Genetic Diversity of Lowbush Blueberry throughout the United States in Managed and Non-Managed Populations




Abstract
1. Introduction
2. Materials and Methods
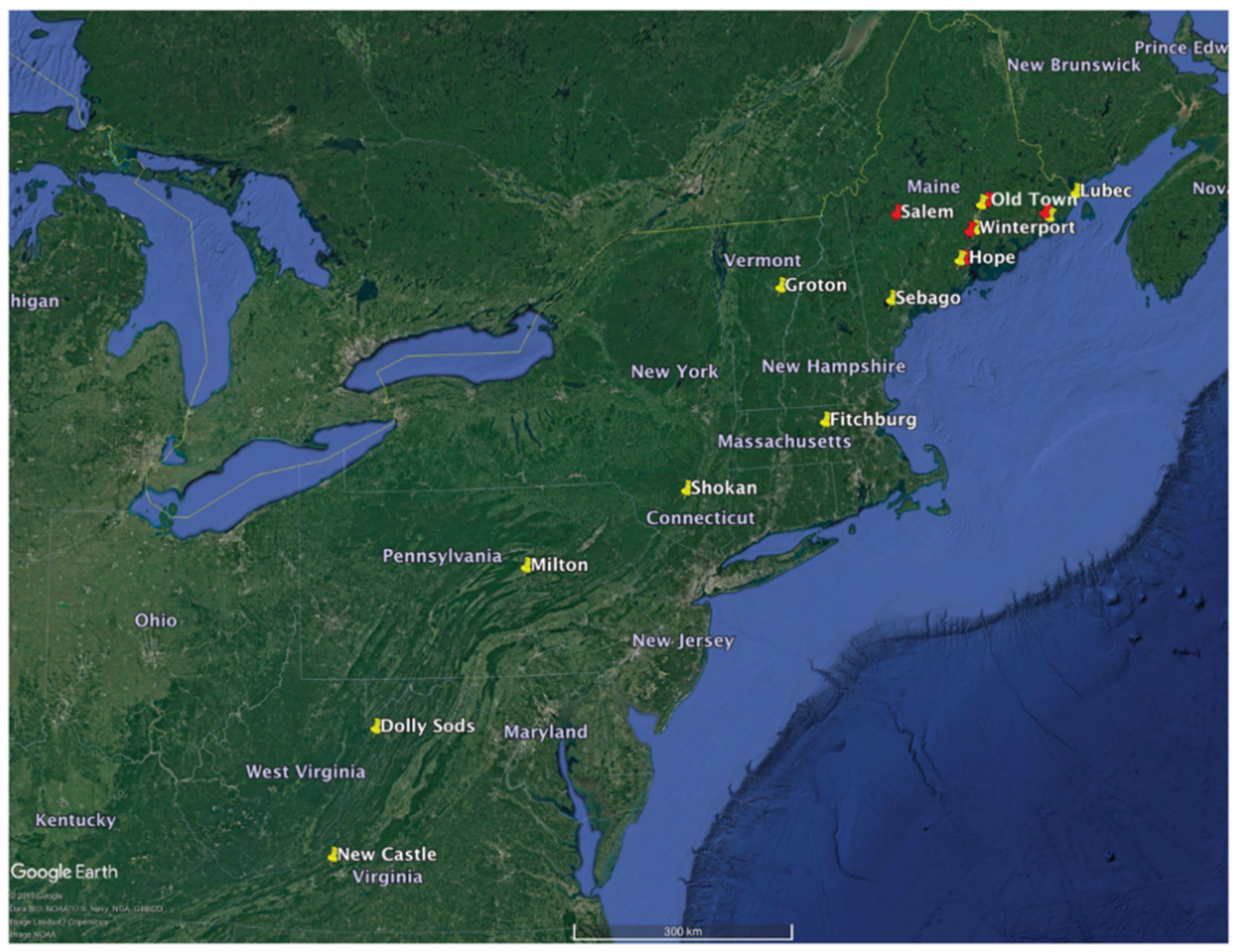
2.1. Plant Material
2.2. DNA Isolation and Amplification
2.3. Molecular Marker Analysis
2.4. Sequencing of Amplified Fragments
3. Results
3.1. Managed vs. Non-Managed Populations
3.2. Genetic Diversity throughout the Eastern United States
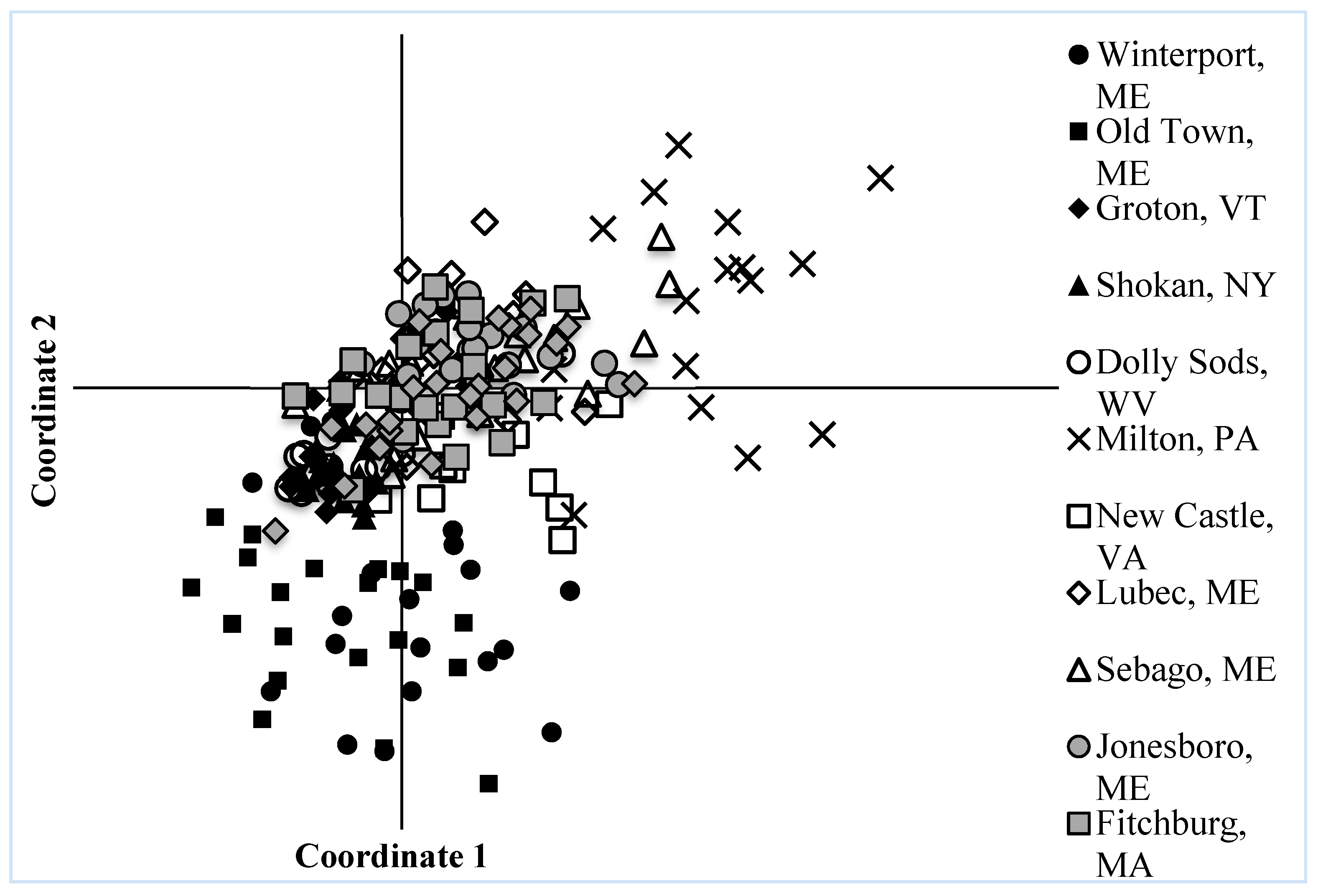
3.3. Spatial Analysis and Population Structure
3.4. Evaluation of Stress-Related Expressed Sequenced Tagged-Polymerase Chain Reaction (EST-PCR) Molecular Markers
3.5. Sequencing of Selected Polymorphic Bands
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bell, D.J.; Rowland, L.J.; Smagula, J.; Drummond, F.A. Recent Advances in the Biology and Genetics of Lowbush Blueberry. Tech. Bull. 2009, 203, 1–28. [Google Scholar]
- Vander Kloet, S.P. The Genus Vaccinium in North. America; Research Branch, Agriculture Canada: Ottawa, ON, Canada, 1988; 201p. [Google Scholar]
- Drummond, F.; Smagula, J.; Annis, S.; Yarborough, D. Organic Wild Blueberry Production. Tech. Bull. 2009, 852, 43. [Google Scholar]
- Yarborough, D. Factors contributing to the increase in productivity in the wild blueberry industry. Small Fruits Rev. 2004, 3, 33–43. [Google Scholar] [CrossRef]
- Barker, W.; Hall, I.; Aalders, L.; Wood, G. The lowbush blueberry industry in Eastern Canada. Econ. Bot. 1964, 18, 357–365. [Google Scholar] [CrossRef]
- Hepler, P.; Yarborough, D. Natural variability in yield of lowbush blueberries. HortScience 1991, 26, 245–246. [Google Scholar] [CrossRef]
- Cappiello, P.; Dunham, S. Seasonal variation in low-temperature tolerance of Vaccinium angustifolium Ait. HortScience 1994, 29, 302–304. [Google Scholar] [CrossRef]
- Bell, D.J.; Stommel, J.; Rowland, L.; Drummond, F. Yield variation among clones of lowbush blueberry as a function of kinship and self-compatibility. J. Am. Soc. Hort. Sci. 2010, 135, 259–270. [Google Scholar] [CrossRef]
- Penman, L.N.; Annis, S.L. Leaf and flower blight caused by Monilinia vaccinii-corymbosi on lowbush blueberry: Effects on yield and relationship to bud phenology. Phytopathology 2005, 95, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Burgher, K.L.; Jamieson, A.R.; Lu, X. Genetic relationships among lowbush blueberry genotypes as determined by randomly amplified polymorphic DNA analysis. J. Am. Soc. Hort. Sci. 2002, 127, 98–103. [Google Scholar] [CrossRef]
- Bell, D.J.; Rowland, L.; Drummond, F.; Polashock, J. Suitability of EST-PCR markers developed in highbush blueberry (Vaccinium corymbosum L.) for genetic fingerprinting and relationship studies in lowbush blueberry and related species. J. Am. Soc. Hort. Sci. 2008, 133, 701–707. [Google Scholar] [CrossRef]
- Bell, D.J.; Rowland, L.; Zhang, D.; Drummond, F. Spatial genetic structure of lowbush blueberry, Vaccinium angustifolium, in four fields in Maine. Botany 2009, 87, 932–946. [Google Scholar] [CrossRef]
- Bell, D.J.; Rowland, L.; Drummond, F. Does pollen ‘neighborhood’ affect berry yield in lowbush blueberry (Vaccinium angustifolium Ait)? Int. J. Fruit Sci. 2012, 12, 65–74. [Google Scholar] [CrossRef]
- Bell, D.J.; Drummond, F.; Rowland, L. Evidence of functional gender polymorphisms in a population of the hermaphroditic lowbush bluberry (Vaccinium angustifolium). Botany 2012, 90, 393–399. [Google Scholar] [CrossRef]
- Rowland, L.J.; Bell, D.J.; Alkharouf, N.; Bassil, N.V.; Drummond, F.; Beers, L.; Buck, E.; Finn, C.E.; Graham, J.; Mccallum, S.; et al. Generating genomic tools for blueberry improvement. Int. J. Fruit Sci. 2012, 12, 276–287. [Google Scholar] [CrossRef]
- Eckert, C.G.; Samis, K.E.; Lougheed, S.C. Genetic variation across species’ geographical ranges: The central–marginal hypothesis and beyond. Mol. Ecol. 2008, 17, 1170–1188. [Google Scholar] [CrossRef] [PubMed]
- Howe, G.T.; Hackett, W.P.; Furnier, G.R.; Klevorn, R.E. Photoperiodic responses of a northern and southern ecotype of black cottonwood. Physiol. Plant 1995, 93, 695–708. [Google Scholar] [CrossRef]
- Friedman, J.; Roelle, J.; Gaskin, J.; Pepper, A.; Manhart, J. Latitudinal variation in cold hardiness in introduced Tamarix and native Populus. Evol. Appl. 2008, 1, 598–607. [Google Scholar]
- Fernandez, I.J.; Schmitt, C.; Stancioff, E.; Birkel, S.D. Maine’s Climate Future: 2015 Update; University of Maine Climate Change Institute: Orono, ME, USA, 2015; 20p. [Google Scholar]
- Drummond, F.A.; Yarborough, D.E. Growing season effects on wild blueberry (Vaccinium angustifolium) in Maine and implications for management. Acta Hortic. 2012, 1017, 101–107. [Google Scholar] [CrossRef]
- Collins, J.A.; Drummond, F. Fertilizer and fungicides: Effects on wild blueberry growth, insect attack, and leaf spot disease incidence. In Proceedings of the North American Blueberry Research and Extension Workers Conference, Orono, ME, USA, 12–15 August 2018; 24p. [Google Scholar]
- Drummond, F.A.; Dibble, A.C.; Stubbs, C.; Bushmann, S.; Ascher, J.S.; Ryan, J. A Natural History of Change in Native Bees Associated with Lowbush Blueberry in Maine. Northeast. Nat. 2017, 24, 49–68. [Google Scholar] [CrossRef]
- Eck, P.; Childers, N.F. Blueberry Culture; Rutgers University Press: New Brunswick, NJ, USA, 1966; 400p. [Google Scholar]
- Hall, I.; Aalders, L. Blomidon Lowbush Blueberry. Can. J. Plant Sci. 1992, 62, 519–521. [Google Scholar] [CrossRef][Green Version]
- Hall, I. Genetic improvement of the lowbush blueberry, Vaccinium angustifolium. Can. J. Plant Sci. 1983, 63, 1091–1092. [Google Scholar] [CrossRef]
- Doebley, J.F.; Gaut, B.S.; Smith, B.D. The molecular genetics of crop domestication. Cell 2006, 127, 1309–1321. [Google Scholar] [CrossRef]
- van de Wouw, M.; Kik, C.; van Hintum, T.; van Treuren, R.; Visser, B. Genetic erosion in crops: Concept, research results and challenges. Plant Gen. Resour. 2009, 8, 1. [Google Scholar] [CrossRef]
- Aruna, M.; Ozias-Akins, P.; Austin, M.; Kochert, G. Genetic relatedness among rabbiteye blueberry (Vaccinium ashei) cultivars determined by DNA amplification using single primers of arbitrary sequence. Genome 1993, 36, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Loveless, M.D.; Hamrick, J.L. Ecological determinants of genetic structure in plant populations. Ann. Rev. Ecol. Syst. 1984, 15, 65–95. [Google Scholar] [CrossRef]
- Rowland, L.J.; Dhanaraj, A.L.; Polashock, J.J.; Arora, R. Utility of blueberry-derived EST-PCR primers in related Ericaceae species. HortScience 2003, 38, 1428–1432. [Google Scholar] [CrossRef]
- Groff, S.C.; Loftin, C.S.; Drummond, F.A.; Bushmann, S.; McGill, B. Spatial prediction of lowbush blueberry native bee pollinators in Maine, USA. Environ. Model. Softw. 2016, 79, 1–9. [Google Scholar] [CrossRef]
- Gleason, H.A.; Cronquist, A. Manual of Vascular Plants of Northeastern United States and Adjacent Canada, 2nd ed.; Botanical Garden: New York, NY, USA, 1991; 910p. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Darwish, O.; Rowland, L.J.; Alkharouf, N.W. BBGD454: A database for transcriptome analysis of blueberry using 454 sequences. Bioinformation 2013, 9, 883–886. [Google Scholar] [CrossRef][Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J. Exp. Bot. 2007, 58, 221–227. [Google Scholar] [CrossRef]
- Coker, J.S.; Davies, E. Selection of candidate housekeeping controls in tomato plants using EST data. BioTechniques 2003, 35, 740–748. [Google Scholar] [CrossRef] [PubMed]
- Rowland, L.J.; Mehra, S.; Dhanaraj, A.; Ogden, E.L.; Slovin, J.P.; Ehlenfeld, M.K. Development of EST-PCR markers for DNA fingerprinting and genetic relationship studies in blueberry (Vaccinium, section Cyanococcus). J. Am. Soc. Hort. Sci. 2003, 128, 682–690. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes. 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Simes, R.J. An improved Bonferroni procedure for multiple tests of significance. Biometrika 1986, 73, 751–754. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Zhen, Y.; Ungerer, M. Relaxed selection on the CBF/DREB1 regulatory genes and reduced freezing tolerance in the southern range of Arabidopsis thaliana. Mol. Biol. Evol. 2008, 25, 2547. [Google Scholar] [CrossRef]
- Holt, J.S.; Powles, S.B.; Holtum, J. Mechanisms and agronomic aspects of herbicide resistance. Ann. Rev. Plant Biol. 1993, 44, 203–229. [Google Scholar] [CrossRef]
- Yarborough, D.; Cote, J. Pre-and post-emergence applications of herbicides for control of resistant fine leaf sheep fescue in wild blueberry fields in Maine. In Proceedings of the North American Blueberry Research and Extension Workers Conference, Atlantic City, NJ, USA, 23–26 June 2014; 8p. [Google Scholar]
- Jensen, K.I.N.; Yarborough, D.E. An Overview of Weed Management in the Wild Lowbush Blueberry—Past and Present. Small Fruits Rev. 2004, 3, 229–255. [Google Scholar] [CrossRef]
- Yarborough, D. Wild blueberry culture in Maine. Univ. Maine Coop. Ext. Fact. Sheet. 2015. No. 220. Available online: https://extension.umaine.edu/blueberries/factsheets/production/wild-blueberry-culture-in-maine/ (accessed on 28 May 2019).
- DeGomez, T. Pruning lowbush blueberry fields. Univ. of Maine Cooop. Ext. Wild Blueberry Fact. Sheet. 1988. No. 229. Available online: https://extension.umaine.edu/blueberries/factsheets/production/pruning-lowbush-blueberry-fields/ (accessed on 28 May 2019).
- Smith, D.W.; Hilton, R.J. The Comparative Effects of Pruning by Burning or Clipping on Lowbush Blueberries in North-Eastern Ontario. J. Appl. Ecol. 1971, 8, 781. [Google Scholar] [CrossRef]
- Hanson, E.J.; Ismail, A.A.; Metzger, H.B. A Cost Analysis of Pruning Procedures in Lowbush Blueberry Production; Life Sciences and Agriculture Experiment Station: Ithaca, NY, USA, 1982; Bulletin 780. [Google Scholar]
- Rao, V.R.; Hodgkin, T. Genetic diversity and conservation and utilization of plant genetic resources. Plant Cell Tissue Organ. Culture 2002, 68, 1–19. [Google Scholar]
- Debnath, S.C. Development of ISSR markers for genetic diversity studies in Vaccinium angustifolium. Nordic J. Botany 2009, 27, 141–148. [Google Scholar] [CrossRef]
- Hewitt, G. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Zoratti, L.; Palmieri, L.; Jaakola, L.; Häggman, H. Genetic diversity and population structure of an important wild berry crop. AoB Plants 2015, 7, plv117. [Google Scholar] [CrossRef]
- Hamrick, J.L.; Godt, M.J.W.; Sherman-Broyles, S.L. Factors influencing levels of genetic diversity in woody plant species. New For. 1992, 6, 95–124. [Google Scholar] [CrossRef]
- Tailor, S.; Bykova, N.V.; Igamberdiev, A.U.; Debnath, S.C. Structural pattern and genetic diversity in blueberry (Vaccinium) clones and cultivars using EST-PCR and microsattelite markers. Genet. Resour. Crop. Evol. 2017, 64, 2071–2081. [Google Scholar] [CrossRef]
- Willi, Y.; Van Buskirk, J. Genomic compatibility occurs over a wide range of parental genetic similarity in an outcrossing Plant. Proc. R. Soc. 2005, 272, 1533–1538. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′ to 3′) | Tm (°C) | Annealing Temperature (°C) |
|---|---|---|---|
| 02675 | AAGGAAGGGGGAGGGTTTAT | 58 | 49 |
| AAAAAGGGGCACAAAGAAGAA | 54 | ||
| 00125 | AGTAGGGGACACAGCCACAC | 62 | 55 |
| TGGCAGAGGGTAGAACTTGC | 60 | ||
| 00064 | CACAGTTTTGACGGTGATGG | 56 | 51 |
| TGATTGCTGCACCAAGACTC | 58 | ||
| CO | AAACTACCCGATGTCGATGC | 57 | 49 |
| TCACAAAAACGATGGAACGA | 54 | ||
| 00313 | CAGCACAAATTGCAGAGCAT | 56 | 50 |
| GCATGGGAAAGGAATTCTGA | 55 | ||
| NA799 | TTTACCTCCCTTTGCCACAC | 57 | 52 |
| GGAAATCCCACAGCTCAGAA | 57 | ||
| CA448F | GTGGGCAGAGTGAGGAAGAG | 60 | 53 |
| ACACAAACCAGGGAGAAACG | 58 | ||
| CA15F | ACCAAAGCTGAGCAACCAAT | 57 | 52 |
| GTCTGCCATGAAAACCCAAC | 57 | ||
| CA65F | GTCGAATCCGAAGCCTTCTC | 58 | 48 |
| AAACAGCAAATTCCCAATCG | 53 | ||
| CA1463 | GAAGATGTCGTGGAGGTGGT | 59 | 51 |
| TAATGCGGGTTGATGTAGCA | 56 | ||
| CA1423 | TCATAGCCAATACACTCGAACC | 56 | 51 |
| GCCCCACCTTAGCAAATC | 56 | ||
| CA1785L | CACCACCACTGCGTACACC | 62 | 50 |
| GCATGAGCCGAACATAATCA | 55 | ||
| EST133 | AAACAATCCACCAATCAACTTGT | 54 | 49 |
| CCTCTCCACAGTCCGATCAA | 59 | ||
| EST193 | GAGGGATTCAGCACGAAGAG | 58 | 50 |
| CAACATCATCAACCCCAACA | 55 | ||
| EST248 | TGGAGACTGGAGTGATGCAA | 58 | 49 |
| AAGTGCATTAAGCATCCGAAA | 54 | ||
| EST1029 | GAAGTTTTCCGTTCTCTGCAA | 55 | 50 |
| CTGCAGCTAGGACCGAAGAG | 60 |
| Managed | Non-Managed | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Hope | Jonesboro | Old Town | Winterport | Hope | Jonesboro | Old Town | Winterport | ||
| Non- Managed | Winterport | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | |
| Old Town | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | |||
| Jonesboro | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | ||||
| Hope | 0.0001 | 0.0001 | 0.0001 | 0.0001 | |||||
| Managed | Winterport | 0.0001 | 0.0056 (ns) | 0.0001 | |||||
| Old Town | 0.0001 | 0.0023 † | |||||||
| Jonesboro | 0.0001 | ||||||||
| Hope | |||||||||
| Source | df | SS | MS | F | p |
|---|---|---|---|---|---|
| Location | 3 | 7.1233 | 2.3744 | 16.299 | 0.0002 |
| Management | 1 | 3.7856 | 3.7856 | 25.986 | 0.0002 |
| Interaction | 3 | 3.1031 | 1.0344 | 7.1004 | 0.0002 |
| Residual | 152 | 22.143 | 0.14568 | ||
| Total | 159 | 36.155 |
| Source | df | SS | MS | Est. Var. | % |
|---|---|---|---|---|---|
| Among Pops | 16 | 1482.821 | 92.676 | 4.063 | 25% |
| Within Pops | 321 | 3878.995 | 12.084 | 12.084 | 75% |
| Total | 337 | 5361.817 | 16.147 | 100% | |
| Stat | Value | p | |||
| PhiPT | 0.252 | 0.0001 | |||
| Hope, ME | Hope, ME (Managed) | Fitchburg, MA | Jonesboro, ME | Sebago, ME | Salem, ME (Managed) | Old Town, ME (Managed) | Winterport, ME (Managed) | Jonesboro, ME (Managed) | Lubec, ME | New Castle, VA | Milton, PA | Dolly Sods, WV | Shokan, NY | Groton, VT | Old Town, ME | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Winterport, ME | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 |
| Old Town, ME | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | |
| Groton, VT | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0004 | 0.0026 | ||
| Shokan, NY | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0007 | |||
| Dolly Sods, WV | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | ||||
| Milton, PA | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | |||||
| New Castle, VA | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | ||||||
| Lubec, ME | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0005 | 0.2281 | |||||||
| Jonesboro, ME (Managed) | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0025 | 0.0037 | ||||||||
| Winterport, ME (Managed) | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | |||||||||
| Old Town, ME (Managed) | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | ||||||||||
| Salem, ME (Managed) | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | |||||||||||
| Sebago, ME | 0.0001 | 0.0001 | 0.0001 | 0.0001 | ||||||||||||
| Jonesboro, ME | 0.0001 | 0.0001 | 0.0001 | |||||||||||||
| Fitchburg, MA | 0.0001 | 0.0001 | ||||||||||||||
| Hope, ME (Managed) | 0.0001 | |||||||||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beers, L.; Rowland, L.J.; Drummond, F. Genetic Diversity of Lowbush Blueberry throughout the United States in Managed and Non-Managed Populations. Agriculture 2019, 9, 113. https://doi.org/10.3390/agriculture9060113
Beers L, Rowland LJ, Drummond F. Genetic Diversity of Lowbush Blueberry throughout the United States in Managed and Non-Managed Populations. Agriculture. 2019; 9(6):113. https://doi.org/10.3390/agriculture9060113
Chicago/Turabian StyleBeers, Lee, Lisa J. Rowland, and Francis Drummond. 2019. "Genetic Diversity of Lowbush Blueberry throughout the United States in Managed and Non-Managed Populations" Agriculture 9, no. 6: 113. https://doi.org/10.3390/agriculture9060113
APA StyleBeers, L., Rowland, L. J., & Drummond, F. (2019). Genetic Diversity of Lowbush Blueberry throughout the United States in Managed and Non-Managed Populations. Agriculture, 9(6), 113. https://doi.org/10.3390/agriculture9060113