role of ABA and the Transpiration Stream in the Regulation of the Osmotic Water Permeability of Leaf Cells. Proc. Natl. Acad. Sci. USA 2001, 98, 14138–14143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sleator, R.D.; Hill, C. Bacterial Osmoadaptation: The Role of Osmolytes in Bacterial Stress and Virulence. FEMS Microbiol. Rev. 2002, 26, 49–71. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, A.; Shahbaz, M.; Asadullah; Imran, A.; Marghoob, M.U.; Imtiaz, M.; Mubeen, F. Potential of Salt Tolerant PGPR in Growth and Yield Augmentation of Wheat (Triticum aestivum L.) Under Saline Conditions. Front. Microbiol. 2020, 11, 2019. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Asthir, B. Proline: A Key Player in Plant Abiotic Stress Tolerance. Biol. Plant 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, Salt, and Temperature Stress-Induced Metabolic Rearrangements and Regulatory Networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [Green Version]
- Kunst, F.; Ogasawara, N.; Moszer, I.; Albertini, A.M.; Alloni, G.; Azevedo, V.; Bertero, M.G.; Bessières, P.; Bolotin, A.; Borchert, S.; et al. The Complete Genome Sequence of the Gram-Positive Bacterium Bacillus Subtilis. Nature 1997, 390, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Meena, M.; Divyanshu, K.; Kumar, S.; Swapnil, P.; Zehra, A.; Shukla, V.; Yadav, M.; Upadhyay, R.S. Regulation of L-Proline Biosynthesis, Signal Transduction, Transport, Accumulation and Its Vital Role in Plants during Variable Environmental Conditions. Heliyon 2019, 5, e02952. [Google Scholar] [CrossRef] [Green Version]
- Kohl, D.H.; Straub, P.F.; Shearer, G. Does Proline Play a Special Role in Bacteroid Metabolism? Plant Cell Environ. 1994, 17, 1257–1262. [Google Scholar] [CrossRef]
- Yasin, N.A.; Akram, W.; Khan, W.U.; Ahmad, S.R.; Ahmad, A.; Ali, A. Halotolerant Plant-Growth Promoting Rhizobacteria Modulate Gene Expression and Osmolyte Production to Improve Salinity Tolerance and Growth in Capsicum annum L. Environ. Sci. Pollut. Res. Int. 2018, 25, 23236–23250. [Google Scholar] [CrossRef] [PubMed]
- Sukweenadhi, J.; Balusamy, S.R.; Kim, Y.-J.; Lee, C.H.; Kim, Y.-J.; Koh, S.C.; Yang, D.C. A Growth-Promoting Bacteria, Paenibacillus Yonginensis DCY84T Enhanced Salt Stress Tolerance by Activating Defense-Related Systems in Panax Ginseng. Front. Plant Sci. 2018, 9, 813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wutipraditkul, N.; Wongwean, P.; Buaboocha, T. Alleviation of Salt-Induced Oxidative Stress in Rice Seedlings by Proline and/or Glycinebetaine. Biol. Plant 2015, 59, 547–553. [Google Scholar] [CrossRef]
- Hoque, M.A.; Banu, M.N.A.; Okuma, E.; Amako, K.; Nakamura, Y.; Shimoishi, Y.; Murata, Y. Exogenous Proline and Glycinebetaine Increase NaCl-Induced Ascorbate-Glutathione Cycle Enzyme Activities, and Proline Improves Salt Tolerance More than Glycinebetaine in Tobacco Bright Yellow-2 Suspension-Cultured Cells. J. Plant Physiol. 2007, 164, 1457–1468. [Google Scholar] [CrossRef] [PubMed]
- Malekzadeh, P. Influence of Exogenous Application of Glycinebetaine on Antioxidative System and Growth of Salt-Stressed Soybean Seedlings (Glycine max L.). Physiol. Mol. Biol. Plants 2015, 21, 225–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kappes, R.M.; Kempf, B.; Kneip, S.; Boch, J.; Gade, J.; Meier-Wagner, J.; Bremer, E. Two Evolutionarily Closely Related ABC Transporters Mediate the Uptake of Choline for Synthesis of the Osmoprotectant Glycine Betaine in Bacillus Subtilis. Mol. Microbiol. 1999, 32, 203–216. [Google Scholar] [CrossRef]
- Salazar, B.; Ortiz, A.; Keswani, C.; Minkina, T.; Mandzhieva, S.; Pratap Singh, S.; Rekadwad, B.; Borriss, R.; Jain, A.; Singh, H.B.; et al. Bacillus spp. as Bio-Factories for Antifungal Secondary Metabolites: Innovation Beyond Whole Organism Formulations. Microb. Ecol. 2022. [Google Scholar] [CrossRef]
- Giri, J. Glycinebetaine and Abiotic Stress Tolerance in Plants. Plant Signal. Behav. 2011, 6, 1746–1751. [Google Scholar] [CrossRef]
- Mukhtar, S.; Zareen, M.; Khaliq, Z.; Mehnaz, S.; Malik, K.A. Phylogenetic Analysis of Halophyte-Associated Rhizobacteria and Effect of Halotolerant and Halophilic Phosphate-Solubilizing Biofertilizers on Maize Growth under Salinity Stress Conditions. J. Appl. Microbiol. 2020, 128, 556–573. [Google Scholar] [CrossRef]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Al-Huqail, A.A.; Shah, M.A. Induction of Osmoregulation and Modulation of Salt Stress in Acacia Gerrardii Benth. by Arbuscular Mycorrhizal Fungi and Bacillus Subtilis (BERA 71). Biomed. Res. Int. 2016, 2016, 6294098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masouleh, S.S.S.; Aldine, N.J.; Sassine, Y.N. The Role of Organic Solutes in the Osmotic Adjustment of Chilling-Stressed Plants (Vegetable, Ornamental and Crop Plants). Ornam. Hortic. 2020, 25, 434–442. [Google Scholar] [CrossRef]
- Moghaieb, R.E.A.; Tanaka, N.; Saneoka, H.; Murooka, Y.; Ono, H.; Morikawa, H.; Nakamura, A.; Nguyen, N.T.; Suwa, R.; Fujita, K. Characterization of Salt Tolerance in Ectoine-Transformed Tobacco Plants (Nicotiana tabaccum): Photosynthesis, Osmotic Adjustment, and Nitrogen Partitioning. Plant Cell Environ. 2006, 29, 173–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grammann, K.; Volke, A.; Kunte, H.J. New Type of Osmoregulated Solute Transporter Identified in Halophilic Members of the Bacteria Domain: TRAP Transporter TeaABC Mediates Uptake of Ectoine and Hydroxyectoine in Halomonas Elongata DSM 2581T. J. Bacteriol. 2002, 184, 3078–3085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsakhawy, T.A.; Nashwa, A.H.F.; Ghazi, A.A. The Potential Use of Ectoine Produced by a Moderately Halophilic Bacteria Chromohalobacter Salexigens KT989776 for Enhancing Germination and Primary Seedling of Flax “Linum usitatissimum L.” under Salinity Conditions. Biotechnol. J. Int. 2019, 23, 1–12. [Google Scholar] [CrossRef]
- Chandra, G.; Chater, K.F.; Bornemann, S. Unexpected and Widespread Connections between Bacterial Glycogen and Trehalose Metabolism. Microbiology 2011, 157, 1565–1572. [Google Scholar] [CrossRef] [Green Version]
- Vílchez, J.I.; García-Fontana, C.; Román-Naranjo, D.; González-López, J.; Manzanera, M. Plant Drought Tolerance Enhancement by Trehalose Production of Desiccation-Tolerant Microorganisms. Front. Microbiol. 2016, 7, 1577. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Salazar, J.; Suárez, R.; Caballero-Mellado, J.; Iturriaga, G. Trehalose Accumulation in Azospirillum Brasilense Improves Drought Tolerance and Biomass in Maize Plants. FEMS Microbiol. Lett. 2009, 296, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Müller, K.M.; Charles, T.C.; Vesely, S.; Glick, B.R. 1-Aminocyclopropane-1-Carboxylate (ACC) Deaminase Genes in Rhizobia from Southern Saskatchewan. Microb. Ecol. 2009, 57, 423–436. [Google Scholar] [CrossRef]
- Suárez, R.; Wong, A.; Ramírez, M.; Barraza, A.; Orozco, M.D.C.; Cevallos, M.A.; Lara, M.; Hernández, G.; Iturriaga, G. Improvement of Drought Tolerance and Grain Yield in Common Bean by Overexpressing Trehalose-6-Phosphate Synthase in Rhizobia. Mol. Plant Microbe Interact. 2008, 21, 958–966. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.C.; Wingender, J. Relevance of Microbial Extracellular Polymeric Substances (EPSs)--Part I: Structural and Ecological Aspects. Water Sci. Technol. 2001, 43, 1–8. [Google Scholar] [CrossRef]
- Liu, X.; Luo, Y.; Li, Z.; Wang, J.; Wei, G. Role of Exopolysaccharide in Salt Stress Resistance and Cell Motility of Mesorhizobium Alhagi CCNWXJ12-2T. Appl. Microbiol. Biotechnol. 2017, 101, 2967–2978. [Google Scholar] [CrossRef]
- Qin, Y.; Druzhinina, I.S.; Pan, X.; Yuan, Z. Microbially Mediated Plant Salt Tolerance and Microbiome-Based Solutions for Saline Agriculture. Biotechnol. Adv. 2016, 34, 1245–1259. [Google Scholar] [CrossRef] [PubMed]
- Ruppel, S.; Franken, P.; Witzel, K. Properties of the Halophyte Microbiome and Their Implications for Plant Salt Tolerance. Funct. Plant Biol. 2013, 40, 940–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vuyst, L.; Degeest, B. Heteropolysaccharides from Lactic Acid Bacteria. FEMS Microbiol. Rev. 1999, 23, 153–177. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, M.; Wani, S.P. Rhizobacterial-Plant Interactions: Strategies Ensuring Plant Growth Promotion under Drought and Salinity Stress. Agric. Ecosyst. Environ. 2016, 231, 68–78. [Google Scholar] [CrossRef]
- Meneses, C.; Gonçalves, T.; Alquéres, S.; Rouws, L.; Serrato, R.; Vidal, M.; Baldani, J.I. Gluconacetobacter Diazotrophicus Exopolysaccharide Protects Bacterial Cells against Oxidative Stress in Vitro and during Rice Plant Colonization. Plant Soil 2017, 416, 133–147. [Google Scholar] [CrossRef]
- Xiong, Y.-W.; Ju, X.-Y.; Li, X.-W.; Gong, Y.; Xu, M.-J.; Zhang, C.-M.; Yuan, B.; Lv, Z.-P.; Qin, S. Fermentation Conditions Optimization, Purification, and Antioxidant Activity of Exopolysaccharides Obtained from the Plant Growth-Promoting Endophytic Actinobacterium Glutamicibacter Halophytocola KLBMP 5180. Int. J. Biol. Macromol. 2020, 153, 1176–1185. [Google Scholar] [CrossRef]
- Gupta, G.; Parihar, S.; Ahirwar, N.; Snehi, S.K.; Singh, V. Plant Growth Promoting Rhizobacteria (PGPR): Current and Future Prospects for Development of Sustainable Agriculture. Microb. Biochem. Technol. 2015, 7, 096–102. [Google Scholar]
- Xiong, Y.-W.; Gong, Y.; Li, X.-W.; Chen, P.; Ju, X.-Y.; Zhang, C.-M.; Yuan, B.; Lv, Z.-P.; Xing, K.; Qin, S. Enhancement of Growth and Salt Tolerance of Tomato Seedlings by a Natural Halotolerant Actinobacterium Glutamicibacter Halophytocola KLBMP 5180 Isolated from a Coastal Halophyte. Plant Soil 2019, 445, 307–322. [Google Scholar] [CrossRef]
- Mishra, J.; Arora, N.K. Secondary Metabolites of Fluorescent Pseudomonads in Biocontrol of Phytopathogens for Sustainable Agriculture. Appl. Soil Ecol. 2018, 125, 35–45. [Google Scholar] [CrossRef]
- Tewari, S.; Arora, N.K. Role of Salicylic Acid from Pseudomonas Aeruginosa PF23EPS+ in Growth Promotion of Sunflower in Saline Soils Infested with Phytopathogen Macrophomina Phaseolina. Environ. Sustain. 2018, 1, 49–59. [Google Scholar] [CrossRef]
- Ashraf, M.; Hasnain, S.; Berge, O.; Mahmood, T. Inoculating Wheat Seedlings with Exopolysaccharide-Producing Bacteria Restricts Sodium Uptake and Stimulates Plant Growth under Salt Stress. Biol. Fertil. Soils 2004, 40, 157–162. [Google Scholar] [CrossRef]
- Mahmood, S.; Daur, I.; Al-Solaimani, S.G.; Ahmad, S.; Madkour, M.H.; Yasir, M.; Hirt, H.; Ali, S.; Ali, Z. Plant Growth Promoting Rhizobacteria and Silicon Synergistically Enhance Salinity Tolerance of Mung Bean. Front. Plant Sci. 2016, 7, 876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Lei, P.; Wang, Q.; Ma, J.; Zhan, Y.; Jiang, K.; Xu, Z.; Xu, H. The Endophyte Pantoea Alhagi NX-11 Alleviates Salt Stress Damage to Rice Seedlings by Secreting Exopolysaccharides. Front. Microbiol. 2020, 10, 3112. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Tapias, D.; Moreno-Galván, A.; Pardo-Díaz, S.; Obando, M.; Rivera, D.; Bonilla, R. Effect of Inoculation with Plant Growth-Promoting Bacteria (PGPB) on Amelioration of Saline Stress in Maize (Zea mays). Appl. Soil Ecol. 2012, 61, 264–272. [Google Scholar] [CrossRef]
- Fatima, T.; Mishra, I.; Verma, R.; Arora, N.K. Mechanisms of Halotolerant Plant Growth Promoting Alcaligenes Sp. Involved in Salt Tolerance and Enhancement of the Growth of Rice under Salinity Stress. 3 Biotech 2020, 10, 361. [Google Scholar] [CrossRef] [PubMed]
- Mhlongo, M.I.; Piater, L.A.; Dubery, I.A. Profiling of Volatile Organic Compounds from Four Plant Growth-Promoting Rhizobacteria by SPME–GC–MS: A Metabolomics Study. Metabolites 2022, 12, 763. [Google Scholar] [CrossRef]
- Vespermann, A.; Kai, M.; Piechulla, B. Rhizobacterial Volatiles Affect the Growth of Fungi and Arabidopsis thaliana. Appl. Environ. Microbiol. 2007, 73, 5639–5641. [Google Scholar] [CrossRef] [Green Version]
- Kanchiswamy, C.N.; Malnoy, M.; Maffei, M.E. Chemical Diversity of Microbial Volatiles and Their Potential for Plant Growth and Productivity. Front. Plant Sci. 2015, 6, 151. [Google Scholar] [CrossRef] [Green Version]
- Tirranen, L.S.; Gitelson, I.I. The Role of Volatile Metabolites in Microbial Communities of the LSS Higher Plant Link. Adv. Space Res. 2006, 38, 1227–1232. [Google Scholar] [CrossRef]
- Tahir, H.A.S.; Gu, Q.; Wu, H.; Raza, W.; Hanif, A.; Wu, L.; Colman, M.V.; Gao, X. Plant Growth Promotion by Volatile Organic Compounds Produced by Bacillus Subtilis SYST2. Front. Microbiol. 2017, 8, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meldau, D.G.; Meldau, S.; Hoang, L.H.; Underberg, S.; Wünsche, H.; Baldwin, I.T. Dimethyl Disulfide Produced by the Naturally Associated Bacterium Bacillus Sp B55 Promotes Nicotiana Attenuata Growth by Enhancing Sulfur Nutrition. Plant Cell 2013, 25, 2731–2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, S.M.; Kang, B.R.; Han, S.H.; Anderson, A.J.; Park, J.-Y.; Lee, Y.-H.; Cho, B.H.; Yang, K.-Y.; Ryu, C.-M.; Kim, Y.C. 2R,3R-Butanediol, a Bacterial Volatile Produced by Pseudomonas Chlororaphis O6, Is Involved in Induction of Systemic Tolerance to Drought in Arabidopsis thaliana. Mol. Plant Microbe Interact. 2008, 21, 1067–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Kim, M.-S.; Krishnamachari, V.; Payton, P.; Sun, Y.; Grimson, M.; Farag, M.A.; Ryu, C.-M.; Allen, R.; Melo, I.S.; et al. Rhizobacterial Volatile Emissions Regulate Auxin Homeostasis and Cell Expansion in Arabidopsis. Planta 2007, 226, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Li, H.F.; Imai, T.; Ukita, M.; Sekine, M.; Higuchi, T. Compost Stability Assessment Using a Secondary Metabolite: Geosmin. Environ. Technol. 2004, 25, 1305–1312. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Luna, F.M.; López-Bucio, J.; Altamirano-Hernández, J.; Valencia-Cantero, E.; de la Cruz, H.R.; Macías-Rodríguez, L. Plant Growth-Promoting Rhizobacteria Modulate Root-System Architecture in Arabidopsis thaliana through Volatile Organic Compound Emission. Symbiosis 2010, 51, 75–83. [Google Scholar] [CrossRef]
- Cordovez, V.; Schop, S.; Hordijk, K.; Dupré de Boulois, H.; Coppens, F.; Hanssen, I.; Raaijmakers, J.M.; Carrión, V.J. Priming of Plant Growth Promotion by Volatiles of Root-Associated Microbacterium spp. Appl. Environ. Microbiol. 2018, 84, e01865-18. [Google Scholar] [CrossRef] [Green Version]
- Ledger, T.; Rojas, S.; Timmermann, T.; Pinedo, I.; Poupin, M.J.; Garrido, T.; Richter, P.; Tamayo, J.; Donoso, R. Volatile-Mediated Effects Predominate in Paraburkholderia Phytofirmans Growth Promotion and Salt Stress Tolerance of Arabidopsis thaliana. Front. Microbiol. 2016, 7, 1838. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, D.; Lee, Y.H. A Cocktail of Volatile Compounds Emitted from Alcaligenes Faecalis JBCS1294 Induces Salt Tolerance in Arabidopsis thaliana by Modulating Hormonal Pathways and Ion Transporters. J. Plant Physiol. 2017, 214, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Meena, K.K.; Sorty, A.M.; Bitla, U.M.; Choudhary, K.; Gupta, P.; Pareek, A.; Singh, D.P.; Prabha, R.; Sahu, P.K.; Gupta, V.K.; et al. Abiotic Stress Responses and Microbe-Mediated Mitigation in Plants: The Omics Strategies. Front. Plant Sci. 2017, 8, 172. [Google Scholar] [CrossRef] [Green Version]
- Siebers, N.; Kruse, J. Short-Term Impacts of Forest Clear-Cut on Soil Structure and Consequences for Organic Matter Composition and Nutrient Speciation: A Case Study. PLoS ONE 2019, 14, e0220476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, P.A.; Rothballer, M.; Chowdhury, S.P.; Nussbaumer, T.; Gutjahr, C.; Falter-Braun, P. Systems Biology of Plant-Microbiome Interactions. Mol. Plant 2019, 12, 804–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arif, F.; Ghoul, M. Halotolerance of Indigenous Fluorescent Pseudomonads in the Presence of Natural Osmoprotectants. Annu. Res. Rev. Biol. 2018, 24, 1–11. [Google Scholar] [CrossRef]
- Ansari, F.A.; Ahmad, I.; Pichtel, J. Growth stimulation and alleviation of salinity stress to wheat by the biofilm forming Bacillus pumilus strain FAB10. Appl. Soil Ecol. 2019, 143, 45–54. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | HT-PGPR Species | Gene/s Involved | Mechanism to Mitigate Stress | Effect Observed | References |
|---|---|---|---|---|---|
| Arabidopsis thaliana L. | Bacillus oryzicola YC7007 | RD22, KIN1, RD29B, RD20, RD22, and ERD1 | Stem and the root of the seedlings released stress-related genes | Enhanced plant tolerance to salt stress | [35] |
| Pseudomonas putida PS01 | APX2 and GLYI7 | APX2 and GLYI7 genes were downregulated | ABA signalling, jasmonic acid production route, ROS scavenging, detoxification | [36] | |
| Pseudomonas knackmussii MLR6 | NHX1, HKT1, SOS2, SOS3, SAG13, and PR1 | Enhanced stomatal conductance, transpiration rate, chlorophyll, and carotenoid levels | Reduced electrolyte leakage and priming ROS accumulation increasing cell membrane stability | [37] | |
| Bacillus amyloliquefaciens SQR9 | NHX1 and NHX7 | Involved in reducing GSH biosynthesis | Reduced ion toxicity by sequestering Na+ into vacuoles and releasing Na+ from the cell | [38] | |
| Burkholderia phytofirmans PsJN | Upregulation of RD29A and GLYI7, and downregulation of LOX2 | Enhancement of proline and transcription of genes related to abscisic acid signalling and downregulated gene Lipoxygenase 2 | Abscisic acid signalling, ROS reduction, detoxifying, jasmonate synthesis, and ion transport | [39] | |
| Paenibacillus yonginensis DCY84T | AtRSA1, AtVQ9 and AtWRKY8 | Upregulated salt-stress genes | Promoted more resistance to salinity, drought, and aluminium stresses | [40,41] | |
| Enterobacter sp. EJ01 | DREB2b, RD29A, RD29B, RAB18, P5CS1, P5CS2, MPK3, and MPK6 | Upregulated salt-stress genes | Promoted more resistance to salinity and enhanced plant growth | [42] | |
| Bacillus subtilis GB03 | HKT1 | Down- and upregulates HKT1 in roots and shoots, respectively | Decreased total plant Na+ accumulation | [23] | |
| Bacopa monneri L. | Dietzia natronolimnaea STR1 | SOS1, SOS4, TaST, TaNHX1, TaHAK, and TaHKT1 | Reduction in ABA-signalling, upregulated TaABARE and TaOPR1 | Abscisic acid signalling, ROS scavenging, antioxidant enzyme activity, enhanced ion transporter expression, high K+/Na+ ratio | [24] |
| Bacillus pumilus STR2, Exiguobacterium oxidotolerens STR36 | - | Mixture of plant growth-promoting traits under primary and secondary saline condition | Produced higher yield, high proline/lipid content peroxidation | [2] | |
| Cicer arietinum L. | Planococcus rifietoensis (RT4) and Halomonas variabilis (HT1) | - | Biofilm and exopolysaccharides production | Improved crop growth, soil aggregation, and soil fertility | [43] |
| Glycine max L. | Arthrobacter woluwensis AK1 | - | Reduced endogenous ABA and controlled antioxidant activity | Mitigated salinity stress and increased plant growth | [44] |
| Microbacterium oxydans, Arthrobacter woluwensis, Arthrobacter aurescens, Bacillus aryabhattai, and Bacillus megaterium | - | Increased production of IAA, GA, siderophores, and phosphate solubilisation | Increased antioxidant enzymes and K absorption; reduced Na+ in plant tissue; phytohormone | [45] | |
| Pseudomonas simiae AU | P5CS, PPO and HKT1 | Downregulated HKT1, LOX, PPO, and P5CS genes | Increased chlorophyll, phosphate solubilisation, IAA, and siderophores; decreased root surface in saline | [46] | |
| Pseudomonas sp. strain AK-1 | HTK1 | Improve K+/Na+ ratio and Exopolysaccharide production binds free Na+ from soil | Increased shoot/root length and decreased Na+/K+ ratio | [33] | |
| Pseudomonas simiae AU | VSP2 | Increase vegetative storage protein (VSP), gamma-glutamyl hydrolase (GGH), and RuBisCo proteins | Reduced Na, increased K and P in soybean seedling roots, high proline and chlorophyll content | [47] | |
| Helianthus annuus L. | Pseudomonas libanensis TR1 | - | ACC-deaminase and exopolysaccharide production | Ni and Na+ accumulation potential increased along with plant growth. | [48] |
| Pseudomonas spp. | - | Upregulating of ACC deaminase | Improved P and K contents, and K+/Na+ ratio in shoot | [49] | |
| Hordeum vulgare L. | Bacillus mojavensis, B. pumilus and Pseudomonas fluorescens | S1 and S3 | ACC deaminase, IAA, and proline production | Reduced plant Na concentration, stimulated root growth, improved water and nutrient absorption | [50] |
| B. aryabhattai MS3 | BZ8, SOS1, GIG, and NHX1 | Increased salt stress resistance and accumulation | Adaptation of plant under saline condition | [51] | |
| Bacillus amyloliquefaciens SN13 | DHN | Upregulated salt stress-responsive genes and protein-related genes | Lipid peroxidation and electrolyte leakage reduced; increased rice biomass, water content, proline, and total soluble sugar | [52] | |
| Bacillus megaterium ST2-1 | - | IAA production | Stimulated the growth of rice roots and dry biomass | [53] | |
| Pseudomonas pseudoalcaligenes ST1, Bacillus pumilus ST2 | EU440977 and FJ840535 | Accumulation of proline decrease with inoculation, antioxidative activity | Enhanced plant growth by ROS scavenging and higher accumulation of osmoprotectant | [54] | |
| Puccinellia tenuiflora L. | Bacillus subtilis (GB03) | - | Upregulated PtHKT1;5 and PtSOS1 genes, downregulated PtHKT2;1 | Na homeostasis modulation, exclusive K+ absorption | [55] |
| Solanum lycopersicum L. | Leclercia adecarboxylata MO1 | - | ACC deaminase and IAA production | Increased soluble sugars: organic glucose, sucrose, fructose, malic, amino acid, and proline | [56] |
| Sphingobacterium sp. BHU-AV3 | - | Reduction in ROS concentration in plant | Enhanced antioxidant activities and energy metabolism | [57] | |
| Enterobacter sp. EJ01 | DREB2b, RD29A, RD29B, and RAB18 | Downregulated P5CS1 and P5CS2, and upregulated MPK3 and MPK6 | Biosynthesis, defence pathway modulation, salt response | [42] | |
| Pseudomonas putida UW4 | Toc GTPase | Toc GTPase genes were upregulated and reduction in ACC deaminase | Increased shoot length and chlorophyll concentration | [58] | |
| Trifolium repens L. | Bacillus subtilis (GB03) | - | Reduced shoot and root Na+, improving K+/Na+ ratio | Decreased Na+, increased chlorophyll, leaf osmotic potential, cell membrane integrity | [59] |
| Triticum aestivum L. | Pseudomonas aeruginosa GI-1, and Burkholderia gladioli GI-6 | - | P solubilisation, catalase activity, IAA production, N assimilation, and siderophores production | Encouraged growth and yield and improve soil fertility | [60,61] |
| Arthrobacter nitroguajacolicus | - | Upregulated 152 genes whereas 5 genes were downregulated | Amplified ACC, IAA, siderophore, and phosphate solubility. ROS detoxification, Na+ homeostasis, abiotic stress | [62] | |
| Serratia marcescens CDP-13 | - | Increased salt tolerance in plant | ACC deaminase, phosphate solubilisation, siderophore, indole acetic acid, N fixation, and ammonia synthesis | [63] | |
| Pseodomonas sp and Enterobacter cloacae (R-10) | B-22 and S-49 | K and Zn solubilisation for identifying antifungal activity | Enhanced K+ uptake, dry matter of wheat | [64] | |
| Hallobacillus sp. SL3 Bacillus halodenitrificans PU62 | acdS | IAA production and siderophore production, phosphate solubilising, and siderophore production | Increased root elongation and dry weight | [65] | |
| Zea mays L. | Serratia liquefaciens KM4 | Upregulation of stress-related genes (APX, CAT, SOD, RBCS, RBCL, H+-PPase, HKT1, and NHX1) | Regulating redox potential and stress-related gene expression | Higher leaf gas exchange, osmoregulation, antioxidative defence mechanisms, and nutrient uptake boosted maize growth and biomass production | [66] |
| Azospirillum lipoferum, Azospillum sp., Azotobacter chroococcum, Azotobacter sp., and Bacillus sp. | - | Exopolysaccharide inoculation in the soil | Increased root and shoot dry weights, chlorophyll and carotenoids, restricted Na and Cl uptake, and increased shoot N, P, and K | [67] | |
| Abelmoschus esculentus L. | Enterobacter sp. UPMR18 | X55749 | ROS pathway upgradation and enhancement in antioxidant enzyme activities | Higher germination, growth, and chlorophyll improved salt tolerance | [68] |
| HT-PGPR | Crop | Role in Plants | Reference |
|---|---|---|---|
| Streptomyces fradiae NKZ-259 | Tomato | Increases root and shoot length | [1] |
| Stenotrophomonas maltophilia AVO63 | Avocado | Protection against white root rot | [106] |
| Pseudomonas entomophila PE3 | Sunflower | Antioxidant, hydroxyl-scavenging; reduction in Na+ accumulation | [107] |
| Arthrobacter pascens ZZ21 | Forest soil | Enhanced plant growth and remediate fluoranthene-contaminated soil | [108] |
| Rhizobium sp. MRP1 | Pea | Produce HCN, ammonia and EPS | [109] |
| Paenibacillus xylanexedens (PD-R6) and Enterobacter cloacae (PD-P6) | Date palm | Enhances nutrient uptake in roots and improves plant growth | [110] |
| Burkholderia cepacia 0057 | Maize | Enhancement of disease control | [111] |
| Bradyrhizobium japonicum | Wheat | Improved growth and yield | [112] |
| Bacillus tequilensis SI 319, Pseudomonas lini SI 287, P. frederiksbergensis SI 307, and Brevibacterium frigoritolerans SI 433 | Soil sample | Increased plant growth | [113] |
| Bacillus sp. PSB10 | Chickpea | Reduced uptake of chromium | [114] |
| Pseudomonas putida LWPZF | Katsura | Improved plant growth and contained anti-heavy-metal properties | [115] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, V.; Raghuvanshi, N.; Pandey, A.K.; Kumar, A.; Thoday-Kennedy, E.; Kant, S. Role of Halotolerant Plant Growth-Promoting Rhizobacteria in Mitigating Salinity Stress: Recent Advances and Possibilities. Agriculture 2023, 13, 168. https://doi.org/10.3390/agriculture13010168
Kumar V, Raghuvanshi N, Pandey AK, Kumar A, Thoday-Kennedy E, Kant S. Role of Halotolerant Plant Growth-Promoting Rhizobacteria in Mitigating Salinity Stress: Recent Advances and Possibilities. Agriculture. 2023; 13(1):168. https://doi.org/10.3390/agriculture13010168
Chicago/Turabian StyleKumar, Vikash, Nikhil Raghuvanshi, Abhay K. Pandey, Abhishek Kumar, Emily Thoday-Kennedy, and Surya Kant. 2023. "Role of Halotolerant Plant Growth-Promoting Rhizobacteria in Mitigating Salinity Stress: Recent Advances and Possibilities" Agriculture 13, no. 1: 168. https://doi.org/10.3390/agriculture13010168