
Exploring the Roles of Arbuscular Mycorrhizal Fungi in Plant–Iron Homeostasis

Department of Environmental and Energy Engineering, Yonsei University, Wonju 26493, Republic of Korea
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Agriculture 2023, 13(10), 1918; https://doi.org/10.3390/agriculture13101918
Submission received: 28 August 2023
/
Revised: 20 September 2023
/
Accepted: 29 September 2023
/
Published: 30 September 2023
(This article belongs to the Special Issue Mycorrhizal Symbiosis in Agricultural Production)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Arbuscular mycorrhizal fungi (AMF) form a vital symbiotic relationship with plants. Through their extensive hyphal networks, AMF extend the absorptive capacity of plant roots, thereby allowing plants to reach otherwise inaccessible micronutrient sources. Iron, a critical micronutrient involved in photosynthesis and other metabolic processes, often becomes inaccessible owing to its tendency to form insoluble complexes in soil. AMF symbiosis significantly ameliorates this challenge by enhancing iron uptake and homeostasis in plants, altering root architecture, and producing root exudates that improve iron solubility. Moreover, the interaction with diverse soil bacteria, particularly plant growth-promoting rhizobacteria, can potentiate the benefits of AMF symbiosis. Siderophores are low-molecular-weight chelators with iron-binding capacities produced by various microorganisms and plant roots. They play pivotal roles in regulating intracellular iron and have been identified in different mycorrhizal associations, including AMF. While molecular mechanisms behind AMF-mediated iron uptake have been partially explored, the intricate networks involving AMF, plants, siderophores, and other soil microbiota are largely unknown. This review focuses on the multifaceted roles of AMF in plant–iron homeostasis, interactions with soil bacteria, and the potential of siderophores in these processes, emphasizing the possibilities for harnessing these relationships for sustainable agriculture and enhancing plant productivity.
1. Introduction
Arbuscular mycorrhizal fungi (AMF) are soil-dwelling fungi that have been in a mutual symbiotic relationship with plants for millions of years. Approximately 85% of plant species in all terrestrial ecosystems have interdependent associations with AMF [1]. AMF belong to the phylum Glomeromycota and form arbuscules, vesicles, and hyphal coils within root cells during symbiotic development. They use their extensive hyphal network to acquire nutrients, such as phosphorus, nitrogen, potassium, and micronutrients, from the soil, deliver them to the host plant, and take up carbohydrates in return [1,2]. The complex network of fungal hyphae augments the absorptive area of roots, extending beyond the depletion zone around the root and effectively improving the accessibility of soil nutrients for plants. AMF can markedly enhance the uptake of micronutrients, including Cu, Zn, and Fe, which are often limited in bioavailability owing to their tendency to form insoluble complexes or their spatial distribution in soil [3,4]. Furthermore, AMF can mobilize and solubilize micronutrients by excreting organic acids and enzymes, thereby transforming these elements into more plant-accessible forms [5]. By regulating micronutrient concentrations in plant tissues, these fungi offer a buffering mechanism that mitigates the risks of deficiency or toxicity [6]. Consequently, AMF play a pivotal role in enhancing nutrient absorption by plants, transforming less accessible micronutrients into bioavailable forms, and safeguarding against nutrient imbalances.
Iron is a vital micronutrient, fundamental to several metabolic processes, including photosynthesis, respiration, and nitrogen fixation [7]. Even though the total amount of iron is abundant in soil, most iron is present mainly in the Fe3+ form, which is less soluble than Fe2+. The solubility of Fe3+ is largely dependent on pH and redox conditions [8]. Owing to the lower solubility of iron under high pH levels, iron becomes deficient for living organisms, posing a challenge to plant nutrition. In this context, AMF symbiosis significantly enhances iron uptake and homeostasis in plants. The association of plants with AMF alters root architecture, promoting the proliferation and elongation of roots and consequently increasing the root surface area [9,10]. This increase in root size and branching allows for a high contact area with soil, thereby facilitating excellent nutrient acquisition. Additionally, AMF can stimulate the production of root exudates that can chelate and solubilize iron, increasing its bioavailability for plant uptake [11]. The role of AMF extends beyond iron acquisition to encompass a broad role in plant stress tolerance. For instance, under iron-deficient conditions, plants in a symbiotic relationship with AMF exhibit enhanced stress tolerance mediated by improved nutrient homeostasis and triggering of stress-associated signaling pathways [12,13,14]. A noteworthy aspect of this symbiotic relationship is its association with the diversity of other soil bacteria. Bacterial communities in the rhizosphere and mycorrhizosphere can influence AMF–plant symbiosis and consequently affect iron uptake and homeostasis [15]. Certain bacteria, known as plant growth-promoting rhizobacteria, can enhance the efficiency of AMF symbiosis, potentially leading to improved iron acquisition and homeostasis.
Several pieces of evidence have provided insights into the molecular mechanism by which AMF positively alleviate iron deficiency in host plants; however, intricate networks of interactions involving AMF, plant, and other soil bacteria involved in iron homeostasis are still unknown. In particular, siderophores, which play the most important role in iron competition or cooperation in the environment, are important in regulating these networks. Siderophores are low-molecular-weight chelators produced by various microorganisms and plant roots and can produce high-affinity complexes with Fe3+ [16]. Siderophores facilitate iron acquisition and play a vital role in regulating intracellular iron levels by coordinating iron uptake, storage, and utilization [17,18]. Mycorrhizal fungi, representing the ericoid, orchidaceous, and ectomycorrhizal symbioses, release hydroxamate-type siderophores. Arbuscular mycorrhiza, which establishes a nutrient-exchange symbiosis, is identified by the presence of siderophores typical for the phylum Zygomycota. This distinction is made by detecting the rhizoferrin signature in arbuscular mycorrhizal plant tissue [19]. Understanding the role of siderophores in the intricate network of interactions involving AMF, plant, and other soil microbiota for iron homeostasis can open avenues for harnessing these symbiotic relationships for sustainable and resilient agriculture. In this review, we introduce the role of AMF in these interactions, their implications for plant–iron homeostasis, and potential ways to manipulate these interactions to improve plant health and productivity.
2. Do AMF Thrive in the Presence of Iron?
AMF do not thrive specifically in the presence of iron, but iron availability in soil plays a crucial role in the interaction between AMF and plants. Iron deficiency may reduce arbuscule formation and function, hindering the symbiotic relationship and overall nutrient uptake for both the fungi and host plant [20]. Moreover, iron is essential for the growth and development of fungal hyphae, promoting them to form robust hyphal networks and facilitating root colonization [2]. These behaviors indicate that iron availability in soil is vital for optimal AMF–plant interactions [21,22].
The efficient uptake of iron and its transfer to plants are major features of symbiosis. To acquire iron, all plant species must confront the challenges of iron insolubility and toxicity and evolve different mechanisms for iron acquisition [23]. Each fungus utilizes at least two of the four iron uptake mechanisms that are dependent on metalloreductases and involve Fe3+/Fe2+. These involve direct Fe2+ permeation, coupled ferroxidase/permease uptake, heme/hemin uptake, and siderophore-mediated iron uptake [24] (Figure 1). Fe2+ can be transported directly into fungal cells via divalent ion transporters such as Fet4p, which has been characterized in S. cerevisiae. The most abundant membrane reductase in fungi is Fre1p [25]. Initially, Fe3+ is reduced to Fe2+ by Fre1p, and reduced Fe2+ is then specifically taken up by a high-affinity transporter complex, i.e., Fet3p and Ftr1p. Some opportunistic pathogenic fungi, including Candida albicans and Histoplasma capsulatum, express a heme receptor Rbt5 gene required for virulence in organisms, which depends on this heme-iron uptake pathway [26,27]. Regardless of their ability to produce siderophores, fungi generally possess the siderophore transporters that enable them to uptake different types of siderophores such as bacterial coprogen or enterobactin [28]. The siderophore–iron complex is taken up by specific transporters, such as Sit1p, in the fungal cell membrane that is stored in hyphae for inclusion into spores, potentially affecting sporulation [29,30].
An in silico genome-wide analysis of Cu, Fe, and Zn transporters has identified 30 open reading frames in the whole genome of Rhizophagus irregularis, which potentially encode metal transporters [31]. It has been reported that R. irregularis contains a putative ferrireductase, RiFRE1, which encodes approximately 541 amino acids [32]. Ferroxidase pathways are then mediated by iron permease, namely RiFTR1 and RiFTR2 (two putative orthologs), which are, respectively, responsible for iron acquisition and iron homeostasis during iron deficiency. These iron permeases are expressed in germinated spores and mycorrhizal roots. Apart from this, R. irregularis consists of another ferroxidase pathway containing a single transmembrane domain and an extracellular multicopper oxidase (MCO) domain, which is responsible for the uptake of reduced Fe2+ via oxidation through a copper- and oxygen-dependent reaction. Recently, Tamayo et al. [33] observed nine genes that encode MCOs (RiMCO1–9), among which RiMCO1 and RiMCO3 might have possible roles in reductive and high-affinity iron uptake. Although the detailed mechanism of siderophore-mediated iron uptake by AMF remains to be elucidated, genome analysis has shown that AMF, including Gigaspora rosea, R. irregularis, Glomus cerebriforme, and Diversispora epigea, possess a non-ribosomal peptide synthetase-independent siderophore synthase [34].
3. Plant–AMF Interaction
In plants, iron serves as an indispensable cofactor for various enzymes involved in vital metabolic processes, and iron deficiency leads to diminished chlorophyll synthesis and photosynthesis, causing chlorosis and significant growth impairments [35]. To cope with iron deficiency, two main approaches are adopted by plants. The reduction strategy (strategy I) is mainly found in monocots and dicots [36]. Soil pH is decreased as a result of H+ extrusion by epidermal proton H+-ATPases on roots, thereby increasing the solubility of Fe3+. Solubilized Fe3+ is reduced to Fe2+ by a ferric reduction oxidase and transported to the root by iron-regulated transporters. In contrast, the chelation strategy (strategy II) is found in all monocots [37]. It relies on the excretion of mugineic acids that belong to the phytosiderophores family. Being complexed with phytosiderophores, iron is introduced into the plant by Yellow Stripe-like transporters that are structurally related to metal–nicotinamine complexes and play a critical role in iron homeostasis in plants [38].
Mycorrhizal symbioses provide a high amount of essential nutrients to host plants compared to those of non-mycorrhizal plants [39] (Figure 2). Kabir et al. [40] observed that AMF significantly increase the dry mass, width, and chlorophyll synthesis and photosynthesis parameters of sunflower cultivated under iron-deficient conditions, suggesting that AMF restore iron balance in sunflower plants. The improved nutrient uptake by mycorrhizal symbiosis through the wide hyphal network increases the accessible soil volume of plants [41]. The symbiotic association between plants and AMF stimulates the production of strigolactones, which are important for rhizosphere signaling and play a key role in stimulating hyphal branching [42]. In return, AMF makes distant resources available for plants.
In addition to spatial expansion, AMF can enhance iron uptake in plants through various mechanisms. One such mechanism is the release of low-molecular-weight organic compounds, such as oxalic acid and citric acid, which can solubilize insoluble forms of iron in soil [5]. Rahman et al. [14] found that acidification in the rhizosphere is a common response in plants adopting the reduction strategy, and the presence of AMF aggravates the decrease in pH.
AMF can enhance the expression of high-affinity iron transporters in plant roots, facilitating iron uptake from soil [33]. Jung et al. [43] demonstrated that AMF colonization upregulates the expression of iron-regulated transporter 1 (IRT1) and ferric reductase oxidase 2 (FRO2) genes, which are responsible for iron uptake from soil. Kabir et al. [40] also found that AMF upregulate another ferric reductase gene, HaFRO1, alongside the expression of transport genes HaNramp1 and HaIRT1. Moreover, in the context of iron homeostasis, natural resistance-associated macrophage protein transporters also play a key role, and members, such as RiSMF1, RiSMF2, RiSMF3.1, and RiSMF3.2, have been identified in R. irregularis [20,44].
AMF contribute to iron homeostasis by increasing the production of iron-chelating compounds, such as phytosiderophores, in plant roots. Li et al. [45] observed that two iron-phytosiderophore transporters are upregulated during AMF symbiosis in wheat roots, indicating a more substantial release of siderophores in wheat during symbiosis with AMF. The release of phytosiderophores increases in the presence of AMF cultured with Sorghum roots along with the expression of iron uptake-related genes SbDMAS2, SbNAS2, and SbYS1 [46]. Chorianopoulou et al. [47] observed that the enzyme family of nicotinamine-synthases, ZmNAS1 and ZmYS1, are severely overexpressed in non-mycorrhizal plants, while mycorrhizal plants do not intend to enhance the chelation strategy. AMF can even improve iron translocation within plants [48]. They enhance the expression of genes encoding transporters, such as iron-regulated transporter 3, involved in intracellular iron trafficking. This ensures that iron is properly distributed and utilized in various physiological processes in plant tissues to organs [16].
4. Bacteria–AMF Interaction
The interaction between bacteria and AMF in the endosphere and hyphosphere has gained significant attention in recent years. The hyphosphere refers to a more specific zone surrounding fungal hyphae. These interactions are mediated by a combination of physical associations, signaling pathways, and nutrient exchange, which in turn benefits plants by improving iron uptake and translocation within plants [49]. The bacteria–fungi interaction in the hyphosphere is influenced by various factors, including plant genotypes, environmental conditions, and a dynamic network of interactions within the plant biome [50]. Bacterial and fungal communities can differ depending on the developmental stages of plants, indicating a tight interaction with host physiology. Certain ectomycorrhizal fungi, such as Paxillus involutus and Suillus bovinus, have demonstrated the capability to host diverse bacterial communities, underscoring the presence of distinct niches within their mycorrhizospheres that exhibit high dependence on specific fungal species rather than host plants [51].
Bacteria–fungi interactions within the internal microenvironment of plants emerge as crucial modulators of iron dynamics. AMF play a pivotal role in the iron nutrition of plants, particularly in soils with low bioavailable iron. Both mycorrhizal and saprophytic fungi, along with bacteria, contribute to mineral weathering in the rhizosphere by inducing acidification [52]. Moreover, bacteria can produce siderophores and engage in synergistic relationships with fungi, which enhances this process. Bacterial strains derived from the hyphosphere of the ectomycorrhizal fungus Scleroderma citrinum exhibit a heightened capacity for mineral weathering compared to those sourced from the bulk soil [53]. Siderophores, which are essential for iron acquirement by soil microorganisms, have been identified as important factors in bacteria–fungi interactions. Bacteria can produce siderophores that are iron-chelating compounds and enhance iron availability and uptake by both bacteria and fungi [54]. Siderophores can influence the solubility and dissolution kinetics of iron oxides, leading to increased iron availability in the surrounding environment. Bacteria–fungi interactions can also influence the production of antifungal antibiotics by bacteria [55]. These antibiotics can inhibit the growth and activity of pathogenic fungi, thereby protecting the host plant from fungal infections [56]. The production of antifungal antibiotics by bacteria can contribute to iron homeostasis by preventing iron sequestration by pathogenic fungi and promoting iron availability for plants and beneficial microorganisms. Furthermore, bacteria–fungi interactions can be influenced by the regulation of iron uptake and homeostasis in both bacteria and fungi. Bacterial receptors, such as outer membrane TonB-dependent receptors, play a role in iron uptake from siderophores produced by other microorganisms [57]. Iron uptake in bacteria is tightly regulated, and the expression of iron acquisition systems is hierarchically controlled. Therefore, the regulation of iron uptake and homeostasis in both bacteria and fungi is crucial for maintaining iron balance and ensuring efficient iron utilization.
The hyphosphere is basically characterized by distinctive biotic and abiotic factors compared to those of the surrounding bulk soil, and it is an area of intense microbial activity and interactions [58]. AMF release hyphal compounds (e.g., organic acids and sugars) into soil to orchestrate the hyphosphere colonized by diverse microbes [59]. Plants also quickly channel recent photosynthetic products through ectomycorrhizal hyphae, facilitating direct interactions with bacteria in soil areas distant from roots [60]. Recent advances in molecular biology and metagenomics have started unraveling the mechanistic details of these interactions involving several bacterial genera (e.g., Microbacterium, Streptomyces, Bacillus, Enterobacter, and Rhizobium) that colonize hyphae. These bacteria are able to produce indole-3-acetic acid (IAAs) and siderophores, which can be beneficial to plants [61]. Sphingobacterium, which is commonly found in commercial AMF inoculants, exhibits resilience in herbicide-contaminated soils and shows plant growth-promoting traits such as solubilization of inorganic phosphates, surfactant production, and synthesis of IAAs [62]. In contrast, some studies indicate that soil bacteria affect plant growth by affecting the structure or elongation of hyphae [63]. When combined with AMF, Flavobacterium, Brevundimonas, Stenotrophomonas, and Devosia can enhance plant growth during periods of drought or salinity stress and facilitate spore germination of AMF [64]. Mycorrhiza helper bacteria can facilitate or enhance the colonization of AMF on plant roots by suppressing soil-borne pathogens or producing growth factors that stimulate spore germination or hyphal branching of AMF [65]. Paenibacillus validus has shown the potential to stimulate in vitro growth and spore formation in Rhizophagus irregularis [66]. Therefore, enhanced fungal growth may be beneficial in developing mycorrhizal association with plants. Some bacteria, in turn, can solubilize soil-bound nutrients or fix atmospheric nitrogen, which becomes accessible to plants via the mycorrhizal network [67]. This collaborative nutrient exchange benefits both plants and bacteria. Understanding the interactions between AMF, plants, and soil bacteria is crucial for appreciating the intricate dynamics of soil ecosystems.
Recent studies have underscored the role of soil bacteria in chelating iron, making it available to plants, specifically through the intricate network of fungal hyphae. Caris et al. [68] have noticed that iron can be transferred via fungal hyphae in soil to peanut and sorghum plants (Figure 2). Patel et al. [69] observed that iron uptake increases in plants connected to siderophore-producing bacteria through fungal hyphal networks. Furthermore, Verma et al. [70] reported that these bacteria–fungi associations can significantly improve plant growth under iron-limited conditions. Interestingly, Złoch et al. [71] noted that some plants can signal bacteria to produce more siderophores when experiencing iron deficiency. A comprehensive review by Liu et al. [20] highlighted the complex interplay between bacteria, fungi, and plants in the process of iron acquisition, shedding light on the potential for harnessing these interactions in sustainable agriculture. Chelation of iron by soil bacteria and its transfer via hyphae to plants are integral components of soil–plant–microbe interactions.
5. Siderophores, Iron Uptake, and Its Role in AMF Symbiosis
Understanding the interactions between AMF, plants, and soil bacteria is crucial for appreciating the intricate dynamics of soil ecosystems. Microorganisms require elevated concentrations of iron (approximately 10−6 M) for sustenance, triggering the onset of siderophore production when they perceive levels beneath this threshold [72]. Siderophore production can reshape interactions within the environmental microbial community, fostering cooperative or competitive mechanisms [73]. Notably, various types of fungal siderophores have been discovered in soil, mostly hydroxamate-type compounds with several complexities, spanning monomers, dimers, trimers, and cyclic forms that are categorized into three structural families, namely fusarinines, coprogens, and ferrichromes [74]. The ferrichromes also benefit other microorganisms, such as Pantoea, Enterobacter, Erwinia, and Yersinia, which contain receptors to uptake these siderophores [75].
Furthermore, bacterial siderophores are taken up by fungi such as Saccharomyces and Aspergillus [76]. Enterobactin, the principal siderophore generated by enterobacteria, can also be utilized by Saccharomyces, a microorganism that does not produce siderophores [75]. Battini et al. [77] identified bacterial strains from Rhizophagus intraradices spores that are capable of solubilizing phosphorus from organic (phytate) and inorganic sources and producing siderophores. The endobacterial symbiont Candidatus Glomeribacter gigasporarum [78,79], a Gram-negative bacteria affiliated with Burkholderiaceae, is found in the hyphae and spores of Gigaspora margarita, an AMF species [80]. Notably, the siderophore-producing genus Ralstonia also belongs to Burkholderiaceae, prompting speculation about the potential involvement of bacterial genomes in synthesizing carboxylate-type siderophores by G. margarita [34]. The filamentous structure of fungal mycelium, comprising extensive hyphal networks, traverses vast expanses of soil, reaching microenvironments often inaccessible to plant roots. This vast network, acting in tandem with siderophores, boosts the capacity of fungi to solubilize and acquire iron from distant sources.
Apart from this, the biosynthesis of phytosiderophores (produced by plants), as mentioned in Section 3, and fungal siderophores exhibit their respective pathway wherein phytosideropores, such as 2-deoxymugineic acid (DMA), are synthesized in vitro from both L-methionine and nicotianamine [81]. Ma et al. [82] and Sun et al. [83] demonstrated that DMA biosynthesis is associated with the methionine recycling pathway. As fungal siderophores are mostly hydroxamates, except for rhizoferrin (a carboxylate), they form a strong iron bidentate and require acylation with acetyl or anhydromevalonyl [84,85]. Plattner and Diekmann [86] have also explained the biosynthetic pathway of fungal siderophores by stating that hydroxamate siderophores are derived from L-ornithine and share N5-acyl-N5-hydroxyornithine, which has been repeatedly mentioned in studies about the involvement of genes in secondary metabolism [87].
The iron acquisition pathways in fungi, including AMF, involve common mechanisms; however, they exhibit a few differences. Fungi, including AMF, possess specific membrane transporters that recognize and transport iron–siderophore complexes into fungal cells [56]. Fungal transporters facilitate the internalization of iron-loaded siderophores; however, siderophore-mediated iron uptake in AMF is largely unexplored. Glomuferrin (bisimidorhizoferrin), a carboxylate-type siderophore produced by Glomeromycotina, has been observed in the root tissue of Tagetes patula nana [88]. Despite using a reductive pathway for iron uptake, R. irregularis lacks a Fet3p-type ferroxidase homolog in its genome, in contrast to the genomes of all phylogenetically investigated non-glomeromycotina fungi. In various fungal species, including Saccharomyces pombe, Phanerochaete chrysosporium, and Laccaria bicolor, a minimum of one Fet3 homolog is arranged in the genome with an Ftr1 homologue that regulates iron metabolism, which is lacking in AMF [89,90]. Previously, Caris et al. [68] and Kobae et al. [91] observed a radioactive signal in the shoots of peanut, sorghum, and maize host plants when 59Fe was supplied to the hyphae. However, the reductive pathway for iron uptake evolved by AMF is more similar to the reduction strategy of plants than that of other fungi. Furthermore, compared to non-mycorrhizal roots, mycorrhizal roots show increases in transcript levels of oligopeptide transporter genes such as ZmOPT8a and ZmOPT8b [91]. Major iron uptake analogs during iron uptake by AMF and plants are shown in Figure 3.
6. Future Perspectives and Challenges
As the significance of the bacteria–fungi–plant association in iron homeostasis comes to the fore, particularly regarding the role of siderophores, two main avenues emerge as being crucial in bridging current knowledge gaps. First, the intricate relationship between soil bacteria and AMF remains a topic of extensive interest. Siderophores produced by soil bacteria are instrumental in iron solubilization; however, how these molecules interface with AMF for subsequent iron uptake, transport, and translocation in plants is still elusive. Advanced molecular techniques, such as transcriptomic profiling of AMF in the presence of bacterial siderophores and isotopic labeling to track the movement of iron, may shed light on these mechanisms. Additionally, high-resolution imaging and computational modeling could provide invaluable insights into the spatio-temporal dynamics of siderophore-mediated interactions.
On a practical front, the next frontier lies in harnessing these molecular insights to devise effective strategies for optimizing iron uptake and homeostasis in plants. This necessitates a systems biology approach that integrates knowledge of siderophore production by bacteria, its uptake by AMF, and eventual assimilation by plants. Genetic engineering of both bacteria and AMF to potentially enhance siderophore production and uptake, respectively, may offer a promising avenue. However, field trials must be conducted to evaluate the feasibility and efficacy of these strategies, ensuring that laboratory successes translate to tangible agricultural benefits. Moreover, these endeavors must be underpinned by considerations of ecosystem balance, ensuring that any introduced change does not inadvertently destabilize existing microbial communities or plant health.
7. Conclusions
In conclusion, the interaction between plants and AMF plays a crucial role in iron homeostasis. AMF symbiosis enhances the expression of high-affinity iron transporters in plant roots, facilitating iron uptake from soil. Additionally, AMF can enhance the production of iron-chelating compounds, such as phytosiderophores, in plant roots. These mechanisms contribute to improved iron uptake and translocation within plants, leading to enhanced growth and nutrient acquisition. Interactions between bacteria and AMF in endophytes and the hyphosphere play crucial roles in iron dynamics. Bacteria can produce siderophores, thereby enhancing iron availability and uptake by both bacteria and fungi. The presence of siderophores in endophytes can influence the solubility and dissolution kinetics of iron oxides, leading to increased iron availability in plants. Bacteria–fungi interactions in the hyphosphere can help transport iron to plants from usually unreachable, distant places for plant roots. The regulation of iron uptake and homeostasis in both bacteria and fungi is crucial for maintaining iron balance and ensuring efficient iron utilization by plants.
Overall, understanding the intricate interactions among plants, AMF, and bacteria is essential for optimizing iron uptake and homeostasis in plants. Future studies should focus on unraveling the molecular mechanisms underlying these interactions and developing strategies to manipulate them for sustainable agriculture. Plant health and productivity can be improved by harnessing the potential of these interactions, thereby contributing to resilient and sustainable agricultural systems.
Author Contributions
Conceptualization, S.R. and T.K.L.; writing—original draft preparation, S.R. and B.R.K.; writing—review and editing, S.R., B.R.K. and T.K.L.; supervision, T.K.L.; funding acquisition, S.R., B.R.K. and T.K.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was carried out with the support of “Cooperative Research Program for Agriculture Science and Technology Development”, Rural Development Administration, Republic of Korea (PJ01710202).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Khaliq, A.; Perveen, S.; Alamer, K.H.; Ul Haq, M.Z.; Rafique, Z.; Alsudays, I.M.; Althobaiti, A.T.; Saleh, M.A.; Hussain, S.; Attia, H. Arbuscular Mycorrhizal Fungi Symbiosis to Enhance Plant-Soil Interaction. Sustainability 2022, 14, 7840. [Google Scholar] [CrossRef]
- Nasiri, K.; Babaeinejad, T.; Ghanavati, N.; Mohsenifar, K. Arbuscular mycorrhizal fungi affecting the growth, nutrient uptake and phytoremediation potential of different plants in a cadmium-polluted soil. Biometals 2022, 35, 1243–1253. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.W.; Shachar-Hill, Y. Sulfur transfer through an arbuscular mycorrhiza. Plant Physiol. 2009, 149, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Campos, C.; Carvalho, M.; Brigido, C.; Goss, M.J.; Nobre, T. Symbiosis Specificity of the Preceding Host Plant Can Dominate but Not Obliterate the Association Between Wheat and Its Arbuscular Mycorrhizal Fungal Partners. Front. Microbiol. 2018, 9, 2920. [Google Scholar] [CrossRef] [PubMed]
- Plassard, C.; Dell, B. Phosphorus nutrition of mycorrhizal trees. Tree Physiol. 2010, 30, 1129–1139. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, M.S.; Chong, K.P. Effects of Liming on Soil Properties and Its Roles in Increasing the Productivity and Profitability of the Oil Palm Industry in Malaysia. Agriculture 2022, 12, 322. [Google Scholar] [CrossRef]
- Hildebrandt, U.; Schmelzer, E.; Bothe, H. Expression of nitrate transporter genes in tomato colonized by an arbuscular mycorrhizal fungus. Physiol. Plant. 2002, 115, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Lurthy, T.; Cantat, C.; Jeudy, C.; Declerck, P.; Gallardo, K.; Barraud, C.; Leroy, F.; Ourry, A.; Lemanceau, P.; Salon, C.; et al. Impact of Bacterial Siderophores on Iron Status and Ionome in Pea. Front. Plant Sci. 2020, 11, 730. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Mou, X.; Meng, P.; Chen, J.; Tang, X.; Meng, G.; Xin, K.; Zhang, Y.; Wang, C. Effects of arbuscular mycorrhizal fungus inoculation on the growth and nitrogen metabolism of Catalpa bungei C.A.Mey. under different nitrogen levels. Front. Plant Sci. 2023, 14, 1138184. [Google Scholar] [CrossRef] [PubMed]
- Govindarajulu, M.; Pfeffer, P.E.; Jin, H.R.; Abubaker, J.; Douds, D.D.; Allen, J.W.; Bucking, H.; Lammers, P.J.; Shachar-Hill, Y. Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature 2005, 435, 819–823. [Google Scholar] [CrossRef]
- Sardans, J.; Lambers, H.; Preece, C.; Alrefaei, A.F.; Penuelas, J. Role of mycorrhizas and root exudates in plant uptake of soil nutrients (calcium, iron, magnesium, and potassium): Has the puzzle been completely solved? Plant J. 2023, 114, 1227–1242. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.C.; Wang, L.; Ma, F. Arbuscular mycorrhizal fungus modulates the phytotoxicity of Cd via combined responses of enzymes, thiolic compounds, and essential elements in the roots of Phragmites australis. Chemosphere 2017, 187, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.W.; Li, C.; Bai, T.H.; Wang, P. Nutrient use efficiency of plants under abiotic stress. Front. Plant Sci. 2023, 14, 1179842. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Parvin, M.; Das, U.; Ela, E.J.; Lee, S.H.; Lee, K.W.; Kabir, A.H. Arbuscular Mycorrhizal Symbiosis Mitigates Iron (Fe)-Deficiency Retardation in Alfalfa (Medicago sativa L.) Through the Enhancement of Fe Accumulation and Sulfur-Assisted Antioxidant Defense. Int. J. Mol. Sci. 2020, 21, 2219. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Li, Z.; Tang, M. Impact of salt and exogenous AM inoculation on indigenous microbial community structure in the rhizosphere of dioecious plant, Populus cathayana. Sci. Rep. 2021, 11, 18403. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-Y.; Xia, R.-X.; Hu, L.-M.; Dong, T.; Wu, Q.-S. Arbuscular mycorrhizal fungi alleviate iron deficient chlorosis in Poncirus trifoliata L. Raf under calcium bicarbonate stress. J. Hortic. Sci. Biotechnol. 2007, 82, 776–780. [Google Scholar] [CrossRef]
- Andrews, S.C.; Robinson, A.K.; Rodriguez-Quinones, F. Bacterial iron homeostasis. FEMS Microbiol. Rev. 2003, 27, 215–237. [Google Scholar] [CrossRef]
- Hider, R.C.; Kong, X.L. Chemistry and biology of siderophores. Nat. Prod. Rep. 2010, 27, 637–657. [Google Scholar] [CrossRef]
- Brundrett, M.C.; Tedersoo, L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef]
- Liu, Y.; Xiong, Z.M.; Wu, W.F.; Ling, H.Q.; Kong, D.Y. Iron in the Symbiosis of Plants and Microorganisms. Plants 2023, 12, 1958. [Google Scholar] [CrossRef]
- Marro, N.; Grilli, G.; Soteras, F.; Caccia, M.; Longo, S.; Cofre, N.; Borda, V.; Burni, M.; Janouskova, M.; Urcelay, C. The effects of arbuscular mycorrhizal fungal species and taxonomic groups on stressed and unstressed plants: A global meta-analysis. New Phytol. 2022, 235, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Zhen, L.; Songlin, W.; Yunjia, L.; Qing, Y.; Merinda, H.; Narottam, S.; Junjian, W.; Huang, Y.; Huang, L. Arbuscular mycorrhizal fungi regulate plant mineral nutrient uptake and partitioning in iron ore tailings undergoing eco-engineered pedogenesis. Pedosphere, 2023; in press. [Google Scholar] [CrossRef]
- Lanfranco, L.; Fiorilli, V.; Gutjahr, C. Partner communication and role of nutrients in the arbuscular mycorrhizal symbiosis. New Phytol. 2018, 220, 1031–1046. [Google Scholar] [CrossRef] [PubMed]
- Kwok, E.; Kosman, D. Iron in yeast: Mechanisms involved in homeostasis. Mol. Biol. Met. Homeost. Detoxif. Microbes Man 2006, 59–99. [Google Scholar] [CrossRef]
- Kosman, D.J. Molecular mechanisms of iron uptake in fungi. Mol. Microbiol. 2003, 47, 1185–1197. [Google Scholar] [CrossRef] [PubMed]
- de Souza, A.F.; Pigosso, L.L.; Silva, L.O.H.S.; Galo, I.D.C.; Paccez, J.D.; e Silva, K.S.F.; de Oliveira, M.A.P.; Pereira, M.; de Almeida Soares, C.M. Iron Deprivation Modulates the Exoproteome in Paracoccidioides brasiliensis. Front. Cell. Infect. Microbiol. 2022, 12, 903070. [Google Scholar] [CrossRef] [PubMed]
- Kornitzer, D.; Roy, U. Pathways of heme utilization in fungi. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2020, 1867, 118817. [Google Scholar] [CrossRef]
- Manoharan, S.; Ramalakshmi, O.I.; Ramasamy, S. Fungal siderophores: Prospects and applications. In Fungal Siderophores: From Mineral—Microbe Interactions to Anti-Pathogenicity; Springer: Cham, Switzerland, 2021; pp. 141–156. [Google Scholar] [CrossRef]
- Heymann, P.; Gerads, M.; Schaller, M.; Dromer, F.; Winkelmann, G.; Ernst, J.F. The siderophore iron transporter of Candida albicans (Sit1p/Arn1p) mediates uptake of ferrichrome-type siderophores and is required for epithelial invasion. Infect. Immun. 2002, 70, 5246–5255. [Google Scholar] [CrossRef]
- Martinez-Pastor, M.T.; Puig, S. Adaptation to iron deficiency in human pathogenic fungi. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2020, 1867, 118797. [Google Scholar] [CrossRef]
- Zhou, X.; Li, J.; Tang, N.; Xie, H.; Fan, X.; Chen, H.; Tang, M.; Xie, X. Genome-Wide Analysis of Nutrient Signaling Pathways Conserved in Arbuscular Mycorrhizal Fungi. Microorganisms 2021, 9, 1557. [Google Scholar] [CrossRef]
- Tamayo, E.; Gomez-Gallego, T.; Azcon-Aguilar, C.; Ferrol, N. Genome-wide analysis of copper, iron and zinc transporters in the arbuscular mycorrhizal fungus Rhizophagus irregularis. Front. Plant Sci. 2014, 5, 547. [Google Scholar] [CrossRef] [PubMed]
- Tamayo, E.; Shim, C.; Castillo, A.; Benz, J.; Ferrol, N. Characterization of the Rhizophagus irregularis multicopper oxidase family indicates that the iron transporter RiFTR1 does not require a ferroxidase partner. bioRxiv 2022. [Google Scholar] [CrossRef]
- Haselwandter, K.; Haas, H.; Haninger, G.; Winkelmann, G. Siderophores in plant root tissue: Tagetes patula nana colonized by the arbuscular mycorrhizal fungus Gigaspora margarita. Biometals 2020, 33, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Ferrol, N.; Tamayo, E.; Vargas, P. The heavy metal paradox in arbuscular mycorrhizas: From mechanisms to biotechnological applications. J. Exp. Bot. 2016, 67(22), 6253–6265. [Google Scholar] [CrossRef] [PubMed]
- Wairich, A.; de Oliveira, B.H.N.; Arend, E.B.; Duarte, G.L.; Ponte, L.R.; Sperotto, R.A.; Ricachenevsky, F.K.; Fett, J.P. The combined strategy for iron uptake is not exclusive to domesticated rice (Oryza sativa). Sci. Rep. 2019, 9, 16144. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.; Shayaba; Kumar, R.; Kumar, J.; Bhadana, D.; Batra, R.; Singh, R.; Kumar, S.; Roy, J.K.; Balyan, H.S. Comparative analysis of VMT genes/proteins in selected plant species with emphasis on bread wheat (Triticum aestivum L.). Genes Genom. 2023, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.A.; Guo, J.; Peng, H.; Tian, S.X.; Bai, X.X.; Zhu, H.C.; Kang, Z.S.; Guo, J. TaYS1A, a Yellow Stripe-Like Transporter Gene, Is Required for Wheat Resistance to Puccinia striiformis f. sp. Tritici. Genes 2020, 11, 1452. [Google Scholar] [CrossRef] [PubMed]
- Jacott, C.N.; Murray, J.D.; Ridout, C.J. Trade-offs in arbuscular mycorrhizal symbiosis: Disease resistance, growth responses and perspectives for crop breeding. Agronomy 2017, 7, 75. [Google Scholar] [CrossRef]
- Kabir, A.H.; Debnath, T.; Das, U.; Prity, S.A.; Haque, A.; Rahman, M.M.; Parvez, M.S. Arbuscular mycorrhizal fungi alleviate Fe-deficiency symptoms in sunflower by increasing iron uptake and its availability along with antioxidant defense. Plant Physiol. Biochem. 2020, 150, 254–262. [Google Scholar] [CrossRef]
- Bolan, N. A critical review on the role of mycorrhizal fungi in the uptake of phosphorus by plants. Plant Soil 1991, 134, 189–207. [Google Scholar] [CrossRef]
- Besserer, A.; Puech-Pagès, V.; Kiefer, P.; Gomez-Roldan, V.; Jauneau, A.; Roy, S.; Portais, J.-C.; Roux, C.; Bécard, G.; Séjalon-Delmas, N. Strigolactones stimulate arbuscular mycorrhizal fungi by activating mitochondria. PLoS Biol. 2006, 4, e226. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-induced resistance and priming of plant defenses. J. Chem. Ecol. 2012, 38, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Lorca, V.M.; Molina-Luzon, M.J.; Ferrol, N. Characterization of the NRAMP Gene Family in the Arbuscular Mycorrhizal Fungus Rhizophagus irregularis. J. Fungi 2022, 8, 592. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, R.; Tian, H.; Gao, Y. Transcriptome responses in wheat roots to colonization by the arbuscular mycorrhizal fungus Rhizophagus irregularis. Mycorrhiza 2018, 28, 747–759. [Google Scholar] [CrossRef] [PubMed]
- Prity, S.A.; Sajib, S.A.; Das, U.; Rahman, M.M.; Haider, S.A.; Kabir, A.H. Arbuscular mycorrhizal fungi mitigate Fe deficiency symptoms in sorghum through phytosiderophore-mediated Fe mobilization and restoration of redox status. Protoplasma 2020, 257, 1373–1385. [Google Scholar] [CrossRef] [PubMed]
- Chorianopoulou, S.N.; Saridis, Y.I.; Dimou, M.; Katinakis, P.; Bouranis, D.L. Arbuscular mycorrhizal symbiosis alters the expression patterns of three key iron homeostasis genes, ZmNAS1, ZmNAS3, and ZmYS1, in S deprived maize plants. Front. Plant Sci. 2015, 6, 257. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Nishizawa, N.K. Iron uptake, translocation, and regulation in higher plants. Annu. Rev. Plant Biol. 2012, 63, 131–152. [Google Scholar] [CrossRef]
- Sepp, S.K.; Davison, J.; Jairus, T.; Vasar, M.; Moora, M.; Zobel, M.; Öpik, M. Non-random association patterns in a plant–mycorrhizal fungal network reveal host–symbiont specificity. Mol. Ecol. 2019, 28, 365–378. [Google Scholar] [CrossRef]
- Hardoim, P.R.; Van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef]
- Desai, S.; Naik, D.; Cumming, J.R. The influence of phosphorus availability and Laccaria bicolor symbiosis on phosphate acquisition, antioxidant enzyme activity, and rhizospheric carbon flux in Populus tremuloides. Mycorrhiza 2014, 24, 369–382. [Google Scholar] [CrossRef]
- Koshila Ravi, R.; Anusuya, S.; Balachandar, M.; Muthukumar, T. Microbial interactions in soil formation and nutrient cycling. Mycorrhizosphere Pedogenesis 2019, 363–382. [Google Scholar] [CrossRef]
- Sun, Q.; Liu, X.; Wang, S.; Lian, B. Effects of mineral substrate on ectomycorrhizal fungal colonization and bacterial community structure. Sci. Total Environ. 2020, 721, 137663. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Geat, N.; Rajawat, M.V.S.; Prasanna, R.; Kar, A.; Singh, A.M.; Saxena, A.K. Prospecting endophytes from different Fe or Zn accumulating wheat genotypes for their influence as inoculants on plant growth, yield, and micronutrient content. Ann. Microbiol. 2018, 68, 815–833. [Google Scholar] [CrossRef]
- Ryan, R.P.; Germaine, K.; Franks, A.; Ryan, D.J.; Dowling, D.N. Bacterial endophytes: Recent developments and applications. FEMS Microbiol. Lett. 2008, 278, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Haas, H.; Eisendle, M.; Turgeon, B.G. Siderophores in fungal physiology and virulence. Annu. Rev. Phytopathol. 2008, 46, 149–187. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, P. Iron uptake and metabolism in pseudomonads. Appl. Microbiol. Biotechnol. 2010, 86, 1637–1645. [Google Scholar] [CrossRef] [PubMed]
- Jansa, J.; Hodge, A. Swimming, gliding, or hyphal riding? On microbial migration along the arbuscular mycorrhizal hyphal highway and functional consequences thereof. New Phytol. 2021, 230, 14–16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, J.; George, T.S.; Limpens, E.; Feng, G. Arbuscular mycorrhizal fungi conducting the hyphosphere bacterial orchestra. Trends Plant Sci. 2022, 27(4), 402–411. [Google Scholar] [CrossRef] [PubMed]
- Gorka, S.; Dietrich, M.; Mayerhofer, W.; Gabriel, R.; Wiesenbauer, J.; Martin, V.; Zheng, Q.; Imai, B.; Prommer, J.; Weidinger, M. Rapid transfer of plant photosynthates to soil bacteria via ectomycorrhizal hyphae and its interaction with nitrogen availability. Front. Microbiol. 2019, 10, 168. [Google Scholar] [CrossRef]
- Agnolucci, M.; Avio, L.; Pepe, A.; Turrini, A.; Cristani, C.; Bonini, P.; Cirino, V.; Colosimo, F.; Ruzzi, M.; Giovannetti, M. Bacteria associated with a commercial mycorrhizal inoculum: Community composition and multifunctional activity as assessed by Illumina sequencing and culture-dependent tools. Front. Plant Sci. 2019, 9, 1956. [Google Scholar] [CrossRef]
- Nacoon, S.; Jogloy, S.; Riddech, N.; Mongkolthanaruk, W.; Kuyper, T.W.; Boonlue, S. Interaction between phosphate solubilizing bacteria and arbuscular mycorrhizal fungi on growth promotion and tuber inulin content of Helianthus tuberosus L. Sci. Rep. 2020, 10, 4916. [Google Scholar] [CrossRef] [PubMed]
- Giovannini, L.; Palla, M.; Agnolucci, M.; Avio, L.; Sbrana, C.; Turrini, A.; Giovannetti, M. Arbuscular mycorrhizal fungi and associated microbiota as plant biostimulants: Research strategies for the selection of the best performing inocula. Agronomy 2020, 10, 106. [Google Scholar] [CrossRef]
- Talwar, C.; Nagar, S.; Kumar, R.; Scaria, J.; Lal, R.; Negi, R.K. Defining the environmental adaptations of genus Devosia: Insights into its expansive short peptide transport system and positively selected genes. Sci. Rep. 2020, 10, 1151. [Google Scholar] [CrossRef] [PubMed]
- Sangwan, S.; Prasanna, R. Mycorrhizae helper bacteria: Unlocking their potential as bioenhancers of plant–arbuscular mycorrhizal fungal associations. Microb. Ecol. 2022, 84, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Hashimoto, K.; Kobayashi, Y.; Yano, K.; Maeda, T.; Kameoka, H.; Ezawa, T.; Saito, K.; Akiyama, K.; Kawaguchi, M. Asymbiotic mass production of the arbuscular mycorrhizal fungus Rhizophagus clarus. Commun. Biol. 2022, 5, 43. [Google Scholar] [CrossRef] [PubMed]
- van’t Padje, A.; Oyarte Galvez, L.; Klein, M.; Hink, M.A.; Postma, M.; Shimizu, T.; Kiers, E.T. Temporal tracking of quantum-dot apatite across in vitro mycorrhizal networks shows how host demand can influence fungal nutrient transfer strategies. ISME J. 2021, 15, 435–449. [Google Scholar] [CrossRef] [PubMed]
- Caris, C.; Hordt, W.; Hawkins, H.J.; Romheld, V.; George, E. Studies of iron transport by arbuscular mycorrhizal hyphae from soil to peanut and sorghum plants. Mycorrhiza 1998, 8, 35–39. [Google Scholar] [CrossRef]
- Patel, P.; Trivedi, G.; Saraf, M. Iron biofortification in mungbean using siderophore producing plant growth promoting bacteria. Environ. Sustain. 2018, 1, 357–365. [Google Scholar] [CrossRef]
- Verma, N.; Narayan, O.P.; Prasad, D.; Jogawat, A.; Panwar, S.L.; Dua, M.; Johri, A.K. Functional characterization of a high-affinity iron transporter (PiFTR) from the endophytic fungus Piriformospora indica and its role in plant growth and development. Environ. Microbiol. 2022, 24, 689–706. [Google Scholar] [CrossRef]
- Złoch, M.; Thiem, D.; Gadzała-Kopciuch, R.; Hrynkiewicz, K. Synthesis of siderophores by plant-associated metallotolerant bacteria under exposure to Cd2+. Chemosphere 2016, 156, 312–325. [Google Scholar] [CrossRef]
- Miethke, M.; Marahiel, M.A. Siderophore-based iron acquisition and pathogen control. Microbiol. Mol. Biol. Rev. 2007, 71, 413–451. [Google Scholar] [CrossRef] [PubMed]
- Weiland-Bräuer, N. Friends or foes—Microbial interactions in nature. Biology 2021, 10, 496. [Google Scholar] [CrossRef] [PubMed]
- Renshaw, J.C.; Robson, G.D.; Trinci, A.P.J.; Wiebe, M.G.; Livens, F.R.; Collison, D.; Taylor, R.J. Fungal siderophores: Structures, functions and applications. Mycol. Resesrch 2002, 106, 1123–1142. [Google Scholar] [CrossRef]
- Winkelmann, J.; Schormair, B.; Lichtner, P.; Ripke, S.; Xiong, L.; Jalilzadeh, S.; Fulda, S.; Pütz, B.; Eckstein, G.; Hauk, S. Genome-wide association study of restless legs syndrome identifies common variants in three genomic regions. Nat. Genet. 2007, 39, 1000–1006. [Google Scholar] [CrossRef]
- Khan, A.; Singh, P.; Kumar, R.; Das, S.; Singh, R.K.; Mina, U.; Agrawal, G.K.; Rakwal, R.; Sarkar, A.; Srivastava, A. Antifungal activity of siderophore isolated from Escherichia coli against Aspergillus nidulans via iron-mediated oxidative stress. Front. Microbiol. 2021, 12, 729032. [Google Scholar] [CrossRef] [PubMed]
- Battini, F.; Cristani, C.; Giovannetti, M.; Agnolucci, M. Multifunctionality and diversity of culturable bacterial communities strictly associated with spores of the plant beneficial symbiont Rhizophagus intraradices. Microbiol. Res. 2016, 183, 68–79. [Google Scholar] [CrossRef]
- Bianciotto, V.; Genre, A.; Jargeat, P.; Lumini, E.; Bécard, G.; Bonfante, P. Vertical transmission of endobacteria in the arbuscular mycorrhizal fungus Gigaspora margarita through generation of vegetative spores. Appl. Environ. Microbiol. 2004, 70, 3600–3608. [Google Scholar] [CrossRef]
- Ouledali, S.; Lumini, E.; Bianciotto, V.; Khemira, H.; Ennajeh, M. Diversity of Arbuscular Mycorrhizal Fungi in olive orchard soils in arid regions of Southern Tunisia. Arid Land Res. Manag. 2022, 36, 411–427. [Google Scholar] [CrossRef]
- Desiro, A.; Salvioli, A.; Ngonkeu, E.L.; Mondo, S.J.; Epis, S.; Faccio, A.; Kaech, A.; Pawlowska, T.E.; Bonfante, P. Detection of a novel intracellular microbiome hosted in arbuscular mycorrhizal fungi. ISME J. 2014, 8, 257–270. [Google Scholar] [CrossRef]
- Murata, Y.; Murata, J.; Namba, K. Unraveling the new biological roles and possible applications of phytosiderophores in plants and mammals. Met. Res. 2022, 2, rev-1–reg-11. [Google Scholar] [CrossRef]
- Ma, J.F.; Shinada, T.; Matsuda, C.; Nomoto, K. Biosynthesis of Phytosiderophores, Mugineic Acids, Associated with Methionine Cycling. J. Biol. Chem. 1995, 270, 16549–16554. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.T.; Zhou, X.J.; Chen, C.; Zhang, X.; Tian, X.L.; Xiao, K.; Liu, C.X.; Chen, R.M.; Chen, S.J. Maize Interveinal Chlorosis 1 links the Yang Cycle and Fe homeostasis through Nicotianamine biosynthesis. Plant Physiol. 2022, 188, 2131–2145. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L. Iron and siderophores in fungal-host interactions. Mycol. Res. 2008, 112, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Pecoraro, L.; Wang, X.; Shah, D.W.; Song, X.X.; Kumar, V.; Shakoor, A.; Tripathi, K.; Ramteke, P.W.; Rani, R. Biosynthesis Pathways, Transport Mechanisms and Biotechnological Applications of Fungal Siderophores. J. Fungi 2022, 8, 21. [Google Scholar] [CrossRef] [PubMed]
- Plattner, H.J.; Diekmann, H. Enzymology of siderophore biosynthesis in fungi. In Metal Ions in Fungi; CRC Press: Boca Raton, FL, USA, 2020; pp. 99–116. [Google Scholar]
- Sarrocco, S.; Vicente, I.; Staropoli, A.; Vinale, F. Genes Involved in the Secondary Metabolism of Trichoderma and the Biochemistry of These Compounds. In Advances in Trichoderma Biology for Agricultural Applications; Springer: Berlin/Heidelberg, Germany, 2022; pp. 113–135. [Google Scholar]
- Winkelmann, G. A search for glomuferrin: A potential siderophore of arbuscular mycorrhizal fungi of the genus Glomus. Biometals 2017, 30, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Canessa, P.; Larrondo, L.F. Environmental responses and the control of iron homeostasis in fungal systems. Appl. Microbiol. Biotechnol. 2013, 97, 939–955. [Google Scholar] [CrossRef] [PubMed]
- Garcia, K.; Doidy, J.; Zimmermann, S.D.; Wipf, D.; Courty, P.E. Take a Trip Through the Plant and Fungal Transportome of Mycorrhiza. Trends Plant Sci. 2016, 21, 937–950. [Google Scholar] [CrossRef]
- Kobae, Y.; Tomioka, R.; Tanoi, K.; Kobayashi, N.I.; Ohmori, Y.; Nishida, S.; Fujiwara, T. Selective induction of putative iron transporters, OPT8a and OPT8b, in maize by mycorrhizal colonization. Soil Sci. Plant Nutr. 2014, 60, 843–847. [Google Scholar] [CrossRef]
Figure 1.
The four major iron uptake mechanisms in fungi. (i) Direct Fe2+ permeation, (ii) coupled ferroxidase/permease uptake, (iii) heme/hemin uptake, and (iv) siderophore-mediated iron uptake. In AMF, a putative ferrireductase RiFRE1 and iron permease RiFTR1 have been discovered recently. Other mechanisms are still not clear in AMF.
Figure 1.
The four major iron uptake mechanisms in fungi. (i) Direct Fe2+ permeation, (ii) coupled ferroxidase/permease uptake, (iii) heme/hemin uptake, and (iv) siderophore-mediated iron uptake. In AMF, a putative ferrireductase RiFRE1 and iron permease RiFTR1 have been discovered recently. Other mechanisms are still not clear in AMF.

Figure 2.
Importance of symbiotic relationship of arbuscular mycorrhizal fungi in plant growth. Arbuscular mycorrhizal fungi regulate plant metabolism in the rhizosphere to enhance iron uptake, and in the hyphosphere to make distant iron available for plants. Brown and pink zone represent rhizosphere and hyphosphere, respectively.
Figure 2.
Importance of symbiotic relationship of arbuscular mycorrhizal fungi in plant growth. Arbuscular mycorrhizal fungi regulate plant metabolism in the rhizosphere to enhance iron uptake, and in the hyphosphere to make distant iron available for plants. Brown and pink zone represent rhizosphere and hyphosphere, respectively.

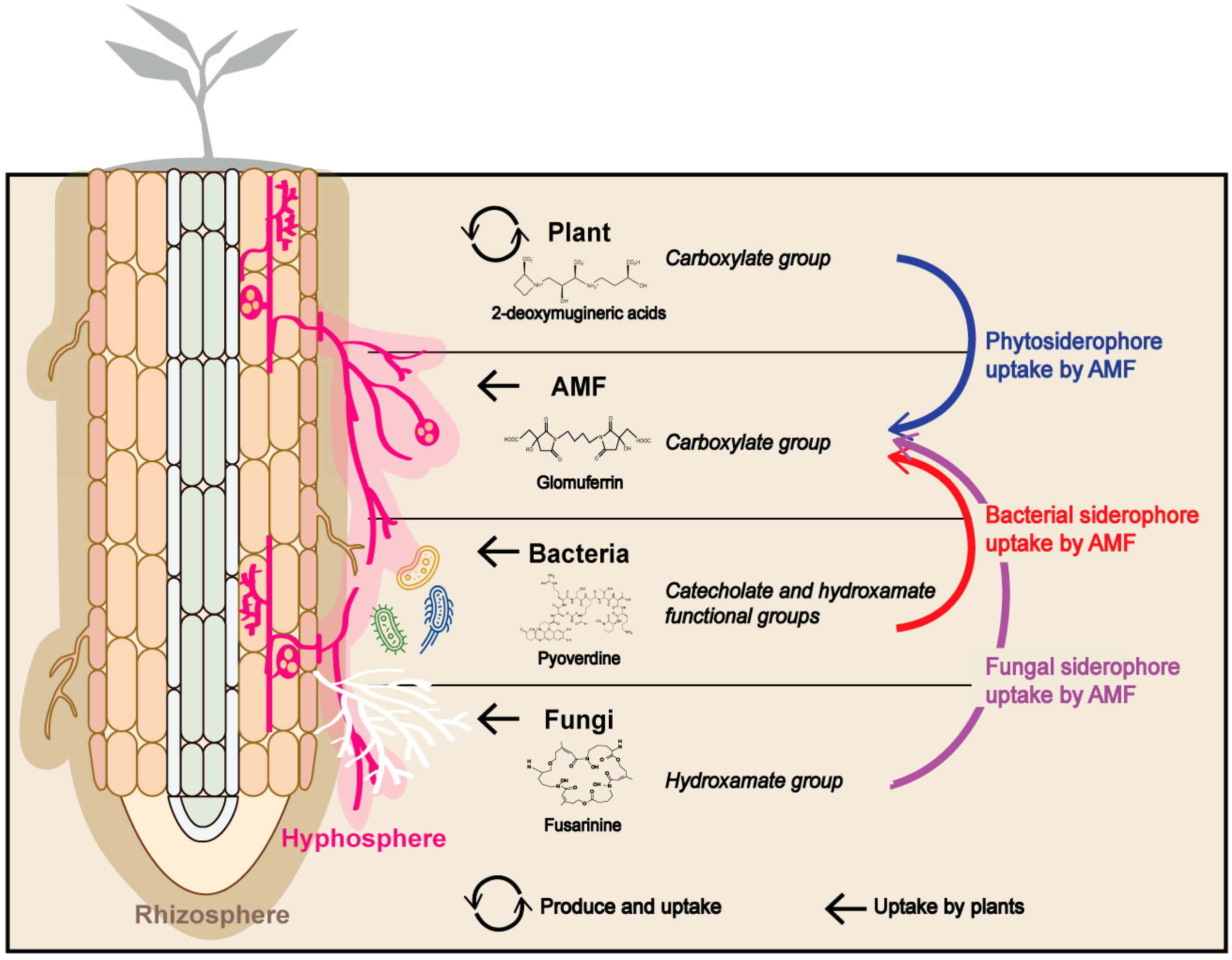
Figure 3.
Production of siderophores by host plants and soil microorganisms. Arbuscular mycorrhizal fungi play an important role in maximizing iron uptake efficiency using phytosiderophores and in the uptake or transfer of iron chelated with siderophores produced by other soil microorganisms including bacteria, fungi, and arbuscular mycorrhizal fungi. Brown and pink zone represent rhizosphere and hyphosphere, respectively.
Figure 3.
Production of siderophores by host plants and soil microorganisms. Arbuscular mycorrhizal fungi play an important role in maximizing iron uptake efficiency using phytosiderophores and in the uptake or transfer of iron chelated with siderophores produced by other soil microorganisms including bacteria, fungi, and arbuscular mycorrhizal fungi. Brown and pink zone represent rhizosphere and hyphosphere, respectively.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rajapitamahuni, S.; Kang, B.R.; Lee, T.K. Exploring the Roles of Arbuscular Mycorrhizal Fungi in Plant–Iron Homeostasis. Agriculture 2023, 13, 1918. https://doi.org/10.3390/agriculture13101918
AMA Style
Rajapitamahuni S, Kang BR, Lee TK. Exploring the Roles of Arbuscular Mycorrhizal Fungi in Plant–Iron Homeostasis. Agriculture. 2023; 13(10):1918. https://doi.org/10.3390/agriculture13101918
Chicago/Turabian StyleRajapitamahuni, Soundarya, Bo Ram Kang, and Tae Kwon Lee. 2023. "Exploring the Roles of Arbuscular Mycorrhizal Fungi in Plant–Iron Homeostasis" Agriculture 13, no. 10: 1918. https://doi.org/10.3390/agriculture13101918
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.