Effects of Shade on Nitrogen and Phosphorus Acquisition in Cereal-Legume Intercropping Systems

Abstract
:1. Introduction
2. Results

{kind=link}
{kind=link}
{kind=link}
| Biomass (g plant−1) | ||||
|---|---|---|---|---|
| Shoot | Root | Total biomass | Shoot: root | |
| Design | Shade | |||
| Monocropping | 0.35 ± 0.063 az | 0.89 ± 0.089 az | 1.25 ± 0.152 az | 0.39 ± 0.034 az |
| Intercropping | ||||
| w/ soybean | 0.32 ± 0.019 az | 0.81 ± 0.055 az | 1.13 ± 0.073 az | 0.40 ± 0.007 az |
| w/ alfalfa | 0.37 ± 0.049 az | 1.05 ± 0.043 az | 1.42 ± 0.087 az | 0.35 ± 0.037 az |
| No Shade | ||||
| Monocropping | 0.65 ± 0.046 ay | 1.32 ± 0.145 ay | 1.97 ± 0.190 ay | 0.50 ± 0.025 az |
| Intercropping | ||||
| w/ soybean | 0.36 ± 0.006 bz | 1.00 ± 0.083 az | 1.37 ± 0.089 az | 0.36 ± 0.023 az |
| w/ alfalfa | 0.60 ± 0.073 abz | 1.37 ± 0.126 az | 1.97 ± 0.143 ay | 0.44 ± 0.063 az |
| Rhizosphere Parameter | |||
|---|---|---|---|
| Root surface area (cm2) | Root length (cm) | ||
| Design | Shade | ||
| Wheat | |||
| Monocropping | 63.8 ± 6.7 az | 799.7 ± 95.5 az | |
| Intercropped w/soybean | 54.9 ± 6.0 az | 667.4 ± 89.0 az | |
| Intercropped w/alfalfa | 65.5 ± 5.8 az | 796.2 ± 71.1 az | |
| Associated legume | |||
| Soybean | 24.0 ± 3.3 z | 350.8 ± 60.3 z | |
| Alfalfa | 17.2 ± 2.4 z | 252.7 ± 36.2z | |
| No Shade | |||
| Wheat | |||
| Monocropping | 94.8 ± 17.2 az | 1101.7 ± 165.7 ay | |
| Intercropped w/soybean | 54.9 ± 3.7 bz | 672.8 ± 71.1 bz | |
| Intercropped w/alfalfa | 75.6 ± 0.7 bz | 810.2 ± 12.1 bz | |
| Associated legume | |||
| Soybean | 24.3 ± 4.1 z | 366.0 ± 47.8 z | |
| Alfalfa | 12.8 ± 1.7 z | 194.9 ± 26.2 z | |
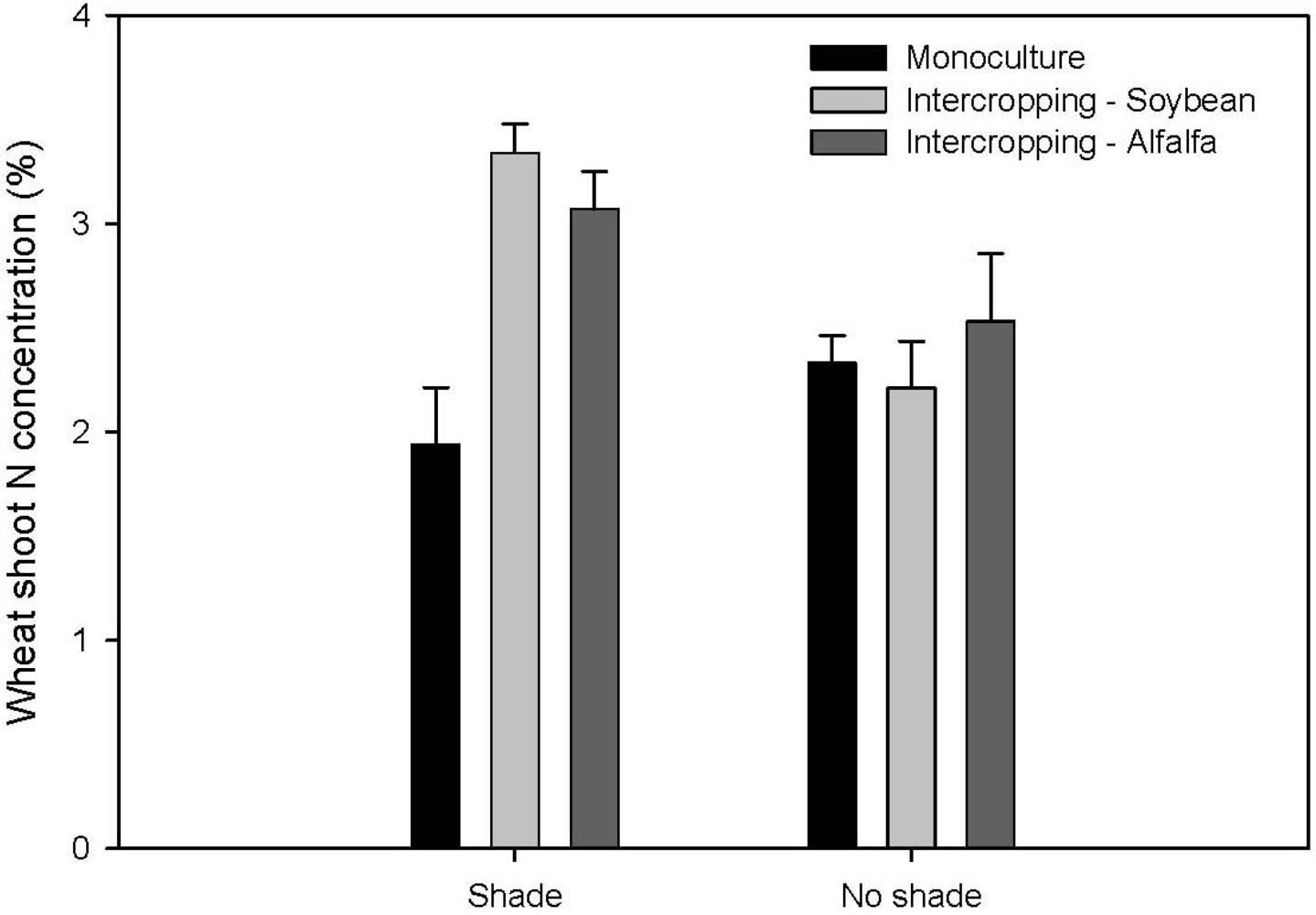
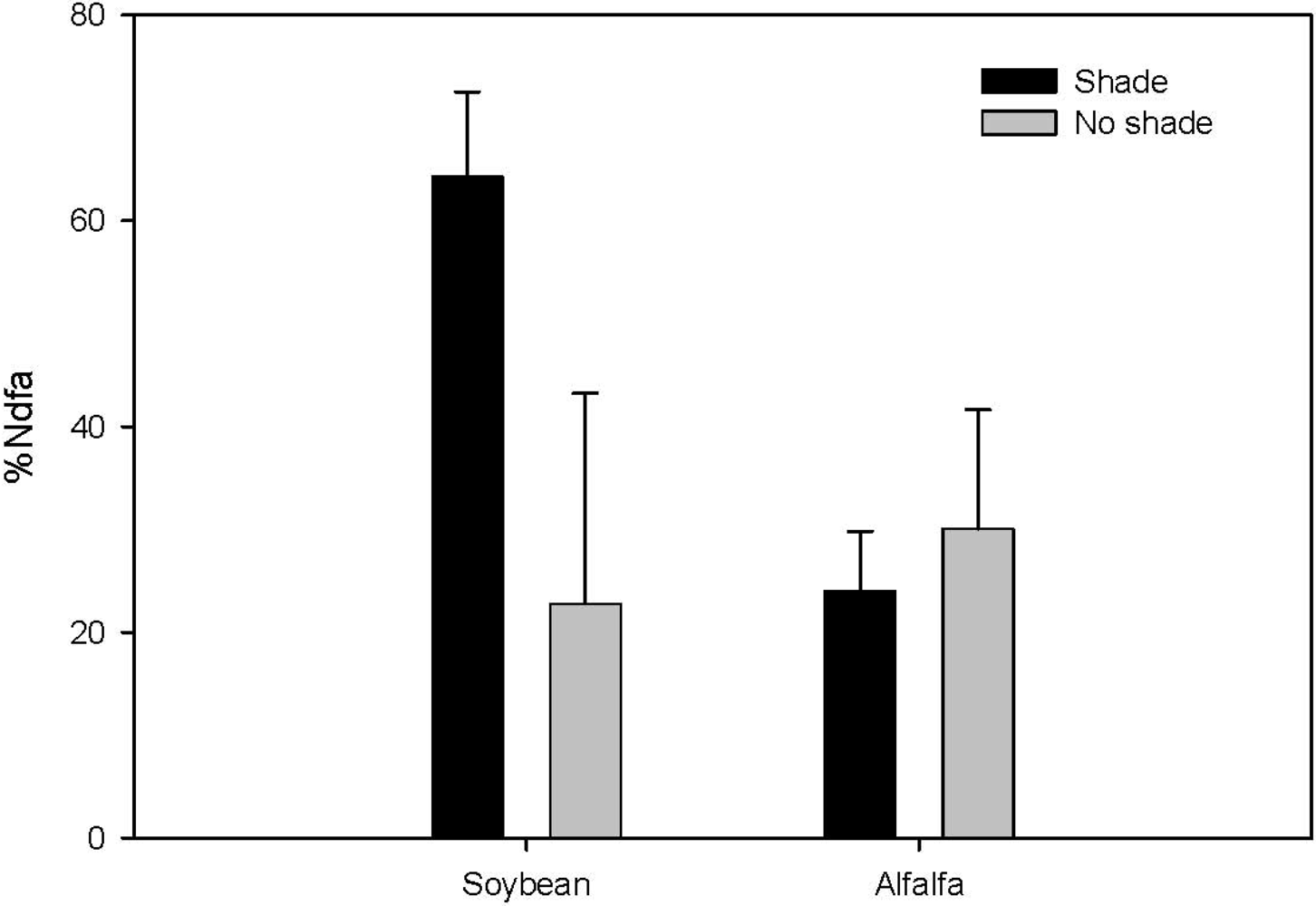
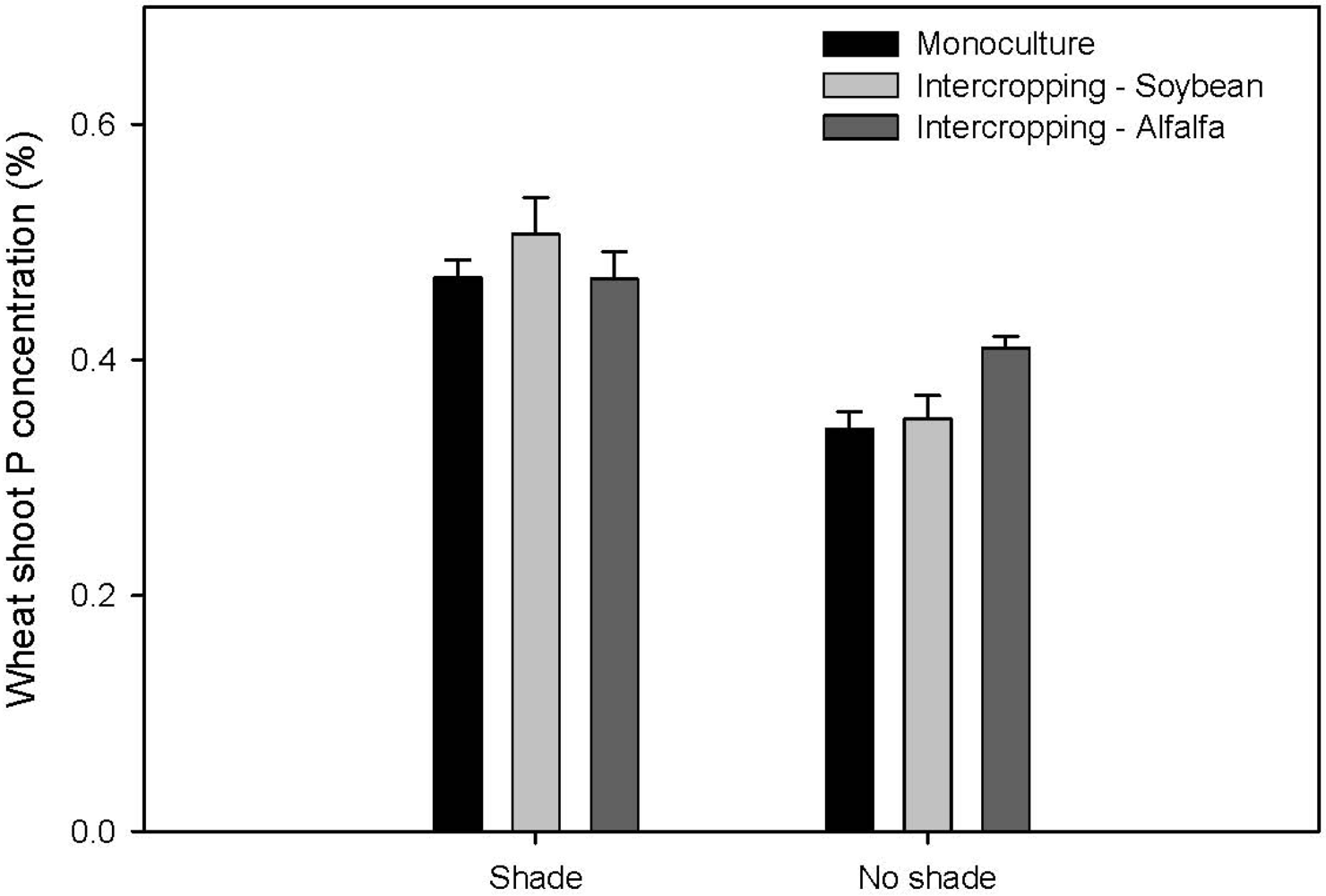
2.1. Nutrition



2.2. Soil Status
| Parameter | |||
|---|---|---|---|
| Total N (%) | Extractable P (mg Kg−1 dry soil) | pH | |
| Design | Shade | ||
| Monocropping | 0.10 ± 0.003 a | 24.2 ± 2.19 a | 7.1 ± 0.09 a |
| Intercropping | |||
| w/ soybean | 0.09 ± 0.023 a | 22.8 ± 1.00 a | 7.2 ± 0.06 a |
| w/ alfalfa | 0.10 ± 0.006 a | 19.4 ± 2.35 a | 6.8 ± 0.13 b |
| No Shade | |||
| Monocropping | 0.08 ± 0.011 a | 20.0 ± 1.02 a | 7.1 ± 0.08 a |
| Intercropping | |||
| w/ soybean | 0.10 ± 0.008 a | 20.8 ± 1.15 a | 6.9 ± 0.08 a |
| w/ alfalfa | 0.09 ± 0.014 a | 18.7 ± 1.01 a | 6.9 ± 0.12 a |
3. Discussion
3.1. Intercropping Effects on Wheat Nutrient Acquisition and Growth
3.2. Light Availability and Plant Performance
4. Experimental Section
4.1. Plant Growth Conditions
4.2. Steady-State Supply of Nitrogen
4.3. Experimental Design
4.4. Plant and Soil Analysis
4.5. N2-Fixation Measurements
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
References
- Schroth, G.; Lehmann, J.; Rodrigues, M.; Barros, E.; Macêdo, J. Plant-soil interactions in multistrata agroforestry in the humid tropics. Agrofor. Syst. 2001, 53, 85–102. [Google Scholar]
- Swift, M.; Izac, A.; van Noordwijk, M. Biodiversity and ecosystem services in agricultural landscapes-are we asking the right questions? Agric. Ecosyst. Environ. 2004, 104, 113–134. [Google Scholar] [CrossRef]
- Crews, T.E.; Peoples, M.B. Can the synchrony of nitrogen supply and crop demand be improved in legume and fertilizer-based agroecosystems? Nutr. Cycl. Agorecosyst. 2005, 72, 101–120. [Google Scholar] [CrossRef]
- Vandermeer, J. The Ecology of Intercropping; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Tilman, D.; Cassman, K.; Matson, P.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar]
- Isaac, M.; Kimaro, A. Diagnosis of nutrient imbalances with vector analysis in agroforestry systems. J. Environ. Qual. 2011, 40, 1–7. [Google Scholar]
- Jose, S.; Gillespie, A.; Pallardy, S. Interspecific interactions in temperate agroforestry. Agrofor. Syst. 2004, 61, 237–255. [Google Scholar]
- Thevathasan, N.; Gordon, A. Ecology of tree intercropping systems in the North temperate region: Experiences from southern Ontario, Canada. Agrofor. Syst. 2004, 61, 257–268. [Google Scholar]
- Vandermeer, J. The ecological basis of alternative agriculture. Annu. Rev. Ecol. Syst. 1995, 26, 201–224. [Google Scholar]
- Høgh-Jensen, H.; Schjoerring, J.; Soussana, J. The influence of phosphorus deficiency on growth and nitrogen fixation of white clover plants. Ann. Bot. 2002, 90, 745–753. [Google Scholar]
- Li, L.; Zhang, F.; Li, X.; Christie, P.; Sun, J.; Yang, S.; Tang, C. Interspecific facilitation of nutrient uptake by intercropped maize and faba bean. Nutr. Cycl. Agroecosyst. 2003, 65, 61–71. [Google Scholar]
- Sierra, J.; Nygren, P. Transfer of N fixed by a legume tree to the associated grass in a tropical silvopastoral system. Soil Biol. Biochem. 2006, 38, 1893–1903. [Google Scholar]
- Chu, G.; Shen, Q.; Cao, J. Nitrogen fixation and N transfer from peanut to rice cultivated in aerobic soil in an intercropping system and its effect on soil N fertility. Plant Soil 2004, 263, 17–27. [Google Scholar]
- Martin, R.; Voldeng, D.; Smith, D. Nitrogen transfer from nodulating soybean to maize or to nonnodulating soybean in intercrops, the 15N dilution method. Plant Soil 1991, 132, 53–63. [Google Scholar]
- Jensen, E. Grain yield, symbiotic N2-fixation and interspecific competition for inorganic N in pea-barley intercrops. Plant Soil 1996, 182, 25–38. [Google Scholar]
- Li, H.; Shen, J.; Zhang, F.; Clairotte, M.; Drevon, J.; Le Cadre, E.; Hinsinger, P. Dynamics of phosphorus fractions in the rhizosphere of common bean (Phaseolus vulgaris L.) and durum wheat (Triticum turgidum durum L.) grown in monocropping and intercropping systems. Plant Soil 2008, 312, 139–150. [Google Scholar] [CrossRef]
- Carlsson, G.; Palmborg, C.; Jumpponen, A.; Scherer-Lorenzen, M.; Högberg, P.; Huss-Danell, K. N2 fixation in three perennial Trifolium species in experimental grasslands of varied plant species richness and composition. Plant Ecol. 2009, 205, 87–104. [Google Scholar] [CrossRef]
- Li, L.; Tang, C.; Rengel, Z.; Zhang, F. Chickpea facilitates phosphorus uptake by intercropped wheat from an organic phosphorus source. Plant Soil 2003, 248, 297–303. [Google Scholar]
- Wang, X.; Tang, C.; Guppy, C.; Sale, P. Phosphorus acquisition characteristics of cotton (Gossypium hirsutum L.), wheat (Triticum aestivum L.) and white lupin (Lupinus albus L.) under P deficient conditions. Plant Soil 2008, 312, 117–128. [Google Scholar] [CrossRef]
- Peoples, M.; Gault, R.; Lean, B.; Sykes, J.; Brockwell, J. Nitrogen fixation by soybean in commercial irrigated crops of central and southern New South Wales. Soil Biol. Biochem. 1995, 27, 553–561. [Google Scholar]
- Carlsson, G.; Huss-Danell, K. Nitrogen fixation in perennial forage legumes in the field. Plant Soil 2003, 253, 353–272. [Google Scholar]
- Houlton, B.Z.; Wang, Y.-P.; Vitousek, P.M.; Field, C.B. A unifying framework for dinitrogen fixation in the terrestrial biosphere. Nature 2008, 454, 327–331. [Google Scholar]
- Isaac, M.E.; Harmand, J.M.; Drevon, J.J. Growth and nitrogen acquisition strategies of Acacia senegal seedlings under exponential phosphorus additions. J. Plant Physiol. 2011, 168, 776–781. [Google Scholar]
- Isaac, M.E.; Harmand, J.M.; Lesueur, D.; Lelon, J. Tree age and soil phosphorus conditions influence N2-fixation rates and soil N dynamics in natural populations of Acacia senegal. For. Ecol. Manag. 2011, 261, 582–588. [Google Scholar]
- Schipanski, M.E.; Drinkwater, L.E.; Russelle, M.P. Understanding the variability in soybean nitrogen fixation across agroecosystems. Plant Soil 2010, 329, 379–397. [Google Scholar]
- Li, L.; Li, S.; Sun, J.; Zhou, L.; Bao, X.; Zhang, H.; Zhang, F. Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus-deficient soils. Proc. Natl. Acad. Sci. USA 2007, 104, 5903–5907. [Google Scholar]
- Chu, A.; Robertson, A. The effects of shading and defoliation on nodulation and nitrogen fixation by white clover. Plant Soil 1974, 41, 509–519. [Google Scholar]
- Timmer, V. Exponential nutrient loading: A new fertilization technique to improve seedling performance on competitive sites. New For. 1996, 13, 279–295. [Google Scholar]
- Hinsinger, P.; Gilkes, R. Dissolution of phosphate rock in the rhizosphere of five plant species grown in an acid, P-fixing mineral substrate. Geoderma 1997, 75, 231–249. [Google Scholar]
- Casarin, V.; Plassard, C.; Hinsinger, P.; Arvieu, J. Quantification of ectomycorrhizal fungal effects on the bioavailability and mobilization of soil P in the rhizosphere of Pinus pinaster. New Phytol. 2004, 163, 177–185. [Google Scholar] [CrossRef]
- Mariotti, A.; Germon, J.; Hubert, P.; Kaiser, P.; Letolle, R.; Tardieux, A.; Tardieux, P. Experimental determination of nitrogen kinetic isotope fractionation: Some principles; illustration for the denitrification and nitrification processes. Plant Soil 1981, 62, 413–430. [Google Scholar] [CrossRef]
- Mariotti, A. Natural 15N abundance measurements and atmospheric nitrogen standard calibration. Nature 1984, 311, 251–252. [Google Scholar]
- Ledgard, S.F. Nutrition, moisture and rhizobial strain influence isotopic fractionation during N2-fixation in pasture legumes. Soil Biol. Biochem. 1989, 21, 65–68. [Google Scholar] [CrossRef]
- Wanek, W.; Arndt, S.K. Difference in δ15N signatures between nodulated roots and shoots of soybean is indicative of the contribution of symbiotic N2-fixation to plant N. J. Exp. Bot. 2002, 53, 1109–1118. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Whitehead, M.; Isaac, M.E. Effects of Shade on Nitrogen and Phosphorus Acquisition in Cereal-Legume Intercropping Systems. Agriculture 2012, 2, 12-24. https://doi.org/10.3390/agriculture2010012
Whitehead M, Isaac ME. Effects of Shade on Nitrogen and Phosphorus Acquisition in Cereal-Legume Intercropping Systems. Agriculture. 2012; 2(1):12-24. https://doi.org/10.3390/agriculture2010012
Chicago/Turabian StyleWhitehead, Meighen, and Marney E. Isaac. 2012. "Effects of Shade on Nitrogen and Phosphorus Acquisition in Cereal-Legume Intercropping Systems" Agriculture 2, no. 1: 12-24. https://doi.org/10.3390/agriculture2010012