
Cucumber Rhizosphere Microbial Community Response to Biocontrol Agent Bacillus subtilis B068150



Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain
2.2. Preparation of Antibiotic Resistant Mutant from Wild-Type Isolate
2.3. B. subtilis B068150 Inoculum Preparation
2.4. Soils Source
2.5. Soils Preparation
2.6. Strain B068150 Introduction into Soils
2.7. Pot Experimental Design

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Type | Sand
(%) | Silt
(%) | Clay
(%) | Bulk Density
(g/cm) | WHC
(%) | pH | T-N
(g/kg) | OC
(g/kg) | WSOC
(mg/kg) | MBC
(mg/kg) | AOC
(mg/kg) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Dwllo loamy sand | 99.1 | 0.2 | 0.7 | 1.67 | 17 | 7.1 | 0.07 | 0.58 | 10 | 11 | 0.2 |
| Arlington sandy loam | 70.9 | 20.8 | 8.3 | 1.54 | 21 | 7.2 | 0.061 | 5.4 | 44 | 56 | 0.9 |
| Willow silty clay | 3.7 | 49.1 | 47.2 | 1.51 | 63 | 7.2 | 1.61 | 20.4 | 242 | 278 | 4.94 |
2.8. DNA Extraction, PCR Amplification, and DGGE Analysis
2.9. Statistical Analysis
3. Results
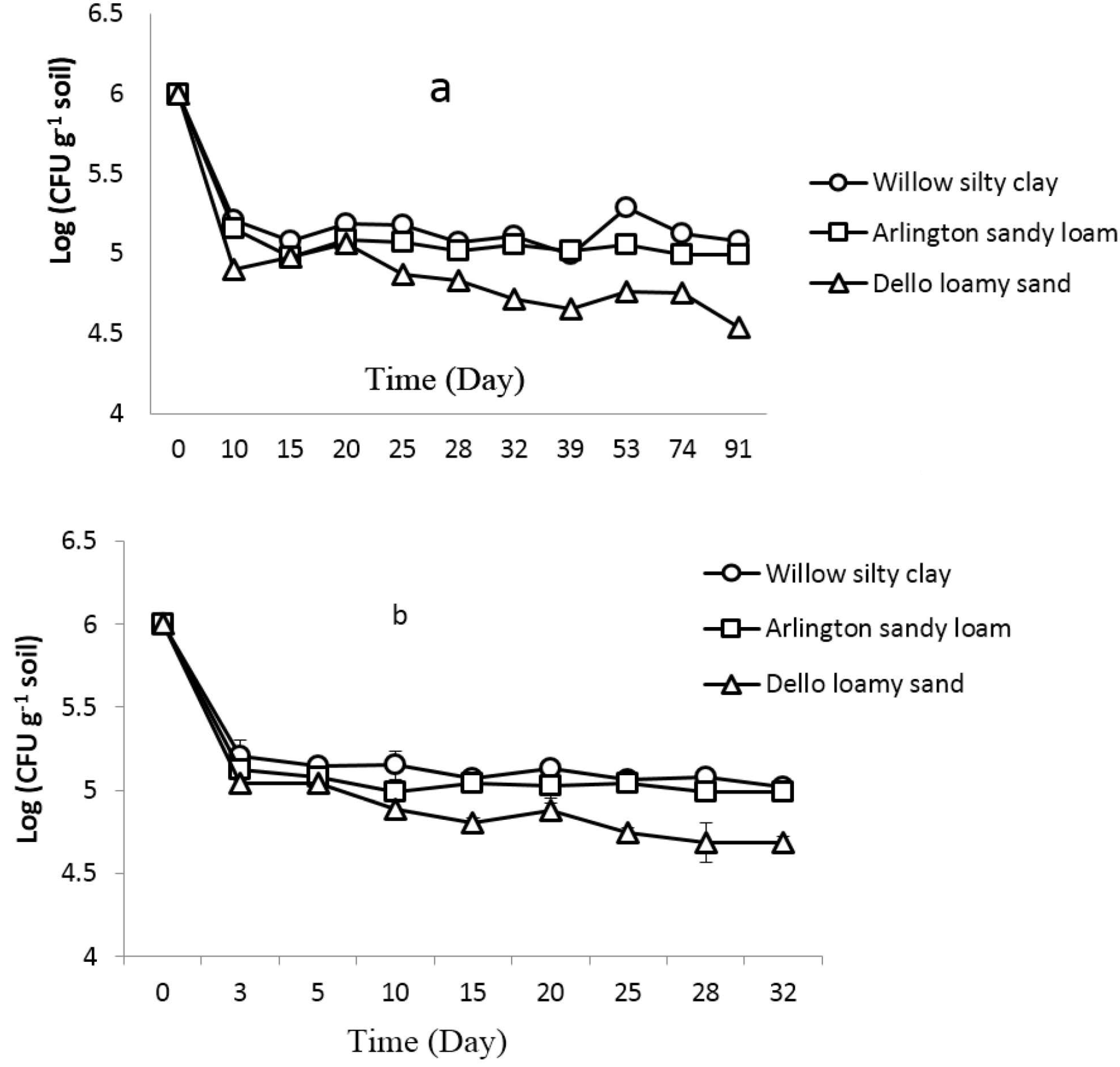
3.1. Abundance of B. subtilis B068150 in Cucumber Rhizosphere and Non-Rhizosphere

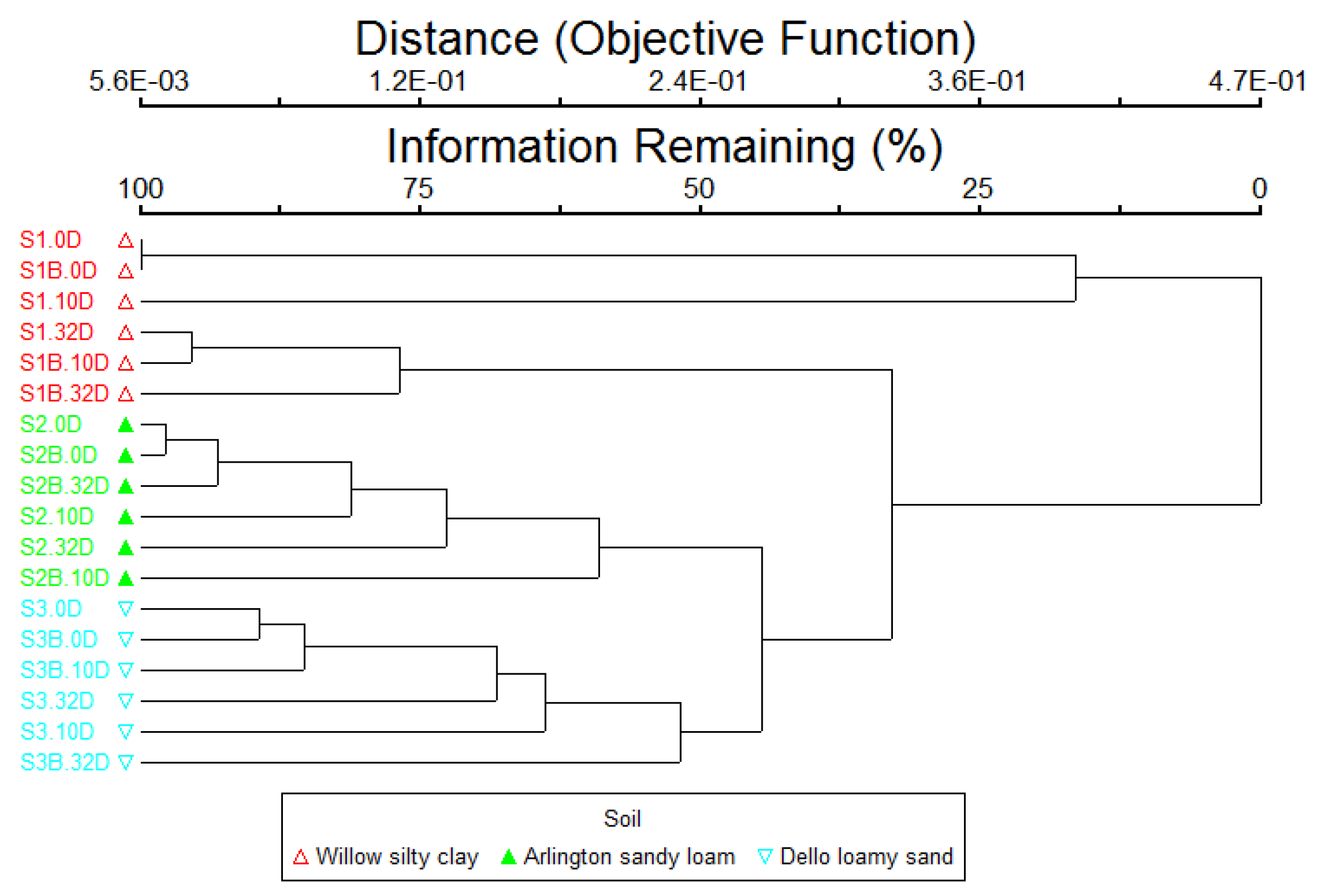
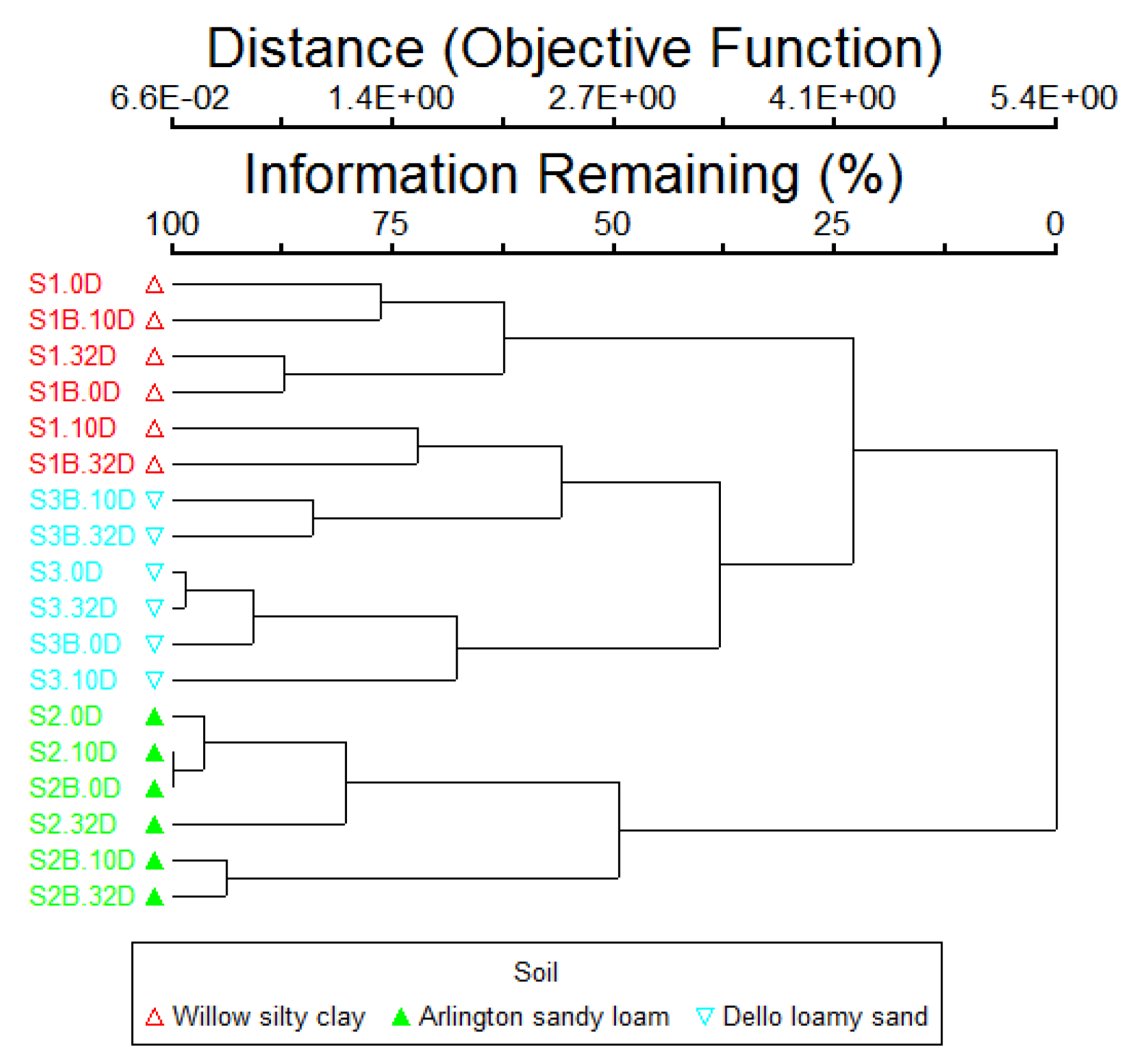
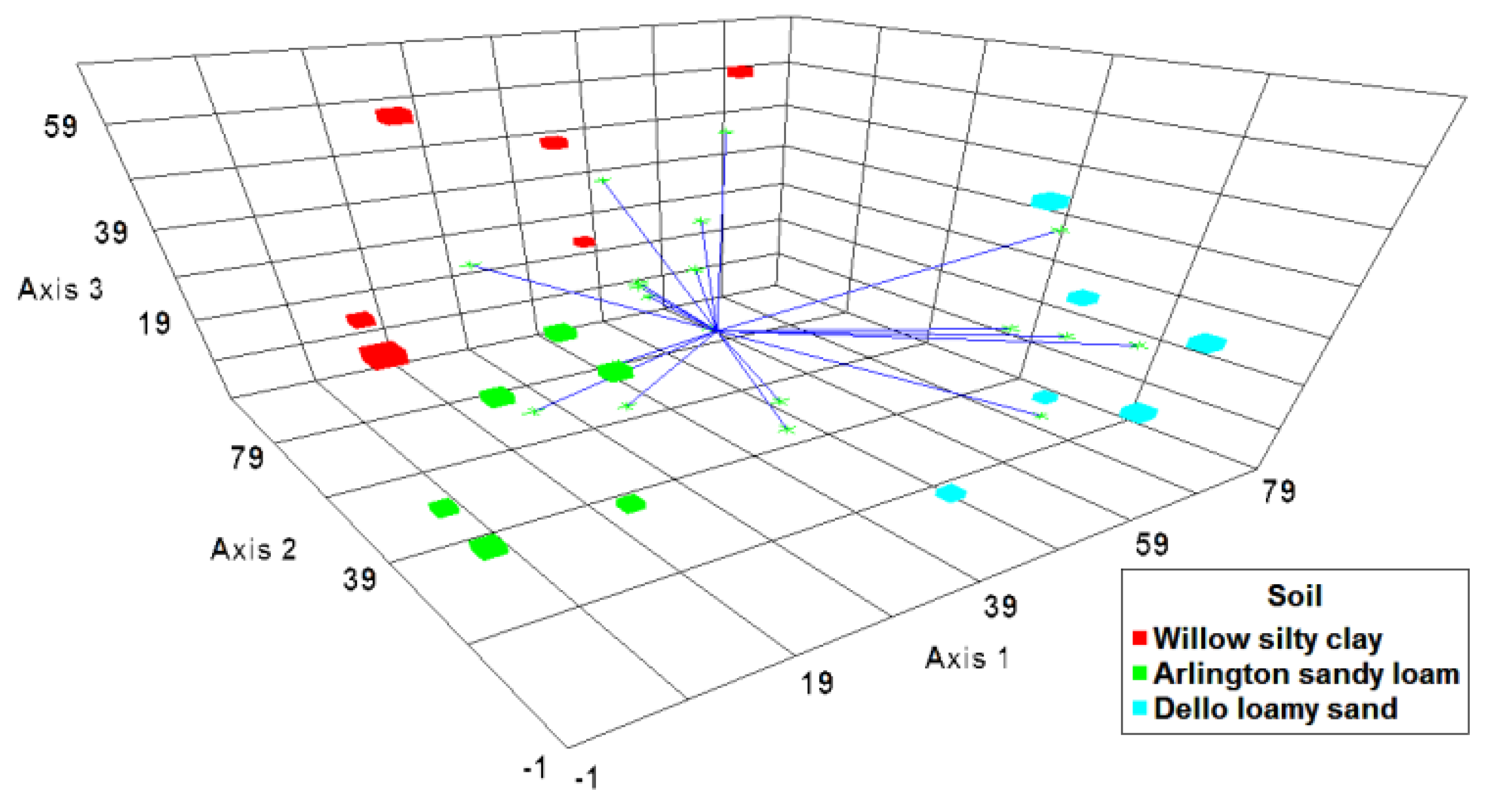
3.2. Changes in Bacterial Community Structure in Cucumber Rhizosphere with B. subtilis B068150


| Colonization in Clay | Colonization in Loam | Colonization in Sand | |
|---|---|---|---|
| Richness | −1.000 * (0.017) | −0.286 (0.815) | −0.109 (0.930) |
| Evenness | +0.685 (0.520) | +0.584 (0.603) | +0. 611 (0.582) |
| Shannon diversity | −1.000 ** (0.007) | −0.253 (0.837) | +0. 018 (0.988) |
| Simpson diversity | −0.999 * (0.034) | +0.411 (0.730) | −0.057 (0.964) |
| Band | The Most Similar Species | Similarity (%) | Phylum | Accesion # |
|---|---|---|---|---|
| 2 | Bacillus asahii | 98 | Firmicutes | KF387676.1 |
| 8 | Bacillus sp. | 100 | Firmicutes | AB748970.1 |
| 10 | Bacillus sp. | 100 | Firmicutes | KJ935909.2 |
| 11 | Bacillus megaterium | 99 | Firmicutes | KC485314.1 |
| 13 | Bacillus sp. | 99 | Firmicutes | KF747082.1 |
| 14 | Bacillus licheniformis | 100 | Firmicutes | HQ009796.1 |
| 15 | Bacillus sp. | 100 | Firmicutes | GQ487543.1 |
| 16 | Bacillus jeotgali | 99 | Firmicutes | HM854268.1 |
| 17 | Bacillus jeotgali | 100 | Firmicutes | HM854268.1 |
| 18 | Bacillus sp. | 100 | Firmicutes | HQ141380.1 |
| 3 | Pedobacter sp. | 99 | Bacteroidetes | KF817798.1 |
| 4 | Chryseobacterium sp. | 99 | Bacteroidetes | GU451187.1 |
| 12 | Chryseobacterium sp. | 100 | Bacteroidetes | AB581570.1 |
| 7 | Herbaspirillum sp. | 97 | Proteobacteria | EU549851.1 |
| 20 | Uncultured Sphingomonas sp. | 100 | Proteobacteria | LN568861.1 |
| 25 | Polyangium sp. | 100 | Proteobacteria | KJ611227.1 |
| 26 | Arthrobacter nicotianae | 100 | Actinobacteria | HM209738.1 |
| 1 | Uncultured bacterium | 100 | unclassified | LN571102.1 |
| 5 | Uncultured bacterium | 99 | unclassified | KJ956651.1 |
| 6 | Uncultured bacterium | 100 | unclassified | GQ866159.1 |
| 9 | Uncultured bacterium | 99 | unclassified | HM273912.1 |
| 19 | Uncultured bacterium | 100 | unclassified | KM207257.1 |
| 21 | Uncultured bacterium | 99 | unclassified | AM935657.1 |
| 22 | Uncultured bacterium | 99 | unclassified | KC605411.1 |
| 23 | Uncultured bacterium | 99 | unclassified | FN567976.1 |
| 24 | Uncultured bacterium | 99 | unclassified | FN567976.1 |
3.3. Fungal Community in Cucumber Rhizosphere

| Band | The Most Similar Species | Similarity (%) | Phylum | Accesion # |
|---|---|---|---|---|
| 3 | Rhizophydium sp. | 95 | Chytridiomycota | AY635821.1 |
| 12 | Uncultured Ceratobasidium | 99 | Basidiomycotina | EF154347.1 |
| 5 | Uncultured ascomycete | 100 | Ascomycota | FN390517.1 |
| 8 | Stilbella fimetaria | 99 | Ascomycota | FJ939395.1 |
| 10 | Uncultured ascomycete | 100 | Ascomycota | FN390419.1 |
| 11 | Microascus cirrosus | 100 | Ascomycota | KM222204.1 |
| 13 | Chaetomium sp. | 99 | Ascomycota | AB521039.1 |
| 14 | Stilbella fimetaria | 98 | Ascomycota | FJ939395.1 |
| 15 | Uncultured ascomycete | 99 | Ascomycota | FN390729.1 |
| 17 | Aspergillus ustus | 99 | Ascomycota | GQ856237.1 |
| 18 | Stilbella fimetaria | 98 | Ascomycota | FJ939395.1 |
| 19 | Aspergillus sp. | 100 | Ascomycota | EU795695.1 |
| 1 | Uncultured fungus | 100 | unclassified | KF192297.1 |
| 2 | Uncultured fungus | 99 | unclassified | AJ635526.1 |
| 4 | Uncultured fungus | 99 | unclassified | AJ635526.1 |
| 6 | Uncultured fungus | 99 | unclassified | EU657332.1| |
| 7 | Uncultured fungus | 100 | unclassified | AB534361.1 |
| 9 | Uncultured fungus | 100 | unclassified | EU657047.1 |
| 16 | Uncultured fungus | 100 | unclassified | AB534361.1 |
| 20 | Uncultured fungus | 100 | unclassified | EU795695.1 |


4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Becker, J.O.; Schwinn, F.J. Control of soil-borne pathogens with living bacteria and fungi: Status and outlook. Pest. Sci. 1993, 37, 355–363. [Google Scholar] [CrossRef]
- Minuto, A.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Control of soilborne pathogens of tomato using a commercial formulation of Streptomyces griseoviridis and solarization. Crop Protect. 2006, 25, 468–475. [Google Scholar] [CrossRef]
- Bailey, K.L.; Boyetchko, S.M.; Längle, T. Social and economic drivers shaping the future of biological control: A Canadian perspective on the factors affecting the development and use of microbial biopesticides. Biol. Control 2010, 52, 221–219. [Google Scholar] [CrossRef]
- Bailey, K.L.; Mupondwa, E.K. Developing microbial weed control products: Commercialization, biological, and technological considerations. In Handbook of Sustainable Weed Management; Singh, H.P., Batish, D.R., Kohli, R.K., Eds.; The Haworth Press Inc.: Binghamton, NY, USA, 2006; pp. 431–473. [Google Scholar]
- Véronique, E.H.; Sylvie, B.; Nadine, G.; Sébastien, A.; Claude, A.; Christian, S. Ecological fitness of the biocontrol agent Fusarium oxysporum Fo47 in soil and its impact on the soil microbial communities. FEMS Microbiol. Ecol. 2009, 68, 37–45. [Google Scholar]
- Timms-Wilson, T.; Kilshaw, K.; Bailey, M.J. Risk assessment for engineered bacteria used in biocontrol of fungal disease in agricultural crops. Plant Soil 2004, 266, 57–67. [Google Scholar] [CrossRef]
- Winding, A.; Binnerup, S.J.; Pritchard, H. Non-target effects of bacterial biological control agents suppressing root pathogenic fungi. FEMS Microbiol. Ecol. 2004, 47, 129–141. [Google Scholar] [CrossRef]
- Bae, Y.S.; Knudsen, G.R. Soil microbial biomass influence on growth and biocontrol efficacy of Trichoderma harzianum. Biol. Control 2005, 32, 236–242. [Google Scholar] [CrossRef]
- Pratibha, P.; Neera, K.; Sarita, S. Rhizosphere: Its structure, bacterial diversity and significance. Rev. Environ. Sci. Biotech. 2014, 13, 63–77. [Google Scholar]
- Van Bruggen, A.H.C.; Semenov, A.M. A new approach to the search for indicators of root disease suppression. Aust. Plant Pathol. 1999, 28, 4–10. [Google Scholar] [CrossRef]
- Van Bruggen, A.H.C.; Semenov, A.M. In search of biological indicators for soil health and disease suppression. Appl. Soil Ecol. 2000, 15, 13–24. [Google Scholar] [CrossRef]
- Garbeva, P.; van Veen, J.A.; van Elsas, J.D. Mocrobial diversity in soil: Selection of microbial populations by plant and soil type and implications for disease suppressiveness. Annu. Rev. Phytopathol. 2004, 42, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Van Bruggen, A.H.C.; Semenov, A.M.; van Diepeningen, A.D.; de Vos, O.J.; Blok, W.J. Relation between soil health, wave-like fluctuations in microbial populations, and soil-borne plant disease management. Eur. J. Plant Pathol. 2006, 115, 105–122. [Google Scholar] [CrossRef]
- Mazzola, M. Assessment and management of soil microbial community structure for disease suppression. Annu. Rev. Phytopathol. 2004, 42, 35–59. [Google Scholar] [CrossRef] [PubMed]
- Hiddink, G.A.; van Bruggen, A.H.C.; Termorshuizen, A.J.; Raaijmakers, J.M.; Semenov, A.V. Effect of organic management of soils on suppressiveness to Gaeumannomyces graminis var. tritici and its antagonist, Pseudomonas fluorescens. Eur. J. Plant Pathol. 2005, 113, 417–435. [Google Scholar] [CrossRef]
- Ibekwe, A.M.; Poss, J.A.; Grattan, S.R.; Grieve, C.M.; Suarez, D. Bacterial diversity in cucumber (Cucumis sativus) rhizosphere in response to salinity, soil pH, and boron. Soil Biol. Biochem. 2010, 42, 567–575. [Google Scholar] [CrossRef]
- Fang, M.; Kremer, R.J.; Motavalli, P.P.; Davis, G. Bacterial diversity of rhizospheres of non-transgenic and transgenic corn. Appl. Environ. Microbiol. 2005, 71, 4132–4136. [Google Scholar] [CrossRef] [PubMed]
- Grayston, S.J.; Wang, S.Q.; Campbell, C.D.; Edwards, A.C. Selective influence of plant species on microbial diversity in the rhizosphere. Soil Biol. Biochem. 1998, 30, 369–378. [Google Scholar] [CrossRef]
- Larkin, R.; Tavantzis, S.; Bernard, E.; Alyokhin, A.; Erich, S.; Gross, S. Compost and biological amendment effects on soil borne disease and soil microbial communities. Phytopathology 2008, 98, S86. [Google Scholar]
- Manici, L.M.; Caputo, F. Soil fungal communities as indicators for replanting new peach orchards in intensively cultivated areas. Eur. J. Agron. 2010, 33, 188–196. [Google Scholar] [CrossRef]
- Bai, Y.H.; Liang, J.S.; Liu, R.P.; Hu, C.Z.; Qu, J.H. Metagenomic analysis reveals microbial diversity and function in the rhizosphere soil of a constructed wetland. Environ. Tech. 2014, 35, 2521–2527. [Google Scholar] [CrossRef] [PubMed]
- Van Diepeningen, A.D.; Vos, O.J.; Zelenev, V.V.; Semenov, A.M.; van Bruggen, A.H.C. DGGE fragments oscillate with or counter to fluctuations of cultivable bacteria along wheat roots. Microb. Ecol. 2005, 50, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Grosch, R.; Junge, H.; Krebs, B.; Bochow, H. Use of Bacillus subtilis as a biocontrol agent. III. Influence of Bacillus subtilis on fungal root diseases and on yield in soil less culture. J. Plant Dis. Protect. 1999, 106, 568–580. [Google Scholar]
- Idriss, E.E.; Bochow, H.; Ross, H.; Borriss, R. Use of Bacillus subtilis as biocontrol agent. VI. Phytohormone-like action of culture filtrates prepared from plant growth-promoting Bacillus amyloliquefaciens FZB24, FZB42, FZB45 and Bacillus subtilis FZB37. J. Plant Dis. Protect. 2004, 111, 583–397. [Google Scholar]
- Koumoutsi, A.; Chen, X.H.; Vater, J.; Borriss, R. DegU and YczE positively regulate the synthesis of bacillomycin D by Bacillusamy-loliquefaciens strain FZB42. Appl. Environ. Microbiol. 2007, 73, 6953–6964. [Google Scholar] [CrossRef] [PubMed]
- Arguelles-Arias, A.; Ongena, M.; Halimi, B.; Lara, Y.; Brans, A.; Joris, B.; Fickers, P. Bacillusamy loliquefaciens GA1 asasource of potentanti biotics and other secondary metabolites for biocontrol of plant pathogens. Microbial Cell Fact. 2009, 8, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.P.; Dietel, K.; Rändler, M.; Schmid, M.; Junge, H.; Boriss, R.; Hartmann, A.; Grosch, R. Effects of Bacillusamy loliquefaciens FZB42 on lettuce growth and health under pathogen pressure and its impact on the rhizosphere bacterial community. PLoS ONE 2013, 8, e68818. [Google Scholar] [CrossRef] [PubMed]
- Baysala, Ö.; Çalışkanc, M.; Yeşilovaa, Ö. An inhibitory effect of a new Bacillus subtilis strain (EU07) against Fusarium oxysporum f. sp. radicis-lycopersici. Physiol. Mol. Plant Path. 2008, 73, 25–32. [Google Scholar] [CrossRef]
- Cazorla, F.M.; Romero, D.; Pérez-García, A.; Lugtenberg, B.J.J.; de Vicente, A.; Bloemberg, G. Isolation and characterization of antagonistic Bacillus subtilis strains from the avocado rhizoplane displaying biocontrol activity. J. Appl. Microbiol. 2007, 103, 1950–1959. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, D.K.; Johri, B.N. Interactions of Bacillus spp. and plants with special reference to induced systemic resistance (ISR). Microbiol. Res. 2009, 164, 493–513. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Kong, H.; Buyer, J.S.; Lakshman, D.K.; Lydon, J.; Kim, S.; Roberts, D.P. Isolation and partial characterization of Bacillus subtilis ME488 for suppression of soilborne pathogens of cucumber and pepper. Appl. Microbiol. Biotechnol. 2008, 80, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Hervás, A.; Landa, B.; Datnof, L.E.; Jiménez-Díaz, R.M. Effects of commercial and indigenous microorganisms on Fusarium wilt development in chickpea. Biol. Control 1998, 13, 166–176. [Google Scholar] [CrossRef]
- Rodgers, P.B. Potential of biological control organisms as a source of antifungal compounds for agrochemical and pharmaceutical product development. Pest. Sci. 1989, 27, 155–164. [Google Scholar] [CrossRef]
- Luo, Y.; Sun, L.F.; Zhu, Z.; Shen, Q. Identification and characterization of an anti-fungi Fusarium oxysporum f. sp cucumerium protease from the Bacillus subtilis strain N7. J. Microbiol. 2013, 51, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.H.; Zhang, R.F.; Wang, D.D.; Qiu, M.H.; Feng, H.C.; Zhang, N.; Shen, Q.R. Enhanced Control of Cucumber Wilt Disease by Bacillus amyloliquefaciens SQR9 by Altering the Regulation of Its DegU Phosphorylation. App. Environ. Microbiol. 2014, 80, 2941–2950. [Google Scholar] [CrossRef] [PubMed]
- Van Bruggen, A.H.C.; Semenov, A.M.; Zelenev, V.V.; Semenov, A.V.; Raaijmakers, J.M.; Sayler, R.; de Vos, O. Wave-like distribution patterns of gfp-marked Pseudomonas fluorescens along roots of wheat plants grown in two soils. Microb. Ecol. 2008, 55, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhang, Z.; Ling, N.; Yuan, Y.; Zhang, X.; Sun, B.; Shen, Q. Bacillus subtilis SQR9 can control Fusarium wilt in cucumber by colonizing plant roots. Biol. Fertil. Soils 2011, 47, 495–506. [Google Scholar] [CrossRef]
- Li, L.; Ma, J.; Yan, L.; Wang, Z.; Gao, T.; Wang, Q. Screening and partial characterization of Bacillus with potential applications in biocontrol of cucumber Fusarium wilt. Crop Protect. 2012, 35, 29–35. [Google Scholar] [CrossRef]
- Ma, J.; Ibekwe, A.M.; Yi, X.; Wang, H.Z.; Yamazaki, A.; Crowley, D.E.; Yang, C.H. Persistence of Escherichia coli O157:H7 and Its Mutants in Soils. PLoS ONE 2011, 6, e23191. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodfellow, M.D., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [PubMed]
- Smit, E.; Leeflang, P.; Glandorf, B.; van Elsas, J.D.; Wernars, K. Analysis of Fungal Diversity in the Wheat Rhizosphere by Sequencing of Cloned PCR-Amplified Genes Encoding 18S rRNA and Temperature Gradient Gel Electrophoresis. Appl. Environ. Microbiol. 1999, 65, 2614–2621. [Google Scholar] [PubMed]
- White, T.J.; Burns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phyologenetics. In PCR Protocols; Innes, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- McCune, B.; Mefford, M.J. PC-ORD for Windows. Multivariate Analysis of Ecological Data; Version 4.01; MjM Sofware: Gleneden Beach, OR, USA, 1999. [Google Scholar]
- Ibekwe, A.M.; Ma, J. Effects of fumigants on microbial diversity and persistence of E. coli O15:H7 in contrasting soil microcosms. Sci. Total Environ. 2011, 409, 3740–3748. [Google Scholar] [CrossRef] [PubMed]
- Shannon, A.E.; Weaver, W. The Mathematical Theory of Communication; University Illinois Press: Urbana, IL, USA, 1963. [Google Scholar]
- Duffy, B.K.; Weller, D.M. Biological control of take-all of wheat in the Pacific North-west of the USA using hypovirulent Gaeumannomyces graminis var. tritici, fluorescent Pseudomonads. J. Phytopathol. 1996, 144, 11–12. [Google Scholar] [CrossRef]
- Cavaglieri, L.R.; Andres, L.; Ibáñez, M.; Etcheverry, M.G. Rhizobacteria and their potential to control Fusarium verticillioides: Effect of maize bacterisation and inoculum density. Antonie van Leeuwenhoek 2005, 87, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Akiba, Y. Microbial ecology of Bacillus thuringiensis VI. Germination of Bacillus thuringiensis spores in the soil. Appl. Entomol. Zool. 1986, 21, 76–80. [Google Scholar]
- Pedersen, J.C.; Damgaard, P.H.; Eilenberg, J.; Hansen, B.M. Dispersal of Bacillus thuringiensis var. kurstaki in an experimental cabbage field. Can. J. Microbiol. 1995, 41, 118–125. [Google Scholar] [CrossRef]
- Van Elsas, J.D.; Dijkstra, A.F.; Govaert, J.M.; van Veen, J.A. Survival of Pseudomonas fluorescens and Bacillus subtilis introduced into two soils of different texture in field microplots. FEMS Microbiol. Ecol. 1986, 38, 151–160. [Google Scholar] [CrossRef]
- Backman, P.A.; Brannen, P.M.; Mahaffee, W.F. Plant response and disease control following seed inoculation with Bacillus subtilis. In Improving Plant Productivity with Rhizosphere Bacteria; Ryder, M.H., Stephens, P.M., Bowen, G.D., Eds.; CSIRO Division of Soils: Adclaidc, Australia, 1994; pp. 3–8. [Google Scholar]
- Gamalero, E.; Lingua, G.; Capr, F.G.; Fusconi, A.; Berta, G.; Lemanceau, P. Colonization pattern of primary tomato roots by Pseudomonas fluorescens A6RI characterized by dilution plating, flow cytometry, fluorescence, confocal and scanning electron microscopy. FEMS Microbiol. Ecol. 2004, 48, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhao, H.; Chen, S. Colonization of maize and rice plants by strain Bacillus megaterium C4. Curr. Microbiol. 2006, 52, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of Plant Growth-Promoting Bacteria for Biocontrol of Plant Diseases: Principles, Mechanisms of Action, and Future Prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [PubMed]
- Príncipe, A.; Alvarez, F.; Castro, M.G.; Zachi, L.; Fischer, S.E.; Mori, G.B.; Jofré, E. Biocontrol and PGPR features in native strains isolated from saline soils of Argentina. Curr. Microbiol. 2007, 55, 314–322. [Google Scholar]
- Domsch, K.H.; Jagnow, G.; Anderson, T.H. An ecological concept for the assessment of side effects of agrochemicals on soil microorganisms. Residue Rev. 1983, 86, 65–105. [Google Scholar]
- Kröber, M.; Wibberg, D.; Grosch, R.; Eikmeyer, F.; Verwaaijen, B.; Chowdhury, S.P.; Hartmann, A.; Pühler, A.; Schlüter, A. Effect of the strain Bacillus amyloliquefaciens FZB42 on the microbial community in the rhizosphere of lettuce under field conditions analyzed by whole metagenome sequencing. Front. Microbiol. 2014, 5, 252. [Google Scholar] [CrossRef] [PubMed]
- Bääth, E.; Anderson, T.H. Comparison of soil fungal/bacterial ratios in a pH gradient using physiological and PLFA-based techniques. Soil Biol. Biochem. 2003, 35, 955–963. [Google Scholar] [CrossRef]
- Borneman, J.; Triplett, E.W. Molecular microbial diversity in soils from eastern Amazonia: Evidence for unusual microorganisms and microbial population shifts associated with deforestation. Appl. Environ. Microbiol. 1997, 63, 2647–2653. [Google Scholar] [PubMed]
- Acea, M.J.; Moore, C.R.; Alexander, M. Survival and growth of bacteria introduced into soil. Soil Biol. Biochem. 1998, 20, 509–515. [Google Scholar] [CrossRef]
- Broeckling, C.D.; Broz, A.K.; Bergelson, J.; Manter, D.K.; Vivanco, J.M. Root exudates regulate soil fungal community composition and diversity. Appl. Environ. Microbiol. 2008, 74, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Fall, R.; Vivanco, J.M. Biocontrol of Bacillus subtilis against infection of arabidopsis roots by Pseudomonas syringae is facilitated by biofilm formation and surfactin production. Plant Physiol. 2004, 134, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Emmert, E.A.B.; Handelsman, J. Biocontrol of plant disease: A gram positive perspective. FEMS Microbiol. Lett. 1999, 171, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by fluorescent Pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Ahn, I.P.; Chung, H.S.; Lee, Y.H. Vegetative compatibility groups and pathogenicity among isolates of Fusarium oxysporum f. sp. cucumerinum. Plant Dis. 1997, 82, 244–246. [Google Scholar] [CrossRef]
- O’Brien, H.E.; Parrent, J.L.; Jackson, J.A.; Moncalvo, J.M.; Vilgalys, R. Fungal community analysis by large-scale sequencing of environmental samples. Appl. Environ. Microbiol. 2005, 71, 5544–5550. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.W.; Kim, B.K.; Kim, C.; Jung, H.S.; Kim, B.S.; Lee, J.H.; Chun, J. Assessment of soil fungal communities using pyrosequencing. J. Microbiol. 2010, 48, 284–289. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Ma, J.; Ibekwe, A.M.; Wang, Q.; Yang, C.-H. Cucumber Rhizosphere Microbial Community Response to Biocontrol Agent Bacillus subtilis B068150. Agriculture 2016, 6, 2. https://doi.org/10.3390/agriculture6010002
Li L, Ma J, Ibekwe AM, Wang Q, Yang C-H. Cucumber Rhizosphere Microbial Community Response to Biocontrol Agent Bacillus subtilis B068150. Agriculture. 2016; 6(1):2. https://doi.org/10.3390/agriculture6010002
Chicago/Turabian StyleLi, Lihua, Jincai Ma, A. Mark Ibekwe, Qi Wang, and Ching-Hong Yang. 2016. "Cucumber Rhizosphere Microbial Community Response to Biocontrol Agent Bacillus subtilis B068150" Agriculture 6, no. 1: 2. https://doi.org/10.3390/agriculture6010002
APA StyleLi, L., Ma, J., Ibekwe, A. M., Wang, Q., & Yang, C.-H. (2016). Cucumber Rhizosphere Microbial Community Response to Biocontrol Agent Bacillus subtilis B068150. Agriculture, 6(1), 2. https://doi.org/10.3390/agriculture6010002