
Morphological Characterization of 1322 Winter Wheat (Triticum aestivum L.) Varieties from EU Referent Collection

Abstract
:1. Introduction
2. Materials and Methods
2.1. Investigated DUS Traits
2.2. Statistical Analysis
3. Results
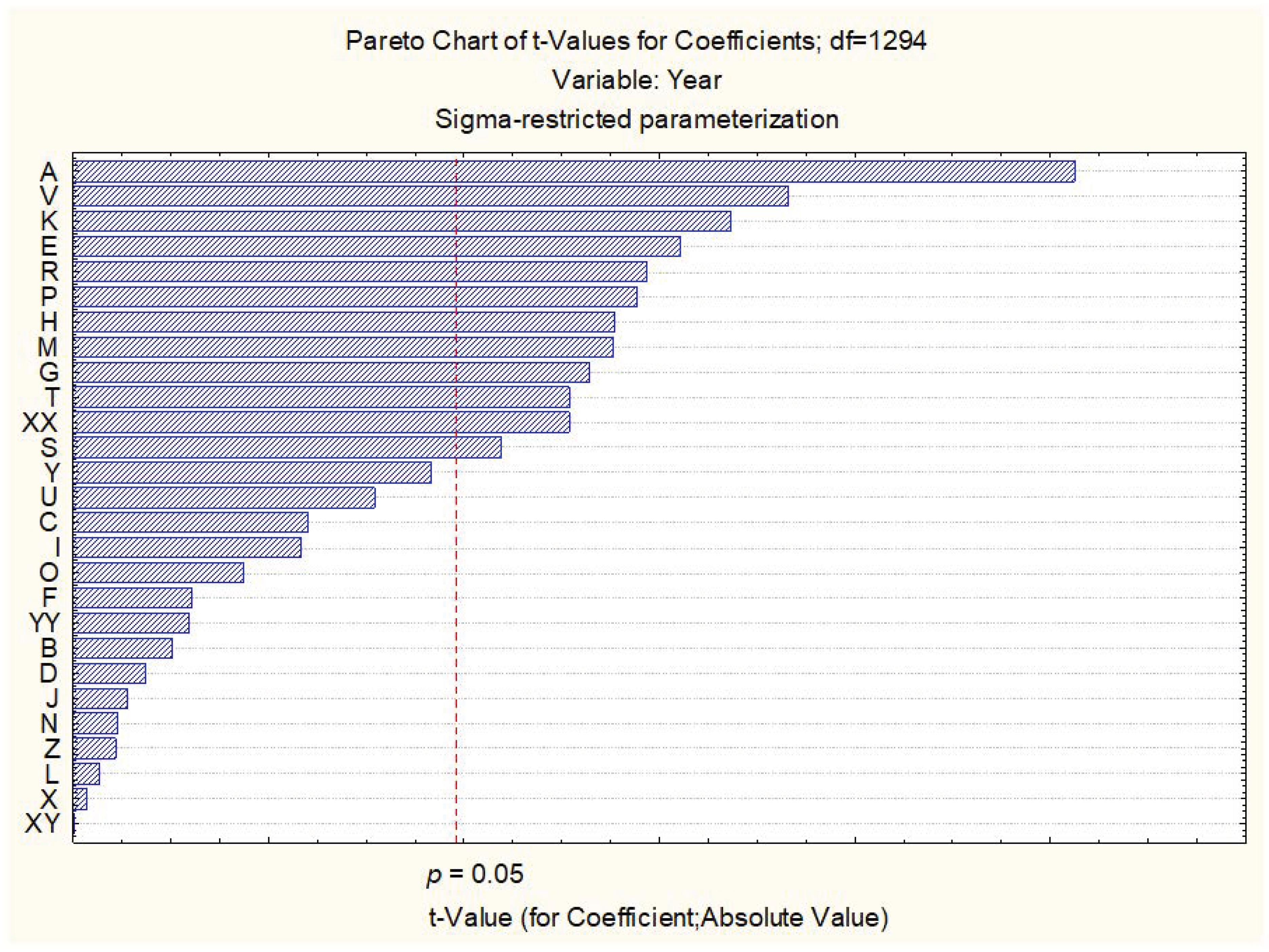
3.1. Regression Analysis of Investigated DUS Traits
3.2. Principal Component Analysis of Twelve Investigated DUS Traits
4. Discussion
4.1. Morphological Characters of Different Plant Tissues or Whole Plant
4.1.1. Seed Characteristics
4.1.2. Flag Leaf and the Whole Plants Characteristics
4.1.3. Glaucosity of Different Wheat Tissues
4.1.4. Ear Morphological Characteristics
4.2. Morphological Traits That Changed over the Period from 2006 to 2023
4.3. The Most Related Morphological Traits over the Years
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Grote, U.; Fasse, A.; Nguyen, T.T.; Erenstein, O. Food security and the dynamics of wheat and maize value chains in Africa and Asia. Front. Sustain. Food Syst. 2021, 4, 617009. [Google Scholar] [CrossRef]
- Spanic, V.; Jukic, G.; Zoric, M.; Varnica, I. Some agronomic properties of winter wheat genotypes grown at different locations in Croatia. Agriculture 2024, 14, 4. [Google Scholar] [CrossRef]
- World Food Situation. Available online: https://www.fao.org/worldfoodsituation/csdb/en/ (accessed on 13 December 2022).
- Yue, Y.; Zhang, P.; Shang, Y. The potential global distribution and dynamics of wheat under multiple climate change scenarios. Sci. Total Environ. 2019, 688, 1308–1318. [Google Scholar] [CrossRef] [PubMed]
- Sunic, K.; Brkljacic, L.; Vukovic, R.; Katanic, Z.; Salopek-Sondi, B.; Spanic, V. Fusarium Head Blight infection induced responses of six winter wheat varieties in ascorbate–glutathione pathway, photosynthetic efficiency and stress hormones. Plants 2023, 12, 3720. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tao, F.; Hao, Y.; Tong, J.; Xiao, Y.; He, Z.; Reynolds, M. Variations in phenological, physiological, plant architectural and yield-related traits, their associations with grain yield and genetic basis. Ann. Bot. 2023, 131, 503–519. [Google Scholar] [CrossRef]
- Wu, X.; Chang, X.; Jing, R. Genetic insight into yield-associated traits of wheat grown in multiple rain-fed environments. PLoS ONE 2012, 7, e31249. [Google Scholar] [CrossRef] [PubMed]
- Hyles, J.; Bloomfield, M.T.; Hunt, J.R.; Trethowan, R.M.; Trevaskis, B. Phenology and related traits for wheat adaptation. Heredity 2020, 125, 417–430. [Google Scholar] [CrossRef]
- Govindaraj, M.; Vetriventhan, M.; Srinivasan, M. Importance of genetic diversity assessment in crop plants and its recent advances: An overview of its analytical perspectives. Genet. Res. Int. 2015, 2015, 431487. [Google Scholar] [CrossRef]
- Min, B.; Salt, L.; Wilde, P.; Kosik, O.; Hassall, K.; Przewieslik-Allen, A.; Burridge, A.J.; Poole, M.; Snape, J.; Wingen, L.; et al. Genetic variation in wheat grain quality is associated with differences in the galactolipid content of flour and the gas bubble properties of dough liquor. Food Chem. 2020, 6, 100093. [Google Scholar] [CrossRef] [PubMed]
- Franco, J.; Crossa, J.; Ribaut, J.M.; Betran, J.; Warburton, M.; Khairallah, M. A method for combining molecular markers and phenotypic attributes for classifying plant genotypes. Theor. Appl. Genet. 2001, 103, 944–952. [Google Scholar] [CrossRef]
- Yu, J.K.; Chung, Y.S. Plant variety protection: Current practices and insights. Genes 2021, 12, 1127. [Google Scholar] [CrossRef] [PubMed]
- UPOV. International Convention for the Protection of New Varieties of Plants of December 2, 1961, as Revised at Geneva on November 10, 1972, on October 23, 1978, and on March 19, 1991. Available online: https://www.upov.int/edocs/pubdocs/en/upov_pub_221.pdf (accessed on 15 January 2024).
- CPVO/TP-003/5. Available online: https://cpvo.europa.eu/sites/default/files/documents/triticum_aestivum_5.pdf (accessed on 4 January 2024).
- Khadka, K.; Earl, H.J.; Raizada, M.N.; Navabi, A. A physio-morphological trait-based approach for breeding drought tolerant wheat. Front. Plant Sci. 2020, 11, 715. [Google Scholar] [CrossRef] [PubMed]
- The World Bank. Available online: https://data.worldbank.org/indicator/AG.YLD.CREL.KG (accessed on 27 March 2024).
- Swarup, S.; Cargill, E.J.; Crosby, K.; Flagel, L.; Kniskern, J.; Glenn, K.C. Genetic diversity is indispensable for plant breeding to improve crops. Crop Sci. 2020, 61, 839–852. [Google Scholar] [CrossRef]
- Malik, R.; Sharma, H.; Sharma, I.; Kundu, S.; Verma, A.; Sheoran, S.; Kumar, R.; Chatrath, R. Genetic diversity of agro-morphological characters in Indian wheat varieties using GT biplot. Aust. J. Crop Sci. 2014, 8, 1266–1271. [Google Scholar]
- Wang, L.; Zheng, Y.; Duan, L.; Wang, M.; Wang, H.; Li, H.; Li, R.; Zhang, H. Artificial selection trend of wheat varieties released in Huang-Huai-Hai region in China evaluated using DUS testing characteristics. Front. Plant Sci. 2022, 13, 898102. [Google Scholar] [CrossRef] [PubMed]
- Dreisigacker, S.; Zhang, P.; Warburton, M.L.; Skovmand, B.; Hoisington, D.; Melchinger, A.E. Genetic diversity among and within CIMMYT wheat landrace accessions investigated with SSRs and implications for plant genetic resources management. Crop Sci. 2005, 45, 653–661. [Google Scholar] [CrossRef]
- Kohyama, N.; Chono, M.; Nakagawa, H.; Matsuo, Y.; Ono, H.; Matsunaka, H. Flavonoid compounds related to seed coat color of wheat. Biosci. Biotechnol. Biochem. 2017, 81, 2112–2118. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, H.; Liu, J.; Huang, T.; Zhang, X.; Xie, H.; Guo, Y.; Wang, Q.; Zhang, P.; Qin, P. Grain color formation and analysis of correlated genes by metabolome and transcriptome in different wheat lines at maturity. Front. Nutr. 2023, 10, 1112497. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.J.; Shelton, D.R.; Martin, T.J.; Sears, R.G.; Williams, E.; Graybosch, R.A. Grain color stability and classification of hard white wheat in the U.S. Euphytica 2001, 119, 101–106. [Google Scholar] [CrossRef]
- Niranjana, M.; Jha, S.K.; Mallick, N.; Verma, A.; Singh, B.; Ahlawat, A.; Tomar, S.M.S.; Vinod. Distribution of genes producing phenol colour reaction in grains of wheat and its related species, mode of inheritance and breeding for low polyphenol activity. Indian J. Genet. 2018, 78, 433–442. [Google Scholar]
- Bogdanova, E.D.; Sarbaev, A.T.; Makhmudova, K.K. Resistance of common wheat to bunt. In Proceedings of the Research Conference on Genetics, Moscow, Russia, 26–27 February 2002. [Google Scholar]
- Sakamoto, T.; Morinaka, Y.; Ohnishi, T.; Sunohara, H.; Fujioka, S.; Ueguchi-Tanaka, M.; Mizutani, M.; Sakata, K.; Takatsuto, S.; Yoshida, S.; et al. Erect leaves caused by brassinosteroid deficiency increase biomass production and grain yield in rice. Nat. Biotechnol. 2006, 24, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Shaaf, S.; Bretani, G.; Biswas, A.; Fontana, I.M.; Rossini, L. Genetics of barley tiller and leaf development. J. Integr. Plant Biol. 2018, 61, 226–256. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.A.; Ali, Z.; Tahir, M.H.N.; Ghaffar, A.; Ahmad, J. Genetic effects of GA-responsive dwarfing gene Rht13 on plant height, peduncle length, internodal length and grain yield of wheat under drought stress. Genes 2023, 14, 699. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Dutt, D.; Tyagi, C.H. Complete characterization of wheat straw (Triticum aestivum PBW-343 L. Emend. Fiori & Paul.)—A renewable source of fibers for pulp and paper making. BioResources 2011, 6, 154–177. [Google Scholar]
- Liu, X.; Feakins, S.J.; Dong, X.; Xue, Q.; Marek, T.; Leskovar, D.I.; Neely, C.B.; Ibrahim, A. Experimental study of leaf wax n-alkane response in winter wheat cultivars to drought conditions. Org. Geochem. 2017, 113, 210–223. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, W.; Li, W. Genetic interactions underlying the biosynthesis and inhibition of β-diketones in wheat and their impact on glaucousness and cuticle permeability. PLoS ONE 2013, 8, e54129. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Sun, M.; Xie, Y.; Wang, F.; Zhao, Z. Photochemical and antioxidative responses of the glume and flag leaf to seasonal senescence in wheat. Front. Plant Sci. 2015, 6, 358. [Google Scholar] [CrossRef] [PubMed]
- Dixon, L.E.; Boden, S.A. A modified intron of VRT2 drives glume and grain elongation in wheat. Mol. Plant 2021, 14, 1421–1423. [Google Scholar] [CrossRef]
- Hu, X.; Zuo, J. Population genomics and haplotype analysis in bread wheat identify a gene regulating glume pubescence. Front. Plant Sci. 2022, 13, 897772. [Google Scholar] [CrossRef]
- Artemenko, N.V.; Genaev, M.A.; Epifanov, R.U.I.; Komyshev, E.G.; Kruchinina, Y.V.; Koval, V.S.; Goncharov, N.P.; Afonnikov, D.A. Image-based classification of wheat spikes by glume pubescence using convolutional neural networks. Front. Plant Sci. 2024, 14, 1336192. [Google Scholar] [CrossRef]
- Dandrifosse, S.; Ennadifi, E.; Carlier, A.; Gosselin, B.; Dumont, B.; Mercatoris, B. Deep learning for wheat ear segmentation and ear density measurement: From heading to maturity. Comput. Electron. Agric. 2022, 199, 107161. [Google Scholar] [CrossRef]
- Maydup, M.L.; Antonietta, M.; Graciano, C.; Guiamet, J.J.; Tambussi, E.J. The contribution of the awns of bread wheat (Triticum aestivum L.) to grain filling: Responses to water deficit and the effects of awns on ear temperature and hydraulic conductance. Field Crops Res. 2014, 167, 102–111. [Google Scholar] [CrossRef]
- Fujii, K.; Tsuji, T.; Yoshida, T.; Maruyama-Funatsuki, W.; Tatsuya, M.; Ikeda, T. Applicability of ‘glume color’ controlled by Rg-B1 locus as a field-marker to detect Glu-B3 alleles encoding low-molecular-weight glutenin subunits in common wheat. Breed. Sci. 2011, 61, 11–16. [Google Scholar] [CrossRef]
- Othmani, A.; Mosbahi, M.; Ayed, S.; Slim-Amara, H.; Boubaker, M. Morphological characterization of some Tunisian bread wheat (Triticum aestivum L.) accessions. J. New Sci. Agric. Biotechnol. 2015, 15, 503–510. [Google Scholar]
- Ateş Sönmezoğlu, Ö.; Betül, B.; Yildirim, A.; Kandemir, N.; Aydin, N. Genetic characterization of Turkish bread wheat landraces based on microsatellite markers and morphological characters. Turk. J. Biol. 2012, 36, 589–597. [Google Scholar] [CrossRef]
- Mondal, S.; Dutta, S.; Crespo-Herrera, L.; Huerta-Espino, J.; Braun, H.J.; Singh, R.P. Fifty years of semi-dwarf spring wheat breeding at CIMMYT: Grain yield progress in optimum, drought and heat stress environments. Field Crops Res. 2020, 250, 107757. [Google Scholar] [CrossRef]
- Tsenov, N.; Gubatov, T.; Yanchev, I. Date of ear emergence: A factor for notable changing the grain yield of modern winter wheat varieties in different environments of Bulgaria. Agric. Sci. Technol. 2021, 13, 12–18. [Google Scholar] [CrossRef]
- Duvnjak, J.; Katanic, Z.; Sarcevic, H.; Spanic, V. Analysis of the photosynthetic parameters, grain yield, and quality of different winter wheat varieties over a two-year period. Agronomy 2024, 14, 478. [Google Scholar] [CrossRef]
- Fradgley, N.; Gardner, K.A.; Bentley, A.R.; Howell, P.; Mackay, I.J.; Scott, M.F.; Mott, R.; Cockram, J. Multi-trait ensemble genomic prediction and simulations of recurrent selection highlight importance of complex trait genetic architecture for long-term genetic gains in wheat. In Silico Plants 2023, 5, diad002. [Google Scholar] [CrossRef]
- Wen, H.; Wang, Y.; Wu, B.; Feng, Y.; Dang, Y.; Yang, B.; Ma, X.; Qiao, L. Analysis of wheat wax regulation mechanism by liposome and transcriptome. Front. Genet. 2021, 12, 757920. [Google Scholar] [CrossRef]
- Li, X.; Tang, Y.; Zhou, C.; Lv, J. Contributions of glume and awn to photosynthesis, 14C assimilates and grain weight in wheat ears under drought stress. Heliyon 2023, 9, e21136. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Label | DUS Characteristic | States of Expression |
|---|---|---|
| A | Seed: color | 1 white, 2 reddish, 3 purple, 4 bluish |
| B | Seed: coloration with phenol | 1 absent or very light, 3 light, 5 medium, 7 dark, 9 very dark |
| C | Coleoptile: anthocyanin coloration | 1 absent or very weak, 3 weak, 5 medium, 7 strong, 9 very strong |
| D | Plant: growth habit | 1 erect, 3 semi-erect, 5 intermediate, 7 semi-prostrate, 9 prostrate |
| E | Plants: frequency of plants with recurved flag leaves | 1 absent or very low, 3 low, 5, medium, 7 high, 9 very high |
| F | Flag leaf: anthocyanin coloration of auricles | 1 absent or very weak, 2 medium, 3 strong |
| G | Time of ear emergence (first spikelet visible on 50% of ears) | 1 very early, 3 early, 5 medium, 7 late, 9 very late |
| H | Flag leaf: glaucosity of sheath | 1 absent or very weak, 3 weak, 5 medium, 7 strong, 9 very strong |
| I | Fleg leaf: glaucosity of blade | 1 absent or very weak, 3 weak, 5 medium, 7 strong, 9 very strong |
| J | Ear: glaucosity | 1 absent or very weak, 3 weak, 5 medium, 7 strong, 9 very strong |
| K | Culm: glaucosity of neck | 1 absent or very weak, 3 weak, 5 medium, 7 strong, 9 very strong |
| L | Lower glume: hairiness of external surface | 1 absent, 9 present |
| M | Plant: length (stem, ears, awns, and scurs) | 1 very short, 3 short, 5 medium, 7 long, 9 very long |
| N | Straw: pith in cross section | 1 thin, 2 medium, 3 thick or filled |
| O | Ear: density | 1 very lax, 3 lax, 5 medium, 7 dense, 9 very dense |
| P | Ear: length (excluding awns and scurs) | 1 very short, 3 short, 5 medium, 7 long, 9 very long |
| R | Ear: scurs and awns | 1 both absent, 2 scurs present, 3 awns present |
| S | Ear: length of scurs or awns | 1 very short, 3 short, 5 medium, 7 long, 9 very long |
| T | Ear: color | 1 white, 2 colored (red) |
| U | Ear: shape in profile | 1 tapering, 2 parallel sided, 3 slightly clavate, 4 strongly clavate, 5 fusiform |
| V | Apical rachis segment: area of hairiness of the convex surface | 1 absent or very small, 3 small, 5 medium, 7 large, 9 very large |
| Z | Lower glume: shoulder width | 1 very narrow, 3 narrow, 5 medium, 7 broad, 9 very broad |
| X | Lower glume: shoulder shape | 1 strongly sloping, 3 slightly sloping, 5 horizontal, 7 slightly elevated, 9 strongly elevated |
| Y | Lower glume: length of beak | 1 very short, 3 short, 5 medium,7 long, 9 very long |
| XX | Lower glume: shape of beak | 1 straight, 3 slightly curved, 5 moderately curved, 7 strongly curved, 9 geniculate |
| YY | Lower glume: area of hairiness on internal surface | 1 very small, 3 medium, 5 very large |
| XY | Seasonal type | 1 winter type, 2 alternative type, 3 spring type |
| R = 0.37474083 R2 = 0.14043069 Adjusted R2 = 0.12249532 F(27,1294) = 7.8298 p | ||||
|---|---|---|---|---|
| Unstandardized Beta (B) | Standard Error of B | t(1294) | p-Level | |
| Seed: color | 0.279 | 0.055 | 5.126 | 0.000 ** |
| Seed: coloration with phenol | 0.009 | 0.018 | 0.504 | 0.614 |
| Coleoptile: anthocyanin coloration | −0.056 | 0.046 | −1.202 | 0.229 |
| Plant: growth habit | 0.012 | 0.031 | 0.368 | 0.713 |
| Plants: frequency of plants with recurved flag leaves | −0.064 | 0.021 | −3.107 | 0.002 ** |
| Flag leaf: anthocyanin coloration of auricles | 0.069 | 0.114 | 0.604 | 0.546 |
| Time of ear emergence (first spikelet visible on 50% of ears) | −0.059 | 0.022 | −2.644 | 0.008 ** |
| Flag leaf: glaucosity of sheath | −0.099 | 0.036 | −2.767 | 0.006 ** |
| Fleg leaf: glaucosity of blade | 0.033 | 0.028 | 1.166 | 0.244 |
| Ear: glaucosity | 0.008 | 0.028 | 0.277 | 0.782 |
| Culm: glaucosity of neck | 0.117 | 0.035 | 3.368 | 0.001 ** |
| Lower glume: hairiness of external surface | −0.007 | 0.050 | −0.135 | 0.893 |
| Plant: length (stem, ears, awns, and scurs) | 0.066 | 0.024 | 2.767 | 0.006 ** |
| Straw: pith in cross section | 0.013 | 0.058 | 0.229 | 0.819 |
| Ear: density | 0.028 | 0.032 | 0.873 | 0.383 |
| Ear: length (excluding awns and scurs) | 0.071 | 0.025 | 2.886 | 0.004 ** |
| Ear: scurs and awns | −0.387 | 0.132 | −2.937 | 0.003 ** |
| Ear: length of scurs or awns | 0.072 | 0.033 | 2.189 | 0.029 * |
| Ear: color | 0.232 | 0.091 | 2.540 | 0.011 * |
| Ear: shape in profile | 0.030 | 0.019 | 1.542 | 0.123 |
| Apical rachis segment: area of hairiness of convex surface | −0.054 | 0.015 | −3.662 | 0.000 ** |
| Lower glume: shoulder width | 0.004 | 0.021 | 0.218 | 0.827 |
| Lower glume: shoulder shape | −0.001 | 0.020 | −0.073 | 0.942 |
| Lower glume: length of beak | 0.050 | 0.027 | 1.833 | 0.067 |
| Lower glume: shape of beak | 0.053 | 0.021 | 2.539 | 0.011 * |
| Lower glume: area of hairiness on internal surface | 0.014 | 0.024 | 0.596 | 0.551 |
| Seasonal type | 0.001 | 0.290 | 0.003 | 0.998 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spanic, V.; Lalic, Z.; Berakovic, I.; Jukic, G.; Varnica, I. Morphological Characterization of 1322 Winter Wheat (Triticum aestivum L.) Varieties from EU Referent Collection. Agriculture 2024, 14, 551. https://doi.org/10.3390/agriculture14040551
Spanic V, Lalic Z, Berakovic I, Jukic G, Varnica I. Morphological Characterization of 1322 Winter Wheat (Triticum aestivum L.) Varieties from EU Referent Collection. Agriculture. 2024; 14(4):551. https://doi.org/10.3390/agriculture14040551
Chicago/Turabian StyleSpanic, Valentina, Zvonimir Lalic, Ivica Berakovic, Goran Jukic, and Ivan Varnica. 2024. "Morphological Characterization of 1322 Winter Wheat (Triticum aestivum L.) Varieties from EU Referent Collection" Agriculture 14, no. 4: 551. https://doi.org/10.3390/agriculture14040551