


Influence of Soil Type, Land Use, and Rootstock Genotype on Root-Associated Arbuscular Mycorrhizal Fungi Communities and Their Impact on Grapevine Growth and Nutrition

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Areas and Soil Sampling
2.2. Grapevine Trap Culture Establishment
2.3. Growth, Nutrition, and Mycorrhizal Colonization Assessment in Grapevine Plants
2.4. Analysis of Mycorrhizal Community Characteristics
2.5. Statistical Analyses
3. Results
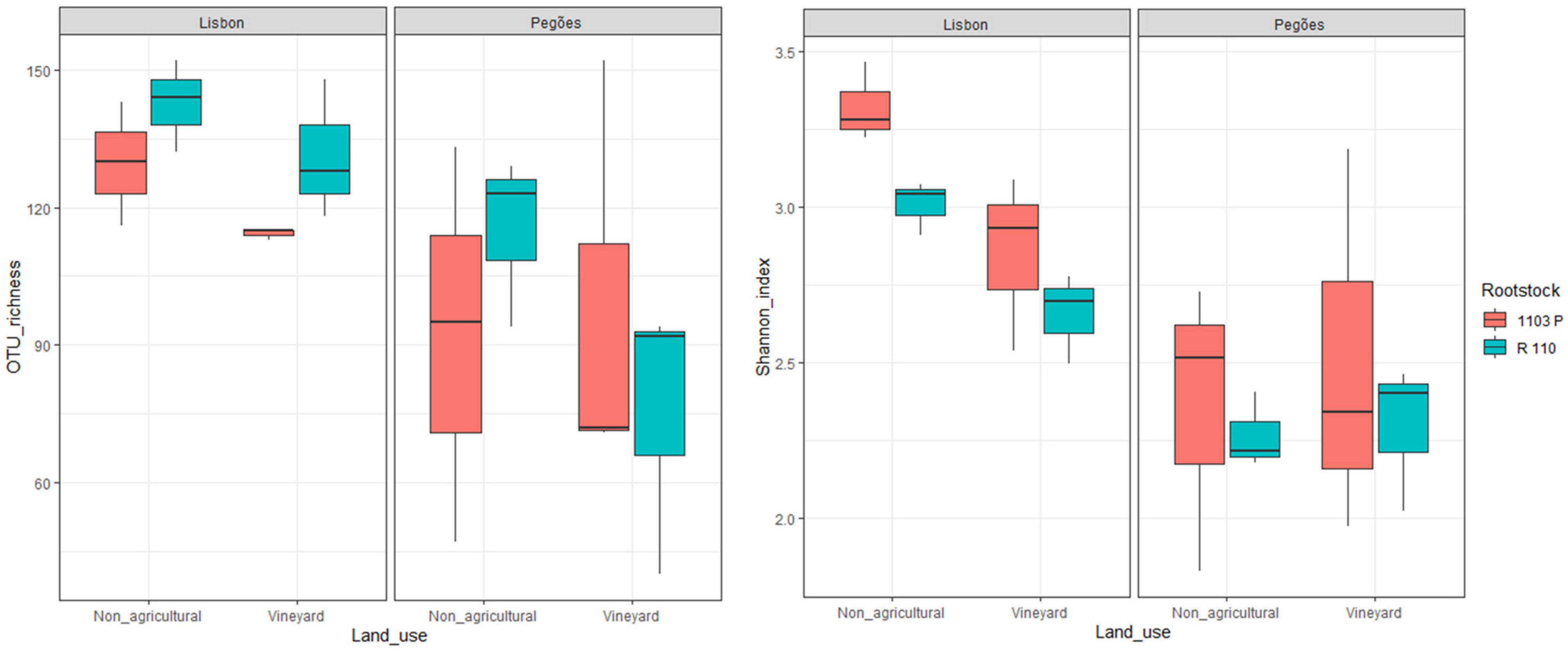
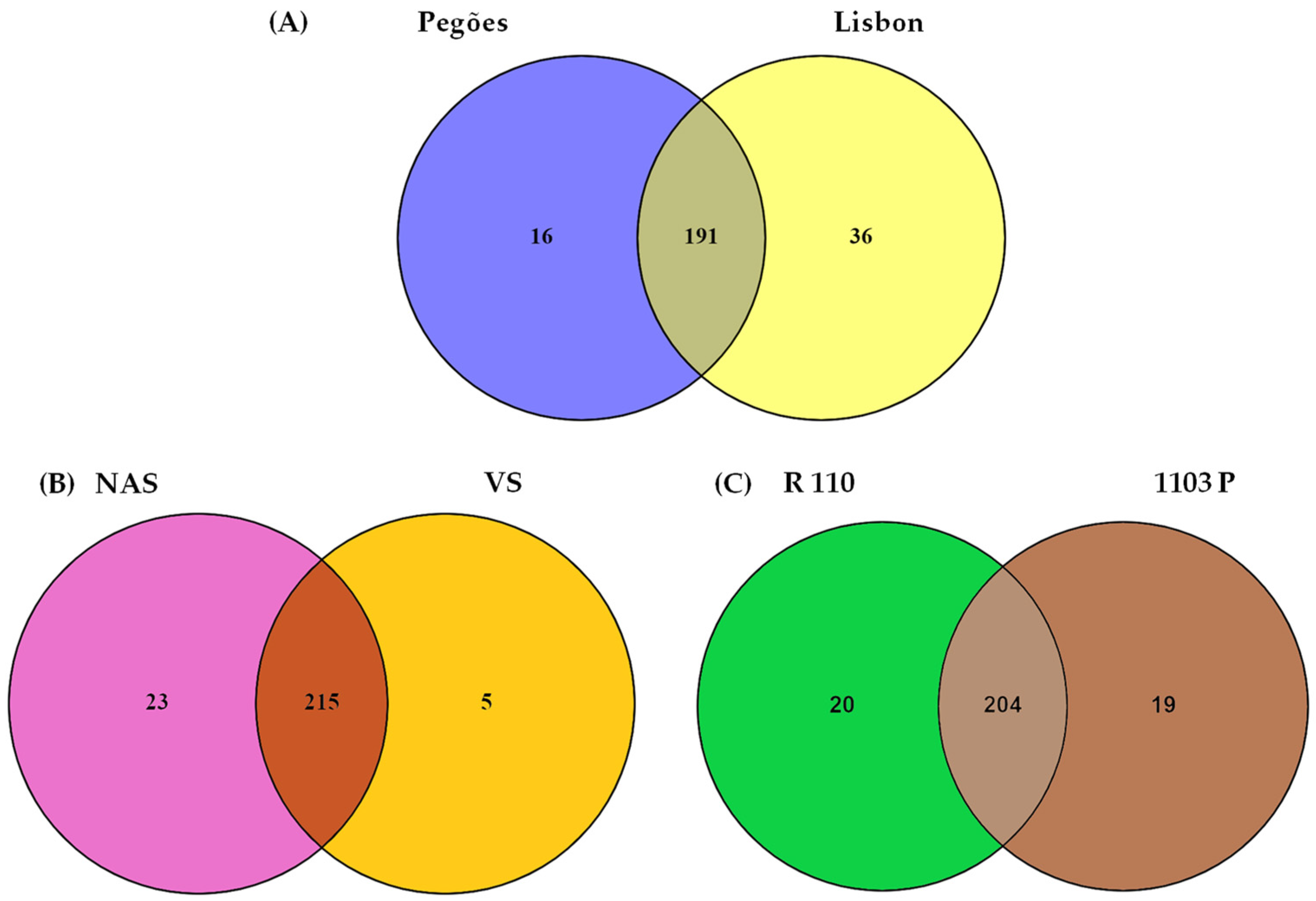
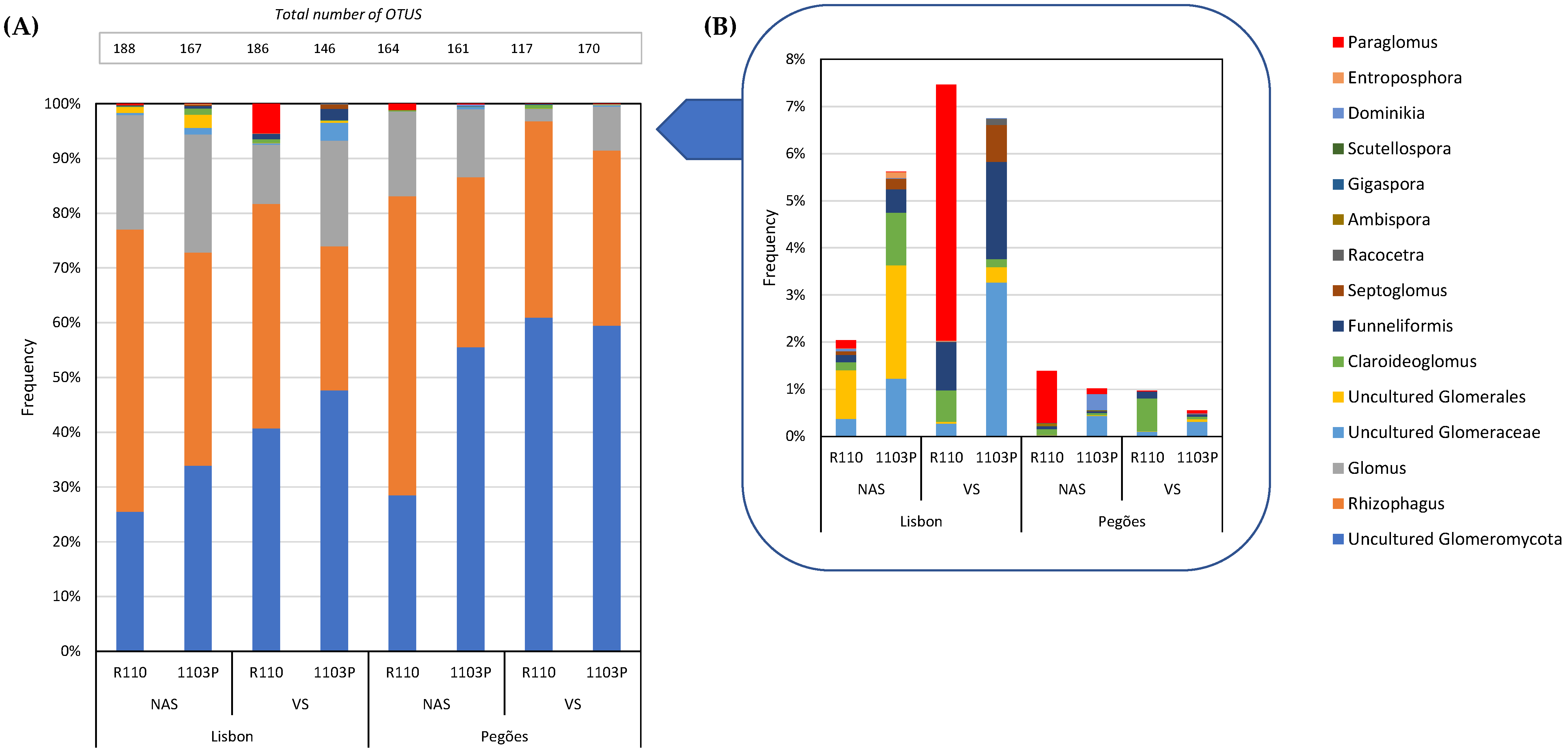
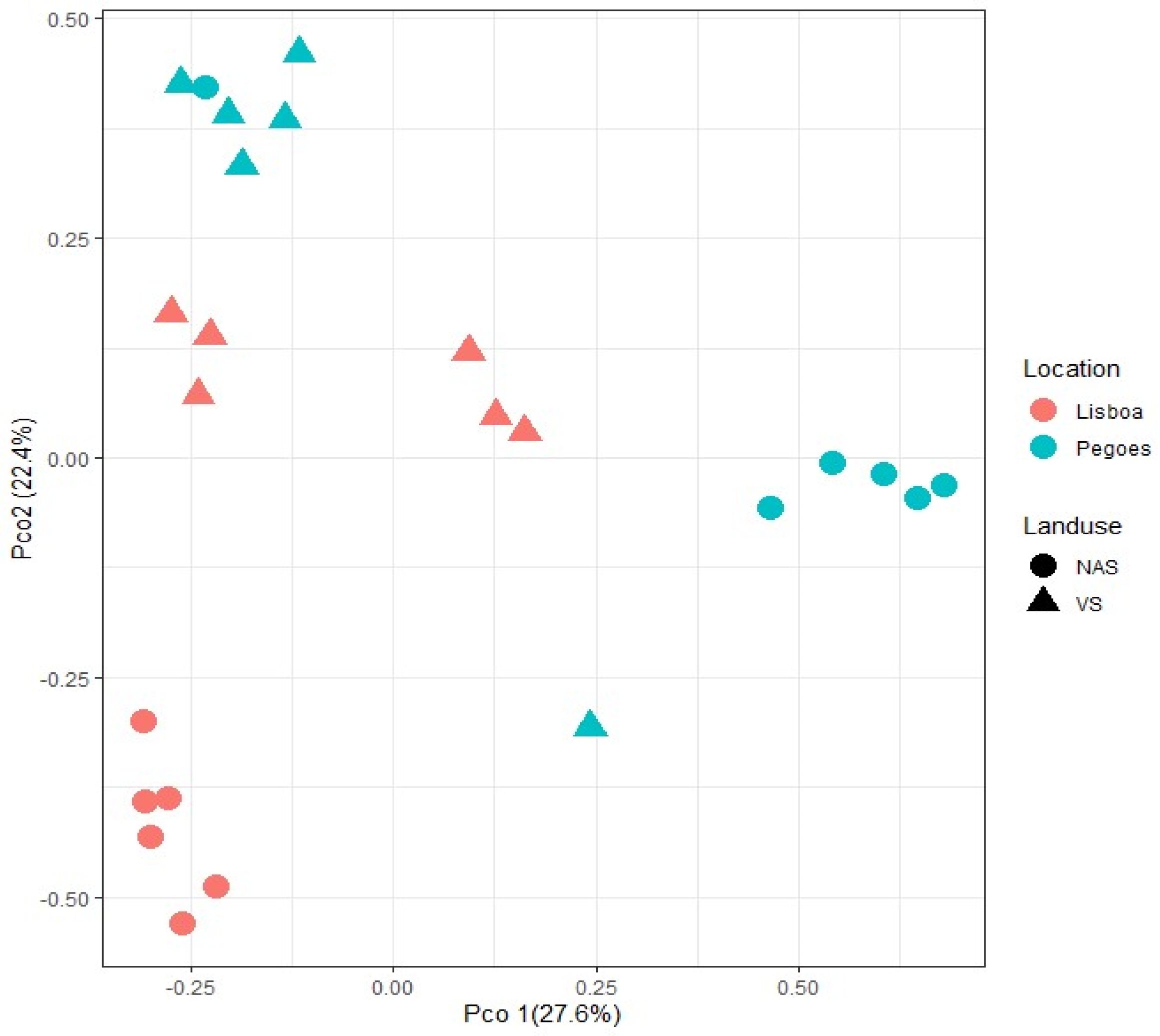
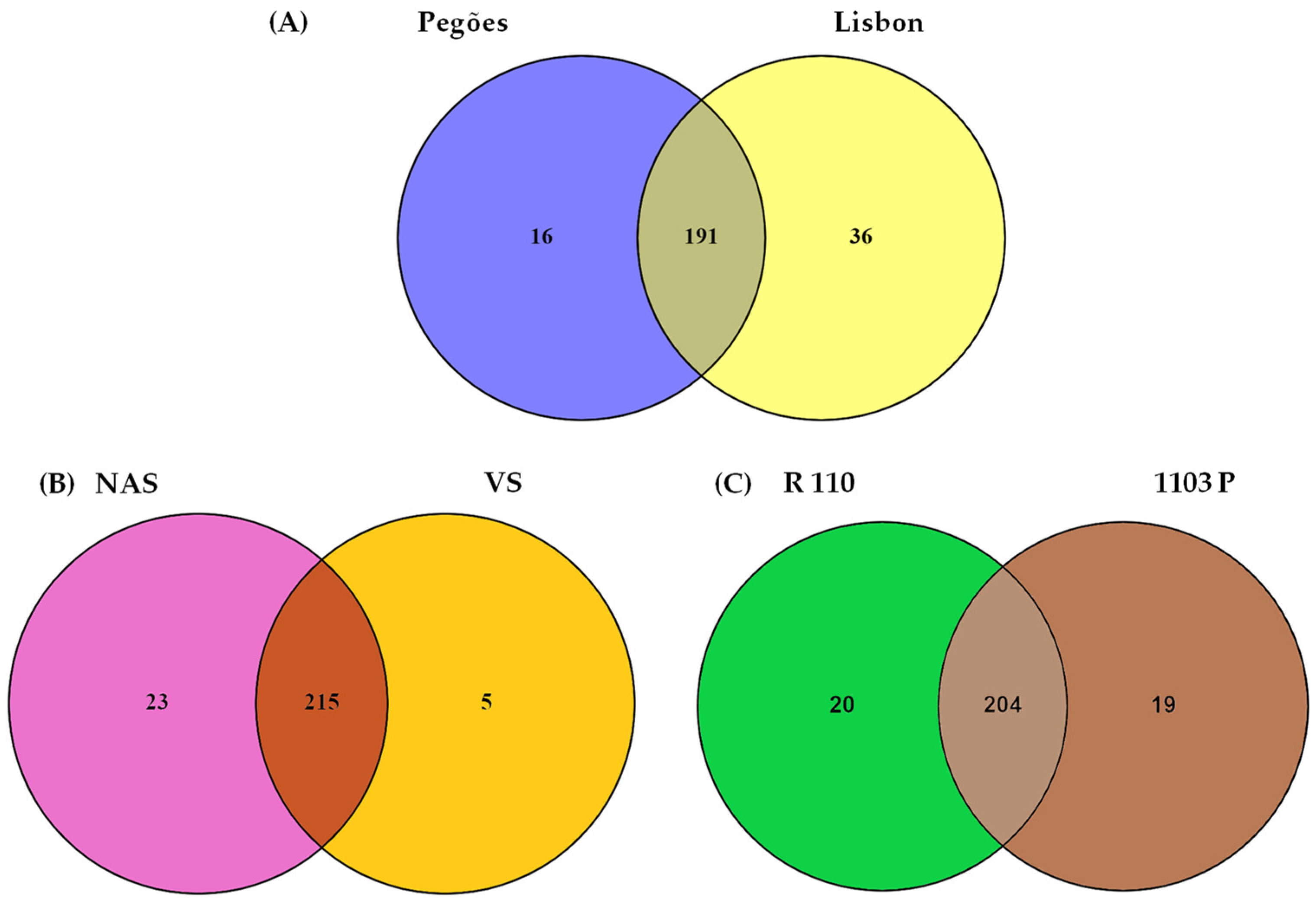
3.1. Soil Type and Land Use Effect on the Diversity of Culturable Mycorrhizal Communities
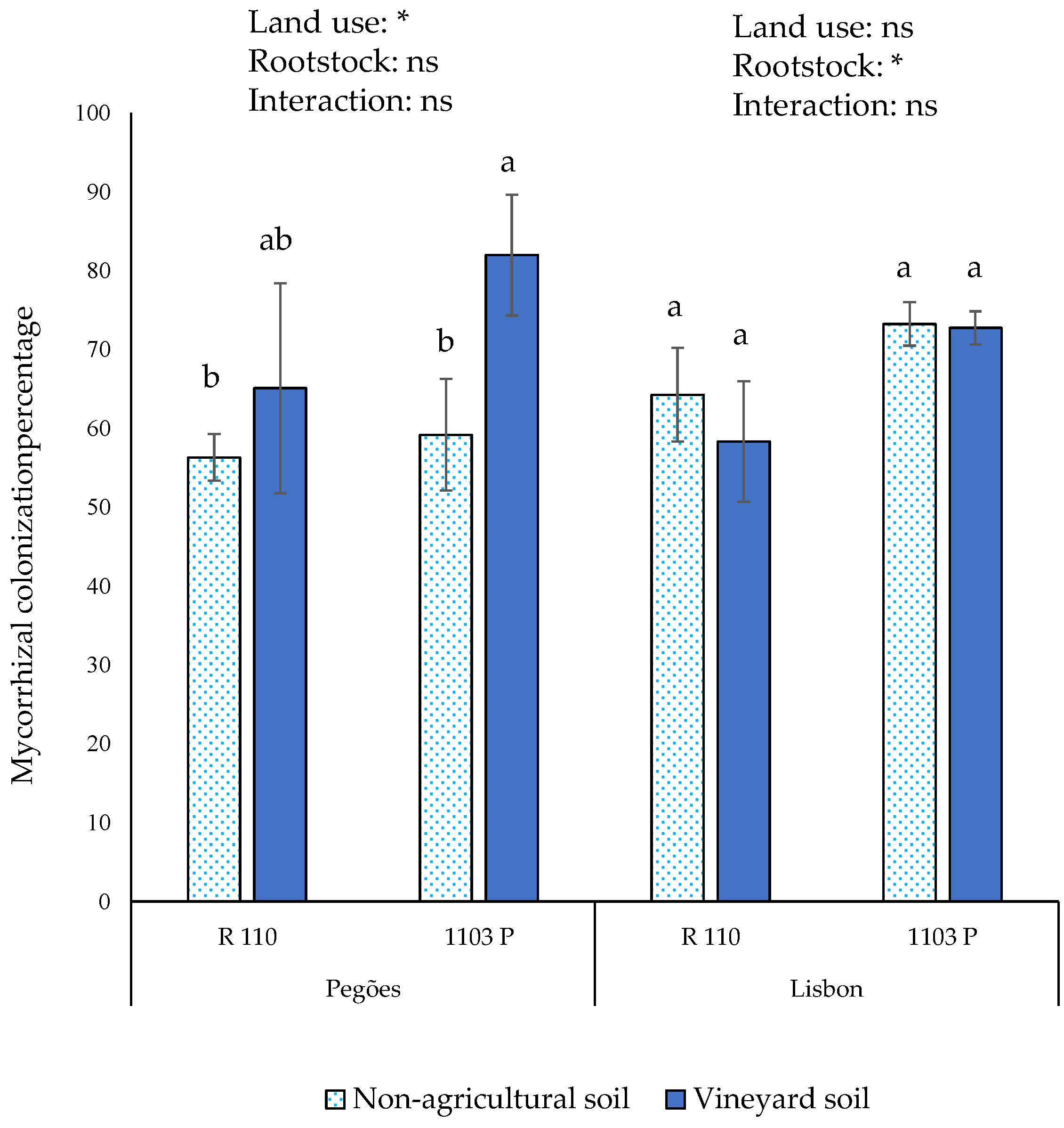
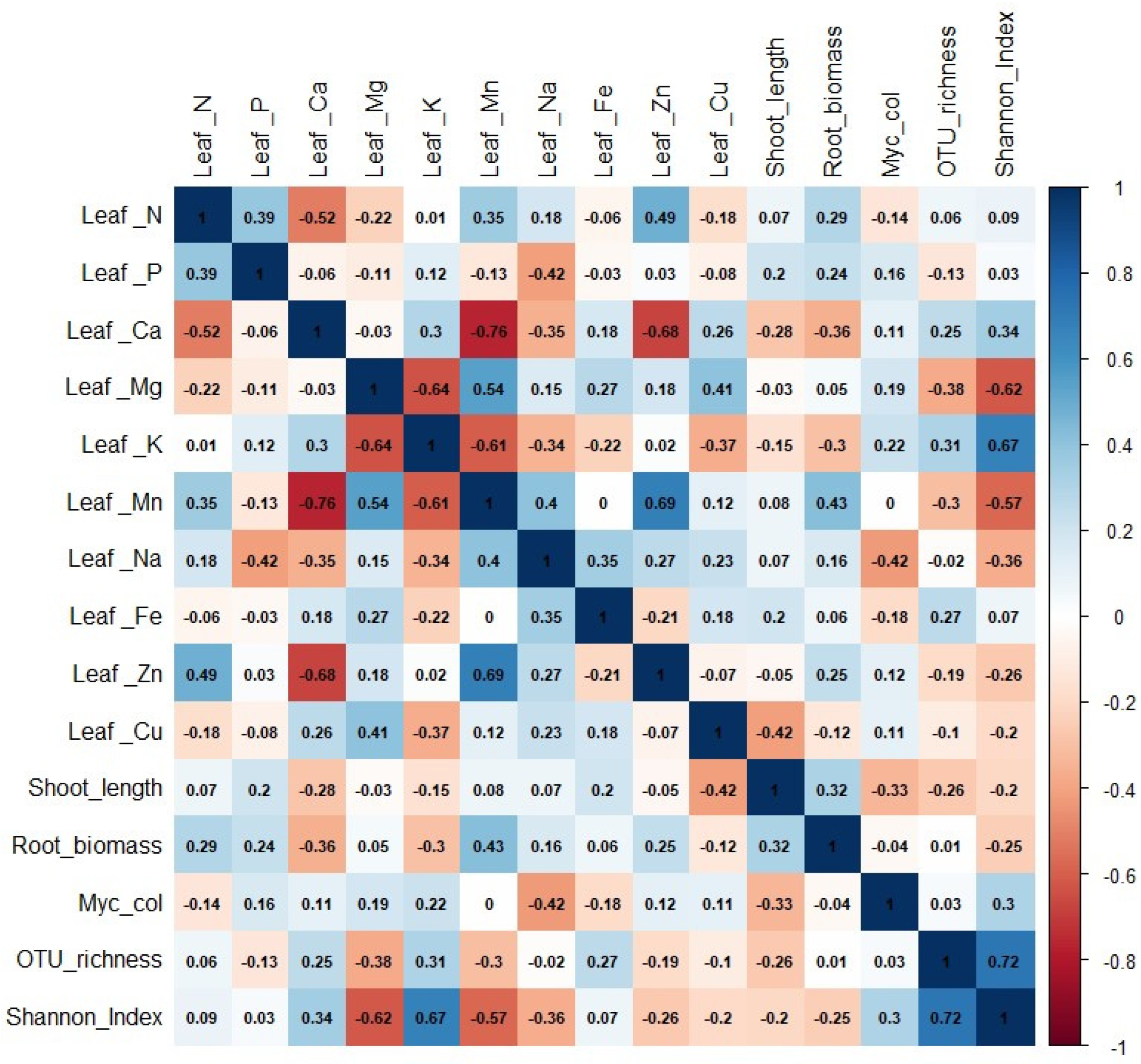
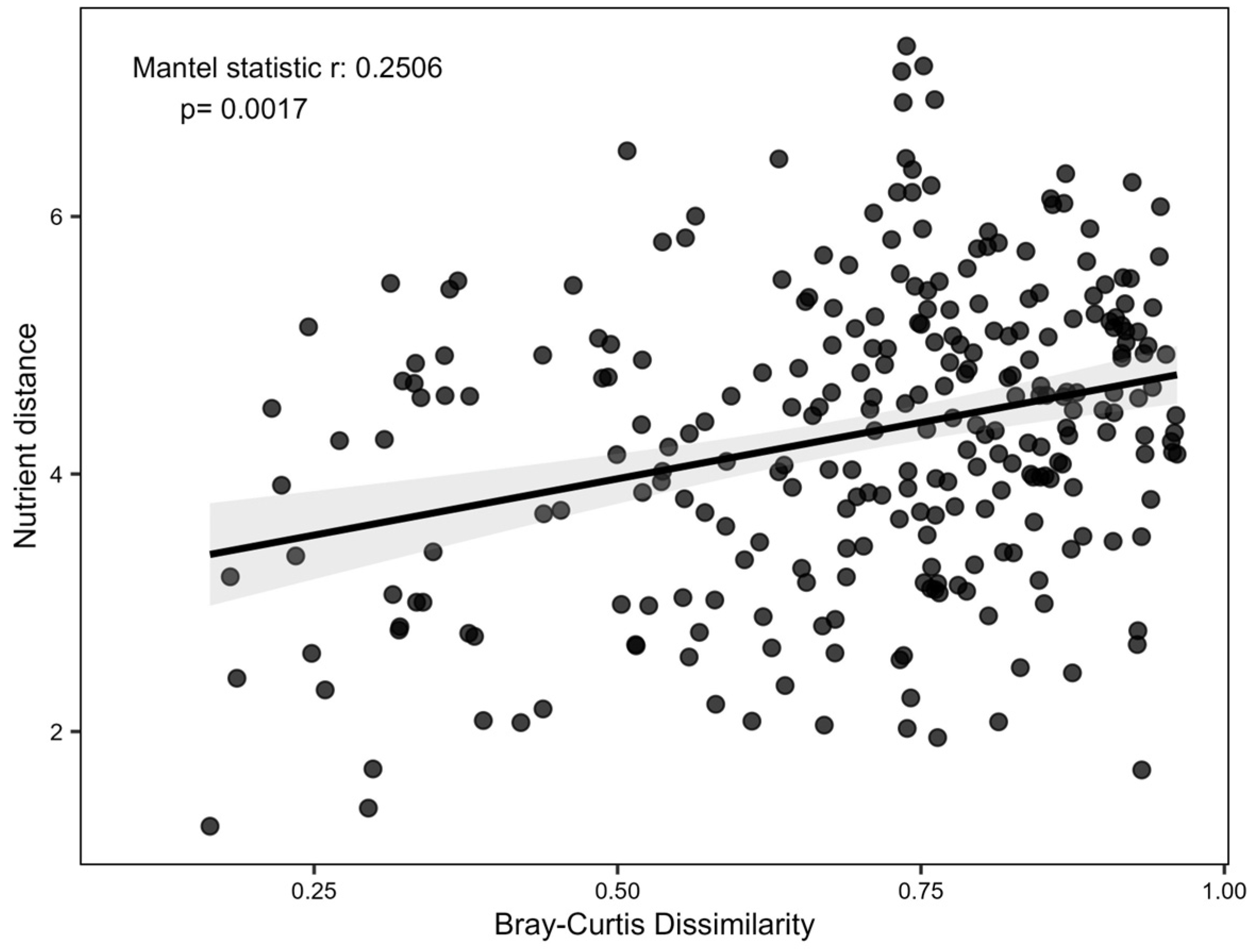
3.2. Effect of Different AMF Communities in the Growth and Nutrition of Grapevine Plants
4. Discussion
4.1. Land Use, Soil Type, and Rootstock Effects on Culturable AMF Communities
4.2. Effects of Different Root-Associated AMF Communities in Grapevine Growth and Nutrition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bardgett, R.D.; van der Putten, W.H. Belowground Biodiversity and Ecosystem Functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Zobel, M. How Mycorrhizal Associations Drive Plant Population and Community Biology. Science 2020, 367, eaba1223. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D. The Symbionts Forming Arbuscular Mycorrhizas. In Mycorrhizal Symbiosis; Elsevier: New York, NY, USA, 2008; pp. 13–41. ISBN 978-0-12-370526-6. [Google Scholar]
- Delavaux, C.S.; Smith-Ramesh, L.M.; Kuebbing, S.E. Beyond Nutrients: A Meta-Analysis of the Diverse Effects of Arbuscular Mycorrhizal Fungi on Plants and Soils. Ecology 2017, 98, 2111–2119. [Google Scholar] [CrossRef]
- Rasmann, S.; Bennett, A.; Biere, A.; Karley, A.; Guerrieri, E. Root Symbionts: Powerful Drivers of Plant above- and Belowground Indirect Defenses. Insect Sci. 2017, 24, 947–960. [Google Scholar] [CrossRef]
- Wilson, G.W.T.; Rice, C.W.; Rillig, M.C.; Springer, A.; Hartnett, D.C. Soil Aggregation and Carbon Sequestration Are Tightly Correlated with the Abundance of Arbuscular Mycorrhizal Fungi: Results from Long-Term Field Experiments. Ecol. Lett. 2009, 12, 452–461. [Google Scholar] [CrossRef]
- Bao, X.; Zou, J.; Zhang, B.; Wu, L.; Yang, T.; Huang, Q. Arbuscular Mycorrhizal Fungi and Microbes Interaction in Rice Mycorrhizosphere. Agronomy 2022, 12, 1277. [Google Scholar] [CrossRef]
- Priyadharsini, P.; Rojamala, K.; Ravi, R.K.; Muthuraja, R.; Nagaraj, K.; Muthukumar, T. Mycorrhizosphere: The Extended Rhizosphere and Its Significance. In Plant-Microbe Interaction: An Approach to Sustainable Agriculture; Choudhary, D.K., Varma, A., Tuteja, N., Eds.; Springer: Singapore, 2016; pp. 97–124. ISBN 978-981-10-2854-0. [Google Scholar]
- Linderman, R.G. Mycorrhizal Interactions with the Rhizosphere Microflora: The Mycorrhizosphere Effect. Phytopathology 1988, 78, 366–371. [Google Scholar]
- Berger, F.; Gutjahr, C. Factors Affecting Plant Responsiveness to Arbuscular Mycorrhiza. Curr. Opin. Plant Biol. 2021, 59, 101994. [Google Scholar] [CrossRef]
- Sawers, R.J.H.; Svane, S.F.; Quan, C.; Grønlund, M.; Wozniak, B.; Gebreselassie, M.-N.; González-Muñoz, E.; Chávez Montes, R.A.; Baxter, I.; Goudet, J.; et al. Phosphorus Acquisition Efficiency in Arbuscular Mycorrhizal Maize Is Correlated with the Abundance of Root-External Hyphae and the Accumulation of Transcripts Encoding PHT1 Phosphate Transporters. New Phytol. 2017, 214, 632–643. [Google Scholar] [CrossRef]
- Dodd, J.C. The Role of Arbuscular Mycorrhizal Fungi in Agro- and Natural Ecosystems. Outlook Agric. 2000, 29, 55–62. [Google Scholar] [CrossRef]
- Munkvold, L.; Kjøller, R.; Vestberg, M.; Rosendahl, S.; Jakobsen, I. High Functional Diversity within Species of Arbuscular Mycorrhizal Fungi. New Phytol. 2004, 164, 357–364. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.A.; Scheublin, T.R. Functional Traits in Mycorrhizal Ecology: Their Use for Predicting the Impact of Arbuscular Mycorrhizal Fungal Communities on Plant Growth and Ecosystem Functioning. New Phytol. 2007, 174, 244–250. [Google Scholar] [CrossRef]
- Marro, N.; Grilli, G.; Soteras, F.; Caccia, M.; Longo, S.; Cofré, N.; Borda, V.; Burni, M.; Janoušková, M.; Urcelay, C. The Effects of Arbuscular Mycorrhizal Fungal Species and Taxonomic Groups on Stressed and Unstressed Plants: A Global Meta-Analysis. New Phytol. 2022, 235, 320–332. [Google Scholar] [CrossRef]
- Avio, L.; Pellegrino, E.; Bonari, E.; Giovannetti, M. Functional Diversity of Arbuscular Mycorrhizal Fungal Isolates in Relation to Extraradical Mycelial Networks. New Phytol. 2006, 172, 347–357. [Google Scholar] [CrossRef]
- Hart, M.M.; Reader, R.J. Taxonomic Basis for Variation in the Colonization Strategy of Arbuscular Mycorrhizal Fungi. New Phytol. 2002, 153, 335–344. [Google Scholar] [CrossRef]
- Klironomos, J.N.; Hart, M.M. Colonization of Roots by Arbuscular Mycorrhizal Fungi Using Different Sources of Inoculum. Mycorrhiza 2002, 12, 181–184. [Google Scholar] [CrossRef]
- Fors, R.O.; Saggin Júnior, O.J.; Carneiro, M.A.C.; Berbara, R.L.L. Selection of Arbuscular Mycorrhizal Fungi for Sugarcane in Four Soils with the Presence of Dark Septate Endophytes. Acta Sci. Agron. 2019, 42, e42477. [Google Scholar] [CrossRef]
- Jeffries, P.; Gianinazzi, S.; Perotto, S.; Turnau, K.; Barea, J.-M. The Contribution of Arbuscular Mycorrhizal Fungi in Sustainable Maintenance of Plant Health and Soil Fertility. Biol. Fertil. Soils 2003, 37, 1–16. [Google Scholar] [CrossRef]
- Verbruggen, E.; Toby Kiers, E. Evolutionary Ecology of Mycorrhizal Functional Diversity in Agricultural Systems. Evol. Appl. 2010, 3, 547–560. [Google Scholar] [CrossRef]
- Tian, H.; Drijber, R.A.; Li, X.; Miller, D.N.; Wienhold, B.J. Arbuscular Mycorrhizal Fungi Differ in Their Ability to Regulate the Expression of Phosphate Transporters in Maize (Zea Mays L.). Mycorrhiza 2013, 23, 507–514. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.A.; Klironomos, J.N.; Ursic, M.; Moutoglis, P.; Streitwolf-Engel, R.; Boller, T.; Wiemken, A.; Sanders, I.R. Mycorrhizal Fungal Diversity Determines Plant Biodiversity, Ecosystem Variability and Productivity. Nature 1998, 396, 69–72. [Google Scholar] [CrossRef]
- Hart, M.M.; Reader, R.J.; Klironomos, J.N. Life-History Strategies of Arbuscular Mycorrhizal Fungi in Relation to Their Successional Dynamics. Mycologia 2001, 93, 1186–1194. [Google Scholar] [CrossRef]
- Cano, C.; Bago, A. Competition and Substrate Colonization Strategies of Three Polyxenically Grown Arbuscular Mycorrhizal Fungi. Mycologia 2005, 97, 1201–1214. [Google Scholar] [CrossRef]
- Engelmoer, D.J.P.; Behm, J.E.; Toby Kiers, E. Intense Competition between Arbuscular Mycorrhizal Mutualists in an in Vitro Root Microbiome Negatively Affects Total Fungal Abundance. Mol. Ecol. 2014, 23, 1584–1593. [Google Scholar] [CrossRef]
- Hepper, C.M.; Azcon-Aguilar, C.; Rosendahl, S.; Sen, R. Competition between Three Species of Glomus Used as Spatially Separated Introduced and Indigenous Mycorrhizal Inocula for Leek (Allium porrum L.). New Phytol. 1988, 110, 207–215. [Google Scholar] [CrossRef]
- Knegt, B.; Jansa, J.; Franken, O.; Engelmoer, D.J.P.; Werner, G.D.A.; Bücking, H.; Kiers, E.T. Host Plant Quality Mediates Competition between Arbuscular Mycorrhizal Fungi. Fungal Ecol. 2016, 20, 233–240. [Google Scholar] [CrossRef]
- Lu, N.; Zhou, X.; Cui, M.; Yu, M.; Zhou, J.; Qin, Y.; Li, Y. Colonization with Arbuscular Mycorrhizal Fungi Promotes the Growth of Morus Alba L. Seedlings under Greenhouse Conditions. Forests 2015, 6, 734–747. [Google Scholar] [CrossRef]
- Nogales, A.; Santos, E.S.; Abreu, M.M.; Arán, D.; Victorino, G.; Pereira, H.S.; Lopes, C.M.; Viegas, W. Mycorrhizal Inoculation Differentially Affects Grapevine’s Performance in Copper Contaminated and Non-Contaminated Soils. Front. Plant Sci. 2019, 9, 1906. [Google Scholar] [CrossRef]
- Thonar, C.; Frossard, E.; Šmilauer, P.; Jansa, J. Competition and Facilitation in Synthetic Communities of Arbuscular Mycorrhizal Fungi. Mol. Ecol. 2014, 23, 733–746. [Google Scholar] [CrossRef]
- Verbruggen, E.; van der Heijden, M.G.A.; Rillig, M.C.; Kiers, E.T. Mycorrhizal Fungal Establishment in Agricultural Soils: Factors Determining Inoculation Success. New Phytol. 2013, 197, 1104–1109. [Google Scholar] [CrossRef]
- Davison, J.; Moora, M.; Öpik, M.; Adholeya, A.; Ainsaar, L.; Bâ, A.; Burla, S.; Diedhiou, A.G.; Hiiesalu, I.; Jairus, T.; et al. Global Assessment of Arbuscular Mycorrhizal Fungus Diversity Reveals Very Low Endemism. Science 2015, 349, 970–973. [Google Scholar] [CrossRef]
- Edlinger, A.; Garland, G.; Hartman, K.; Banerjee, S.; Degrune, F.; García-Palacios, P.; Hallin, S.; Valzano-Held, A.; Herzog, C.; Jansa, J.; et al. Agricultural Management and Pesticide Use Reduce the Functioning of Beneficial Plant Symbionts. Nat. Ecol. Evol. 2022, 6, 1145–1154. [Google Scholar] [CrossRef]
- Riedo, J.; Wettstein, F.E.; Rösch, A.; Herzog, C.; Banerjee, S.; Büchi, L.; Charles, R.; Wächter, D.; Martin-Laurent, F.; Bucheli, T.D.; et al. Widespread Occurrence of Pesticides in Organically Managed Agricultural Soils—The Ghost of a Conventional Agricultural Past? Environ. Sci. Technol. 2021, 55, 2919–2928. [Google Scholar] [CrossRef]
- Helgason, T.; Daniell, T.J.; Husband, R.; Fitter, A.H.; Young, J.P.W. Ploughing up the Wood-Wide Web? Nature 1998, 394, 431. [Google Scholar] [CrossRef]
- Lumini, E.; Orgiazzi, A.; Borriello, R.; Bonfante, P.; Bianciotto, V. Disclosing Arbuscular Mycorrhizal Fungal Biodiversity in Soil through a Land-Use Gradient Using a Pyrosequencing Approach. Environ. Microbiol. 2010, 12, 2165–2179. [Google Scholar] [CrossRef]
- Menéndez, A.B.; Scervino, J.M.; Godeas, A.M. Arbuscular Mycorrhizal Populations Associated with Natural and Cultivated Vegetation on a Site of Buenos Aires Province, Argentina. Biol. Fertil. Soils 2001, 33, 373–381. [Google Scholar] [CrossRef]
- Oehl, F.; Sieverding, E.; Ineichen, K.; Mäder, P.; Boller, T.; Wiemken, A. Impact of Land Use Intensity on the Species Diversity of Arbuscular Mycorrhizal Fungi in Agroecosystems of Central Europe. Appl. Environ. Microbiol. 2003, 69, 2816–2824. [Google Scholar] [CrossRef]
- Brito, I.; Goss, M.J.; de Carvalho, M.; Chatagnier, O.; van Tuinen, D. Impact of Tillage System on Arbuscular Mycorrhiza Fungal Communities in the Soil under Mediterranean Conditions. Soil Tillage Res. 2012, 121, 63–67. [Google Scholar] [CrossRef]
- Bouffaud, M.-L.; Creamer, R.E.; Stone, D.; Plassart, P.; van Tuinen, D.; Lemanceau, P.; Wipf, D.; Redecker, D. Indicator Species and Co-Occurrence in Communities of Arbuscular Mycorrhizal Fungi at the European Scale. Soil Biol. Biochem. 2016, 103, 464–470. [Google Scholar] [CrossRef]
- Schnoor, T.K.; Lekberg, Y.; Rosendahl, S.; Olsson, P.A. Mechanical Soil Disturbance as a Determinant of Arbuscular Mycorrhizal Fungal Communities in Semi-Natural Grassland. Mycorrhiza 2011, 21, 211–220. [Google Scholar] [CrossRef]
- Van Geel, M.; Verbruggen, E.; De Beenhouwer, M.; van Rennes, G.; Lievens, B.; Honnay, O. High Soil Phosphorus Levels Overrule the Potential Benefits of Organic Farming on Arbuscular Mycorrhizal Diversity in Northern Vineyards. Agric. Ecosyst. Environ. 2017, 248, 144–152. [Google Scholar] [CrossRef]
- Banerjee, S.; van der Heijden, M.G.A. Soil Microbiomes and One Health. Nat. Rev. Microbiol. 2023, 21, 6–20. [Google Scholar] [CrossRef]
- OIV. State of the World Vine and Wine Sector 2021; International Organisation of Vine and Wine: Paris, France, 2022; p. 19. [Google Scholar]
- FAO. Crops and Livestock Products. Area Harvested in 2021. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 4 January 2023).
- Trouvelot, S.; Bonneau, L.; Redecker, D.; van Tuinen, D.; Adrian, M.; Wipf, D. Arbuscular Mycorrhiza Symbiosis in Viticulture: A Review. Agron. Sustain. Dev. 2015, 35, 1449–1467. [Google Scholar] [CrossRef]
- Betancur-Agudelo, M.; Meyer, E.; Lovato, P.E. Increased Copper Concentrations in Soil Affect Indigenous Arbuscular Mycorrhizal Fungi and Physiology of Grapevine Plantlets. Rhizosphere 2023, 27, 100711. [Google Scholar] [CrossRef]
- Ambrosini, V.G.; Voges, J.G.; Canton, L.; Couto, R.d.R.; Ferreira, P.A.A.; Comin, J.J.; de Melo, G.W.B.; Brunetto, G.; Soares, C.R.F.S. Effect of Arbuscular Mycorrhizal Fungi on Young Vines in Copper-Contaminated Soil. Braz. J. Microbiol. 2015, 46, 1045–1052. [Google Scholar] [CrossRef]
- Rosa, D.J.; Ambrosini, V.G.; Brunetto, G.; Soares, C.R.F.S.; Borghezan, M.; Pescador, R. Parâmetros fisiológicos em videiras ‘Paulsen 1103’ (Vitis berlandieri x Vitis rupestris) inoculadas com fungos micorrízicos arbusculares em solo contaminado com cobre. Ciência Téc. Vitiv. 2016, 31, 14–23. [Google Scholar] [CrossRef]
- Aguilera, P.; Ortiz, N.; Becerra, N.; Turrini, A.; Gaínza-Cortés, F.; Silva-Flores, P.; Aguilar-Paredes, A.; Romero, J.K.; Jorquera-Fontena, E.; de La Luz Mora, M.; et al. Application of Arbuscular Mycorrhizal Fungi in Vineyards: Water and Biotic Stress Under a Climate Change Scenario: New Challenge for Chilean Grapevine Crop. Front. Microbiol. 2022, 13, 826571. [Google Scholar] [CrossRef]
- IPMA. Ficha Climatológica Lisboa 1971–2000. Available online: https://www.ipma.pt/pt/oclima/normais.clima/1971-2000/normalclimate7100.jsp (accessed on 5 January 2023).
- IUSS Working Group. WRB World Reference Base for Soil Resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, 4th ed.; International Union of Soil Sciences (IUSS): Vienna, Austria, 2022. [Google Scholar]
- Porter, W.M. The “most Probable Number” Method for Enumerating Infective Propagules of Vesicular Arbuscular Mycorrhizal Fungi in Soil. Soil Res. 1979, 17, 515–519. [Google Scholar] [CrossRef]
- Powell, C.L. Mycorrhizal Infectivity of Eroded Soils. Soil Biol. Biochem. 1980, 12, 247–250. [Google Scholar] [CrossRef]
- Koske, R.E.; Gemma, J.N. A Modified Procedure for Staining Roots to Detect VA Mycorrhizas. Mycol. Res. 1989, 92, 486–488. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved Procedures for Clearing Roots and Staining Parasitic and Vesicular-Arbuscular Mycorrhizal Fungi for Rapid Assessment of Infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Jarvis, B.; Wilrich, C.; Wilrich, P.-T. Reconsideration of the Derivation of Most Probable Numbers, Their Standard Deviations, Confidence Bounds and Rarity Values. J. Appl. Microbiol. 2010, 109, 1660–1667. [Google Scholar] [CrossRef]
- Nogales, A.; Rottier, E.; Campos, C.; Victorino, G.; Costa, J.M.; Coito, J.L.; Pereira, H.S.; Viegas, W.; Lopes, C. The Effects of Field Inoculation of Arbuscular Mycorrhizal Fungi through Rye Donor Plants on Grapevine Performance and Soil Properties. Agric. Ecosyst. Environ. 2021, 313, 107369. [Google Scholar] [CrossRef]
- Mottard, G.; Nespoulous, J.; Marcout, P. Les portes-greffes de la vigne: Caractères distinctifs, aptitudes culturales. Bull. Tech. D’inf. Ingénieurs Serv. Agric. 1963, 182, 36. [Google Scholar]
- Infovini. Available online: http://www.infovini.com/classic/pagina.php?codPagina=52&codItem=118 (accessed on 15 June 2023).
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil. Circular. Calif. Agric. Exp. Stn. 1950, 347, 32. [Google Scholar]
- Giovannetti, M.; Mosse, B. An Evaluation of Techniques for Measuring Vesicular Arbuscular Mycorrhizal Infection in Roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Gollotte, A.; van Tuinen, D.; Atkinson, D. Diversity of Arbuscular Mycorrhizal Fungi Colonising Roots of the Grass Species Agrostis Capillaris and Lolium Perenne in a Field Experiment. Mycorrhiza 2004, 14, 111–117. [Google Scholar] [CrossRef]
- Öpik, M.; Vanatoa, A.; Vanatoa, E.; Moora, M.; Davison, J.; Kalwij, J.M.; Reier, Ü.; Zobel, M. The Online Database MaarjAM Reveals Global and Ecosystemic Distribution Patterns in Arbuscular Mycorrhizal Fungi (Glomeromycota). New Phytol. 2010, 188, 223–241. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.6-4. 2019. Available online: https://cran.r-hub.io/web/packages/vegan/vegan.pdf (accessed on 10 April 2023).
- Oliveros, J.C. Venny. An Interactive Tool for Comparing Lists with Venn’s Diagrams. Available online: https://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 15 June 2023).
- Lu, Y.; Zhou, G.; Ewald, J.; Pang, Z.; Shiri, T.; Xia, J. MicrobiomeAnalyst 2.0: Comprehensive Statistical, Functional and Integrative Analysis of Microbiome Data. Nucleic Acids Res. 2023, 51, W310–W318. [Google Scholar] [CrossRef]
- Barberán, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using Network Analysis to Explore Co-Occurrence Patterns in Soil Microbial Communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef]
- Guo, X.; Wang, P.; Wang, X.; Li, Y.; Ji, B. Specific Plant Mycorrhizal Responses Are Linked to Mycorrhizal Fungal Species Interactions. Front. Plant Sci. 2022, 13, 930069. [Google Scholar]
- Griffith, D.M.; Veech, J.A.; Marsh, C.J. Cooccur: Probabilistic Species Co-Occurrence Analysis in R. J. Stat. Softw. 2016, 69, 1–17. [Google Scholar] [CrossRef]
- Almende, B.V.; Benoit, T.; Titouan, R. Package ‘visNetwork’. Network Visualization Using ‘Vis.Js’. Version 2.1.9. 2019. Available online: https://cran.r-project.org/web/packages/visNetwork/visNetwork.pdf (accessed on 10 April 2023).
- Anderson, M.J.; Walsh, D.C.I. PERMANOVA, ANOSIM, and the Mantel Test in the Face of Heterogeneous Dispersions: What Null Hypothesis Are You Testing? Ecol. Monogr. 2013, 83, 557–574. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2; Use R! Springer International Publishing: Cham, Switzerland, 2016; ISBN 978-3-319-24275-0. [Google Scholar]
- Stürmer, S.L.; Bever, J.D.; Morton, J.B. Biogeography of Arbuscular Mycorrhizal Fungi (Glomeromycota): A Phylogenetic Perspective on Species Distribution Patterns. Mycorrhiza 2018, 28, 587–603. [Google Scholar] [CrossRef]
- Balestrini, R.; Magurno, F.; Walker, C.; Lumini, E.; Bianciotto, V. Cohorts of Arbuscular Mycorrhizal Fungi (AMF) in Vitis Vinifera, a Typical Mediterranean Fruit Crop: Cohorts of AM Fungi in Vineyards. Environ. Microbiol. Rep. 2010, 2, 594–604. [Google Scholar] [CrossRef]
- Schreiner, R.P.; Mihara, K.L. The Diversity of Arbuscular Mycorrhizal Fungi Amplified from Grapevine Roots (Vitis Vinifera L.) in Oregon Vineyards Is Seasonally Stable and Influenced by Soil and Vine Age. Mycologia 2009, 101, 599–611. [Google Scholar] [CrossRef]
- Sýkorová, Z.; Ineichen, K.; Wiemken, A.; Redecker, D. The Cultivation Bias: Different Communities of Arbuscular Mycorrhizal Fungi Detected in Roots from the Field, from Bait Plants Transplanted to the Field, and from a Greenhouse Trap Experiment. Mycorrhiza 2007, 18, 1–14. [Google Scholar] [CrossRef]
- Chagnon, P.-L.; Bradley, R.L.; Maherali, H.; Klironomos, J.N. A Trait-Based Framework to Understand Life History of Mycorrhizal Fungi. Trends Plant Sci. 2013, 18, 484–491. [Google Scholar] [CrossRef]
- Jackson, R.S. Wine Science Principles and Applications, 4th ed.; Academic Press: Cambridge, MA, USA, 2014; ISBN 978-0-12-381469-2. [Google Scholar]
- Moukarzel, R.; Ridgway, H.J.; Guerin-Laguette, A.; Jones, E.E. Grapevine Rootstocks Drive the Community Structure of Arbuscular Mycorrhizal Fungi in New Zealand Vineyards. J. Appl. Microbiol. 2021, 131, 2941–2956. [Google Scholar] [CrossRef]
- Likar, M.; Hančević, K.; Radić, T.; Regvar, M. Distribution and Diversity of Arbuscular Mycorrhizal Fungi in Grapevines from Production Vineyards along the Eastern Adriatic Coast. Mycorrhiza 2013, 23, 209–219. [Google Scholar] [CrossRef]
- Vieira, L.C.; da Silva, D.K.A.; Escobar, I.E.C.; da Silva, J.M.; de Moura, I.A.; Oehl, F.; da Silva, G.A. Changes in an Arbuscular Mycorrhizal Fungi Community Along an Environmental Gradient. Plants 2020, 9, 52. [Google Scholar] [CrossRef]
- Maherali, H.; Klironomos, J.N. Influence of Phylogeny on Fungal Community Assembly and Ecosystem Functioning. Science 2007, 316, 1746–1748. [Google Scholar] [CrossRef]
- Breuillin, F.; Schramm, J.; Hajirezaei, M.; Ahkami, A.; Favre, P.; Druege, U.; Hause, B.; Bucher, M.; Kretzschmar, T.; Bossolini, E.; et al. Phosphate Systemically Inhibits Development of Arbuscular Mycorrhiza in Petunia Hybrida and Represses Genes Involved in Mycorrhizal Functioning. Plant J. 2010, 64, 1002–1017. [Google Scholar] [CrossRef]
- Grant, C.; Bittman, S.; Montreal, M.; Plenchette, C.; Morel, C. Soil and Fertilizer Phosphorus: Effects on Plant P Supply and Mycorrhizal Development. Can. J. Plant Sci. 2005, 85, 3–14. [Google Scholar] [CrossRef]
- Nouri, E.; Breuillin-Sessoms, F.; Feller, U.; Reinhardt, D. Phosphorus and Nitrogen Regulate Arbuscular Mycorrhizal Symbiosis in Petunia Hybrida. PLoS ONE 2014, 9, e90841. [Google Scholar] [CrossRef]
- Treseder, K.K.; Allen, M.F. Direct Nitrogen and Phosphorus Limitation of Arbuscular Mycorrhizal Fungi: A Model and Field Test. New Phytol. 2002, 155, 507–515. [Google Scholar] [CrossRef]
- Johnson, D.; IJdo, M.; Genney, D.R.; Anderson, I.C.; Alexander, I.J. How Do Plants Regulate the Function, Community Structure, and Diversity of Mycorrhizal Fungi? J. Exp. Bot. 2005, 56, 1751–1760. [Google Scholar] [CrossRef]
- Zhang, J.; Quan, C.; Ma, L.; Chu, G.; Liu, Z.; Tang, X. Plant Community and Soil Properties Drive Arbuscular Mycorrhizal Fungal Diversity: A Case Study in Tropical Forests. Soil Ecol. Lett. 2021, 3, 52–62. [Google Scholar] [CrossRef]
- Guzman, A.; Montes, M.; Hutchins, L.; DeLaCerda, G.; Yang, P.; Kakouridis, A.; Dahlquist-Willard, R.M.; Firestone, M.K.; Bowles, T.; Kremen, C. Crop Diversity Enriches Arbuscular Mycorrhizal Fungal Communities in an Intensive Agricultural Landscape. New Phytol. 2021, 231, 447–459. [Google Scholar] [CrossRef]
- Karagiannidis, N.; Nikolaou, N. Arbuscular Root Infection as an Important Factor of Grapevine Nutrition Status: Multivariate Analysis Application for Evaluation and Characterization of the Soil and Leaf. Agrochimica 1999, 43, 151–168. [Google Scholar]
- Nikolaou, N.; Karagiannidis, N.; Koundouras, S.; Fysarakis, I. Effects of Different P-Sources in Soil on Increasing Growth and Mineral Uptake of Mycorrhizal Vitis Vinifera L. (Cv Victoria) Vines. OENO One 2002, 36, 195–204. [Google Scholar] [CrossRef]
- Hage-Ahmed, K.; Rosner, K.; Steinkellner, S. Arbuscular Mycorrhizal Fungi and Their Response to Pesticides. Pest Manag. Sci. 2019, 75, 583–590. [Google Scholar] [CrossRef]
- Lori, M.; Symnaczik, S.; Mäder, P.; Deyn, G.D.; Gattinger, A. Organic Farming Enhances Soil Microbial Abundance and Activity—A Meta-Analysis and Meta-Regression. PLoS ONE 2017, 12, e0180442. [Google Scholar] [CrossRef]
- Martínez-García, L.B.; Korthals, G.; Brussaard, L.; Jørgensen, H.B.; De Deyn, G.B. Organic Management and Cover Crop Species Steer Soil Microbial Community Structure and Functionality along with Soil Organic Matter Properties. Agric. Ecosyst. Environ. 2018, 263, 7–17. [Google Scholar] [CrossRef]
- Banerjee, S.; Walder, F.; Büchi, L.; Meyer, M.; Held, A.Y.; Gattinger, A.; Keller, T.; Charles, R.; van der Heijden, M.G.A. Agricultural Intensification Reduces Microbial Network Complexity and the Abundance of Keystone Taxa in Roots. ISME J. 2019, 13, 1722–1736. [Google Scholar] [CrossRef]
- Zaller, J.G.; Cantelmo, C.; Santos, G.D.; Muther, S.; Gruber, E.; Pallua, P.; Mandl, K.; Friedrich, B.; Hofstetter, I.; Schmuckenschlager, B.; et al. Herbicides in Vineyards Reduce Grapevine Root Mycorrhization and Alter Soil Microorganisms and the Nutrient Composition in Grapevine Roots, Leaves, Xylem Sap and Grape Juice. Environ. Sci. Pollut. Res. 2018, 25, 23215–23226. [Google Scholar] [CrossRef]
- Zaller, J.G.; Heigl, F.; Ruess, L.; Grabmaier, A. Glyphosate Herbicide Affects Belowground Interactions between Earthworms and Symbiotic Mycorrhizal Fungi in a Model Ecosystem. Sci. Rep. 2014, 4, 5634. [Google Scholar] [CrossRef]
- Jansa, J.; Mozafar, A.; Anken, T.; Ruh, R.; Sanders, I.; Frossard, E. Diversity and Structure of AMF Communities as Affected by Tillage in a Temperate Soil. Mycorrhiza 2002, 12, 225–234. [Google Scholar] [CrossRef]
- Bainard, L.D.; Dai, M.; Gomez, E.F.; Torres-Arias, Y.; Bainard, J.D.; Sheng, M.; Eilers, W.; Hamel, C. Arbuscular Mycorrhizal Fungal Communities Are Influenced by Agricultural Land Use and Not Soil Type among the Chernozem Great Groups of the Canadian Prairies. Plant Soil 2015, 387, 351–362. [Google Scholar] [CrossRef]
- Darriaut, R.; Antonielli, L.; Martins, G.; Ballestra, P.; Vivin, P.; Marguerit, E.; Mitter, B.; Masneuf-Pomarède, I.; Compant, S.; Ollat, N.; et al. Soil Composition and Rootstock Genotype Drive the Root Associated Microbial Communities in Young Grapevines. Front. Microbiol. 2022, 13, 1031064. [Google Scholar] [CrossRef]
- Moukarzel, R.; Ridgway, H.J.; Waller, L.; Guerin-Laguette, A.; Cripps-Guazzone, N.; Jones, E.E. Soil Arbuscular Mycorrhizal Fungal Communities Differentially Affect Growth and Nutrient Uptake by Grapevine Rootstocks. Microb. Ecol. 2022, 86, 1035–1049. [Google Scholar] [CrossRef]
- Layeghifard, M.; Hwang, D.M.; Guttman, D.S. Disentangling Interactions in the Microbiome: A Network Perspective. Trends Microbiol. 2017, 25, 217–228. [Google Scholar] [CrossRef]
- Berry, D.; Widder, S. Deciphering Microbial Interactions and Detecting Keystone Species with Co-Occurrence Networks. Front. Microbiol. 2014, 5, 219. [Google Scholar]
- Fox, A.; Widmer, F.; Lüscher, A. Soil Microbial Community Structures Are Shaped by Agricultural Systems Revealing Little Temporal Variation. Environ. Res. 2022, 214, 113915. [Google Scholar] [CrossRef]
- Torres, N.; Yu, R.; Kurtural, S.K. Arbuscular Mycorrhizal Fungi Inoculation and Applied Water Amounts Modulate the Response of Young Grapevines to Mild Water Stress in a Hyper-Arid Season. Front. Plant Sci. 2021, 11, 622209. [Google Scholar]
- Hartman, K.; van der Heijden, M.G.A.; Wittwer, R.A.; Banerjee, S.; Walser, J.-C.; Schlaeppi, K. Cropping Practices Manipulate Abundance Patterns of Root and Soil Microbiome Members Paving the Way to Smart Farming. Microbiome 2018, 6, 14. [Google Scholar] [CrossRef]
- Michalska-Smith, M.; Song, Z.; Spawn-Lee, S.A.; Hansen, Z.A.; Johnson, M.; May, G.; Borer, E.T.; Seabloom, E.W.; Kinkel, L.L. Network Structure of Resource Use and Niche Overlap within the Endophytic Microbiome. ISME J. 2022, 16, 435–446. [Google Scholar] [CrossRef]
- Xiong, X.; Wang, W.; Xing, Y.; Chen, H.; Luo, X.; Chen, W.; Huang, Q. Niche Overlap Is a Predictor of the Interspecies Correlations Detected by Microbial Network Analysis in Soil Micro-Aggregates. J. Soils Sediments 2022, 22, 1521–1529. [Google Scholar] [CrossRef]
- Yang, Y.; Shi, Y.; Fang, J.; Chu, H.; Adams, J.M. Soil Microbial Network Complexity Varies With pH as a Continuum, Not a Threshold, Across the North China Plain. Front. Microbiol. 2022, 13, 895687. [Google Scholar]
- Marasco, R.; Rolli, E.; Fusi, M.; Michoud, G.; Daffonchio, D. Grapevine Rootstocks Shape Underground Bacterial Microbiome and Networking but Not Potential Functionality. Microbiome 2018, 6, 3. [Google Scholar] [CrossRef]
- Brown, S.P.; Grillo, M.A.; Podowski, J.C.; Heath, K.D. Soil Origin and Plant Genotype Structure Distinct Microbiome Compartments in the Model Legume Medicago Truncatula. Microbiome 2020, 8, 139. [Google Scholar] [CrossRef]
- Liu, J.; Abdelfattah, A.; Wasserman, B.; Wisniewski, M.; Droby, S.; Fazio, G.; Mazzola, M.; Wu, X. Contrasting Effects of Genotype and Root Size on the Fungal and Bacterial Communities Associated with Apple Rootstocks. Hortic. Res. 2022, 9, uhab013. [Google Scholar] [CrossRef]
- Karagiannidis, N.; Velemis, D.; Stavropoulos, N. Root Colonization and Spore Population by VA-Mycorrhizal Fungi in Four Grapevine Rootstocks. VITIS J. Grapevine Res. 1997, 36, 57. [Google Scholar] [CrossRef]
- Schreiner, R.P. Mycorrhizal Colonization of Grapevine Rootstocks under Field Conditions. Am. J. Enol. Vitic. 2003, 54, 143–149. [Google Scholar]
- Druille, M.; Omacini, M.; Golluscio, R.A.; Cabello, M.N. Arbuscular Mycorrhizal Fungi Are Directly and Indirectly Affected by Glyphosate Application. Appl. Soil Ecol. 2013, 72, 143–149. [Google Scholar] [CrossRef]
- Helander, M.; Saloniemi, I.; Omacini, M.; Druille, M.; Salminen, J.-P.; Saikkonen, K. Glyphosate Decreases Mycorrhizal Colonization and Affects Plant-Soil Feedback. Sci. Total Environ. 2018, 642, 285–291. [Google Scholar] [CrossRef]
- Weber, S.E.; Diez, J.M.; Andrews, L.V.; Goulden, M.L.; Aronson, E.L.; Allen, M.F. Responses of Arbuscular Mycorrhizal Fungi to Multiple Coinciding Global Change Drivers. Fungal Ecol. 2019, 40, 62–71. [Google Scholar] [CrossRef]
- Elumeeva, T.G.; Onipchenko, V.G.; Cornelissen, J.H.C.; Semenova, G.V.; Perevedentseva, L.G.; Freschet, G.T.; van Logtestijn, R.S.P.; Soudzilovskaia, N.A. Is Intensity of Plant Root Mycorrhizal Colonization a Good Proxy for Plant Growth Rate, Dominance and Decomposition in Nutrient Poor Conditions? J. Veg. Sci. 2018, 29, 715–725. [Google Scholar] [CrossRef]
- Schreiner, R.P. Mycorrhizas and Mineral Acquisition in Grapevines. In Proceedings of the Soil Environment and Vine Mineral Nutrition Symposium, San Diego, CA, USA, 29–30 June 2004. [Google Scholar]
- Clark, R.B.; Zeto, S.K. Mineral Acquisition by Arbuscular Mycorrhizal Plants. J. Plant Nutr. 2000, 23, 867–902. [Google Scholar] [CrossRef]
- Marschner, H. Functions of Mineral Nutrients: Micronutrients. In Mineral Nutrition of Higher Plants; Academic Press: London, UK, 1995; pp. 313–404. [Google Scholar]
- Khalil, H.A. Influence of Vesicular-Arbuscula Mycorrhizal Fungi (Glomus Spp.) on the Response of Grapevines Rootstocks to Salt Stress. Asian J. Crop Sci. 2013, 5, 393–404. [Google Scholar] [CrossRef]
- Biricolti, S.; Ferrini, F.; Rinaldelli, E.; Tamantini, I.; Vignozzi, N. VAM Fungi and Soil Lime Content Influence Rootstock Growth and Nutrient Content. Am. J. Enol. Vitic. 1997, 48, 93–99. [Google Scholar] [CrossRef]
- Mortimer, P.E.; Archer, E.; Valentine, A.J. Mycorrhizal C Costs and Nutritional Benefits in Developing Grapevines. Mycorrhiza 2005, 15, 159–165. [Google Scholar] [CrossRef]
- Howeler, R.H.; Cadavid, L.F.; Burckhardt, E. Response of Cassava to VA Mycorrhizal Inoculation and Phosphorus Application in Greenhouse and Field Experiments. Plant Soil 1982, 69, 327–339. [Google Scholar] [CrossRef]
- Ceballos, I.; Ruiz, M.; Fernández, C.; Peña, R.; Rodríguez, A.; Sanders, I.R. The In Vitro Mass-Produced Model Mycorrhizal Fungus, Rhizophagus irregularis, Significantly Increases Yields of the Globally Important Food Security Crop Cassava. PLoS ONE 2013, 8, e70633. [Google Scholar] [CrossRef]
- Heberle, E.d.S.; de Armas, R.D.; Heberle, D.A.; Stürmer, S.L.; Peruch, L.A.M.; Lovato, P.E.; Soares, C.R.F.S. Occurrence and Structure of Arbuscular Mycorrhizal Fungal Communities in Cassava after Cultivation of Cover Crops as Observed by the “PCR-DGGE” Technique. Rev. Bras. Ciênc. Solo 2015, 39, 1292–1301. [Google Scholar] [CrossRef]
- Cavallari, L.G.; Fernandes, A.M.; da Silva de Oliveira Mota, L.H.; Leite, H.M.F.; Piroli, V.L.B. Growth and Phosphorus Uptake by Cassava in P-Deficient Soil in Response to Mycorrhizal Inoculation. Rev. Bras. Ciênc. Solo 2021, 45, e0210060. [Google Scholar] [CrossRef]
- Yang, G.; Liu, N.; Lu, W.; Wang, S.; Kan, H.; Zhang, Y.; Xu, L.; Chen, Y. The Interaction between Arbuscular Mycorrhizal Fungi and Soil Phosphorus Availability Influences Plant Community Productivity and Ecosystem Stability. J. Ecol. 2014, 102, 1072–1082. [Google Scholar] [CrossRef]
- Garcia, K.; Zimmermann, S.D. The Role of Mycorrhizal Associations in Plant Potassium Nutrition. Front. Plant Sci. 2014, 5, 337. [Google Scholar] [CrossRef]
- Lehmann, A.; Rillig, M.C. Arbuscular Mycorrhizal Contribution to Copper, Manganese and Iron Nutrient Concentrations in Crops—A Meta-Analysis. Soil Biol. Biochem. 2015, 81, 147–158. [Google Scholar] [CrossRef]
- Faria, J.M.S.; Teixeira, D.M.; Ferreira, D.; Barrulas, P.; Brito, I.; Pinto, A.P.; Carvalho, M. Manganese Uptake to Wheat Shoot Meristems Is Differentially Influenced by Arbuscular Mycorrhiza Fungal Communities Adapted to Acidic Soil. Soil Syst. 2022, 6, 50. [Google Scholar] [CrossRef]
- Baslam, M.; Garmendia, I.; Goicoechea, N. Elevated CO2 May Impair the Beneficial Effect of Arbuscular Mycorrhizal Fungi on the Mineral and Phytochemical Quality of Lettuce. Ann. Appl. Biol. 2012, 161, 180–191. [Google Scholar] [CrossRef]
- Xie, K.; Cakmak, I.; Wang, S.; Zhang, F.; Guo, S. Synergistic and Antagonistic Interactions between Potassium and Magnesium in Higher Plants. Crop. J. 2021, 9, 249–256. [Google Scholar] [CrossRef]
- Aslanpour, M.; Baneh, H.D.; Tehranifar, A.; Shoor, M. Effect of Mycorrhizal Fungi on Macronutrients and Micronutrients in the White Seedless Grape Roots under the Drought Conditions. Int. Trans. J. Eng. Manag. Appl. Sci. Technol. 2019, 10, 397–408. [Google Scholar]
- Von Bennewitz Alvarez, E. Effects of the Inoculation with Arbuscular Mycorrhizal (AM) Fungus of the Genus Glomus on Growth and Leaf Mineral Concentrations of Grapevine (Vitis Vinifera Cv. Cabernet Sauvignon); University of Hohenheim: Hohenheim, Germany, 2007. [Google Scholar]
- Petgen, M.; Schropp, A.; George, E.; Römheld, V. Einfluss Unterschiedlicher Inokulationstiefen Mit Dem Arbuskulären Mykorrhizapilz Glomus mosseae Auf Die Mykorrhizierung Bei Reben (Vitis sp.) in Wurzelbeobachtungskästen. VITIS J. Grapevine Res. 1998, 37, 99–105. [Google Scholar]
- Mensah, J.A.; Koch, A.M.; Antunes, P.M.; Kiers, E.T.; Hart, M.; Bücking, H. High Functional Diversity within Species of Arbuscular Mycorrhizal Fungi Is Associated with Differences in Phosphate and Nitrogen Uptake and Fungal Phosphate Metabolism. Mycorrhiza 2015, 25, 533–546. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Lisbon | Pegões | ||
|---|---|---|---|---|
| NAS | VS | NAS | VS | |
| Coarse soil fraction % (>2 mm) | 34.2 | 34.4 | 8.7 | 10.7 |
| Fine soil fraction % (<2 mm) | 65.8 | 65.6 | 91.3 | 89.3 |
| pH | 7.9 | 7.5 | 5.4 | 6.9 |
| Electric conductivity 1 | 150.10 | 137.00 | 28.15 | 36.89 |
| Cation exchange capacity 2 | 54.4 | 60.2 | 3.1 | 3.6 |
| Total organic carbon 3 | 27.78 | 17.57 | 10.98 | 4.72 |
| Total N 3 | 2.36 | 1.33 | 0.46 | 0.30 |
| Macronutrient concentration in available fraction | ||||
| P 4 | 18.61 | 42.9 | 4.34 | 14.99 |
| Ca 3 | 16.1 | 13.9 | 9.4 | 9.8 |
| Na 3 | 2.9 | 3.1 | 2.5 | 3.0 |
| K 4 | 284.9 | 179.9 | 27.4 | 64.2 |
| Mg 4 | 632.2 | 1229.0 | 26.1 | 36.5 |
| Micronutrient concentration in available fraction | ||||
| Fe 4 | 59.12 | 132.87 | 106.42 | 56.57 |
| Mn 4 | 29.41 | 45.89 | 1.52 | 2.40 |
| Zn 4 | 14.39 | 5.1 | 2.30 | 3.34 |
| Cu 4 | 4.00 | 6.97 | 0.02 | 13.98 |
| Factors/Effects | OTU Richness | Shannon Index |
|---|---|---|
| Rootstock | 0.496 | 0.139 |
| Soil type | 0.007 | <0.001 |
| Land use | 0.192 | 0.227 |
| Rootstock × Soil type | 0.515 | 0.685 |
| Rootstock × Land use | 0.347 | 0.994 |
| Soil type × Land use | 0.882 | 0.073 |
| Rootstock × Soil type × Land use | 0.267 | 0.658 |
| Factor/Effect | F | p-Value |
|---|---|---|
| Rootstock | 1.3517 | 0.224 |
| Land use | 5.7826 | 0.001 |
| Soil type | 7.9101 | 0.001 |
| Rootstock × Land use | 1.7479 | 0.121 |
| Rootstock × Soil type | 0.9196 | 0.487 |
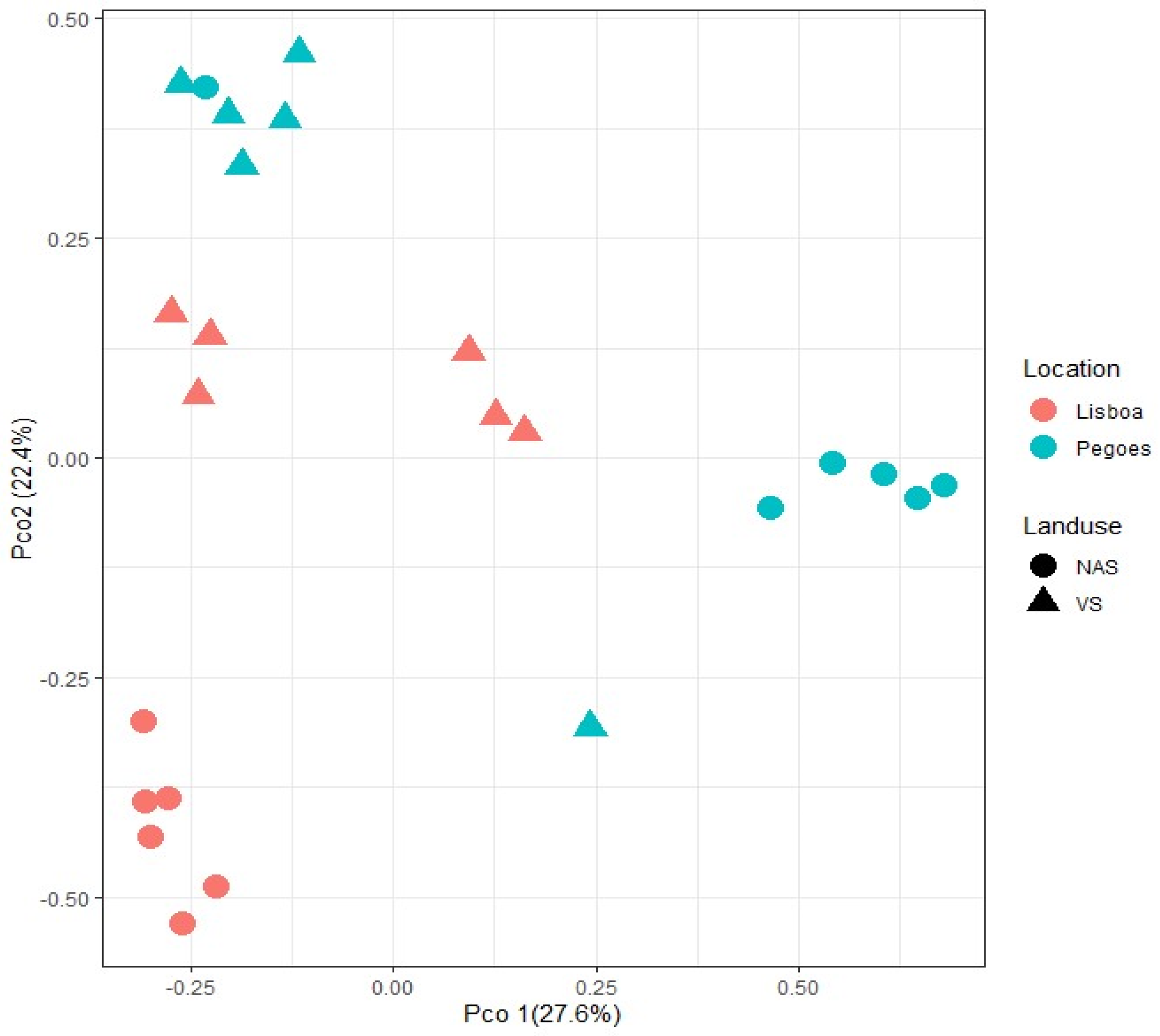
| Land use × Soil type | 6.4062 | 0.001 |
| Rootstock × Land use × Soil type | 1.3307 | 0.246 |
| Experimental Groups | Positive Connections | Negative Connections | Total Significant Connections | Total Analyzed Pairs | Percentage of Non-Random |
|---|---|---|---|---|---|
| Soil type | |||||
| Lisbon | 345 | 273 | 618 | 21,000 | 2.9 |
| Pegões | 725 | 67 | 792 | 16,735 | 4.7 |
| Land use | |||||
| Vineyard soil | 725 | 67 | 792 | 16,735 | 4.7 |
| Non-agricultural soil | 777 | 311 | 1088 | 21,114 | 5.2 |
| Rootstock | |||||
| Richter 110 | 828 | 78 | 906 | 19,387 | 4.7 |
| 1103 Paulsen | 1024 | 213 | 1237 | 18,011 | 6.9 |
| Factors/Effect | Shoot Length | Root Biomass | Mycorrhizal Colonization |
|---|---|---|---|
| Soil type | 0.351 | 0.708 | 0.716 |
| Rootstock | 0.635 | 0.884 | 0.012 |
| Land use | 0.903 | 0.675 | 0.129 |
| Soil type × Rootstock | 0.288 | 0.351 | 0.852 |
| Soil type × Land use | 0.052 | 0.123 | 0.028 |
| Rootstock × Land use | 0.604 | 0.382 | 0.241 |
| Soil type × Rootstock × Land use | 0.279 | 0.075 | 0.620 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fors, R.O.; Sorci-Uhmann, E.; Santos, E.S.; Silva-Flores, P.; Abreu, M.M.; Viegas, W.; Nogales, A. Influence of Soil Type, Land Use, and Rootstock Genotype on Root-Associated Arbuscular Mycorrhizal Fungi Communities and Their Impact on Grapevine Growth and Nutrition. Agriculture 2023, 13, 2163. https://doi.org/10.3390/agriculture13112163
Fors RO, Sorci-Uhmann E, Santos ES, Silva-Flores P, Abreu MM, Viegas W, Nogales A. Influence of Soil Type, Land Use, and Rootstock Genotype on Root-Associated Arbuscular Mycorrhizal Fungi Communities and Their Impact on Grapevine Growth and Nutrition. Agriculture. 2023; 13(11):2163. https://doi.org/10.3390/agriculture13112163
Chicago/Turabian StyleFors, Rosalba O., Emilia Sorci-Uhmann, Erika S. Santos, Patricia Silva-Flores, Maria Manuela Abreu, Wanda Viegas, and Amaia Nogales. 2023. "Influence of Soil Type, Land Use, and Rootstock Genotype on Root-Associated Arbuscular Mycorrhizal Fungi Communities and Their Impact on Grapevine Growth and Nutrition" Agriculture 13, no. 11: 2163. https://doi.org/10.3390/agriculture13112163
APA StyleFors, R. O., Sorci-Uhmann, E., Santos, E. S., Silva-Flores, P., Abreu, M. M., Viegas, W., & Nogales, A. (2023). Influence of Soil Type, Land Use, and Rootstock Genotype on Root-Associated Arbuscular Mycorrhizal Fungi Communities and Their Impact on Grapevine Growth and Nutrition. Agriculture, 13(11), 2163. https://doi.org/10.3390/agriculture13112163