
Transcriptome Analysis of Cultured Limbal Epithelial Cells on an Intact Amniotic Membrane following Hypothermic Storage in Optisol-GS

Abstract
:1. Introduction
2. Results
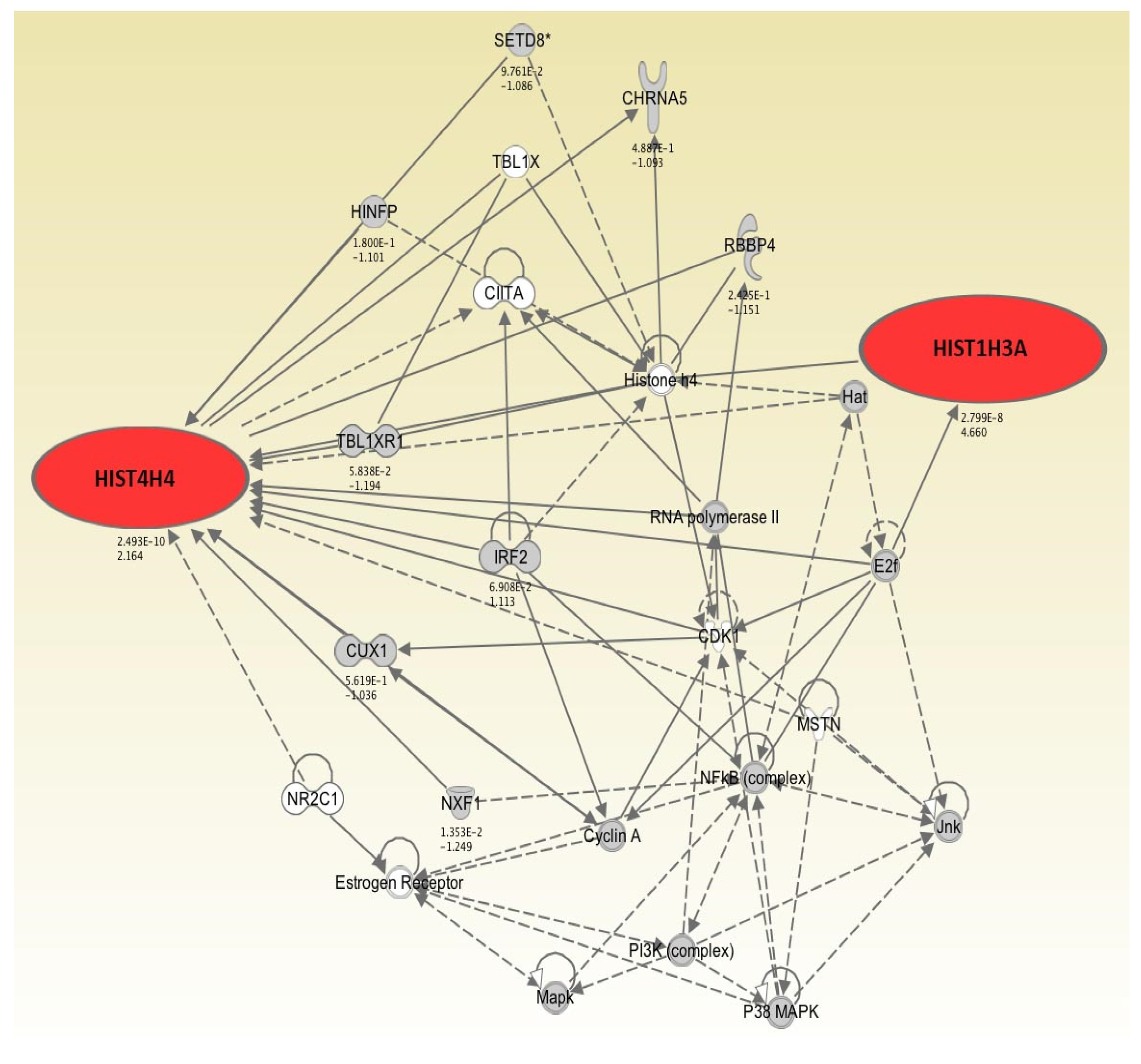
2.1. Genes Exhibiting Higher Levels of Expression Following 2, 4, and 7 Days of Hypothermic Storage Compared to Control
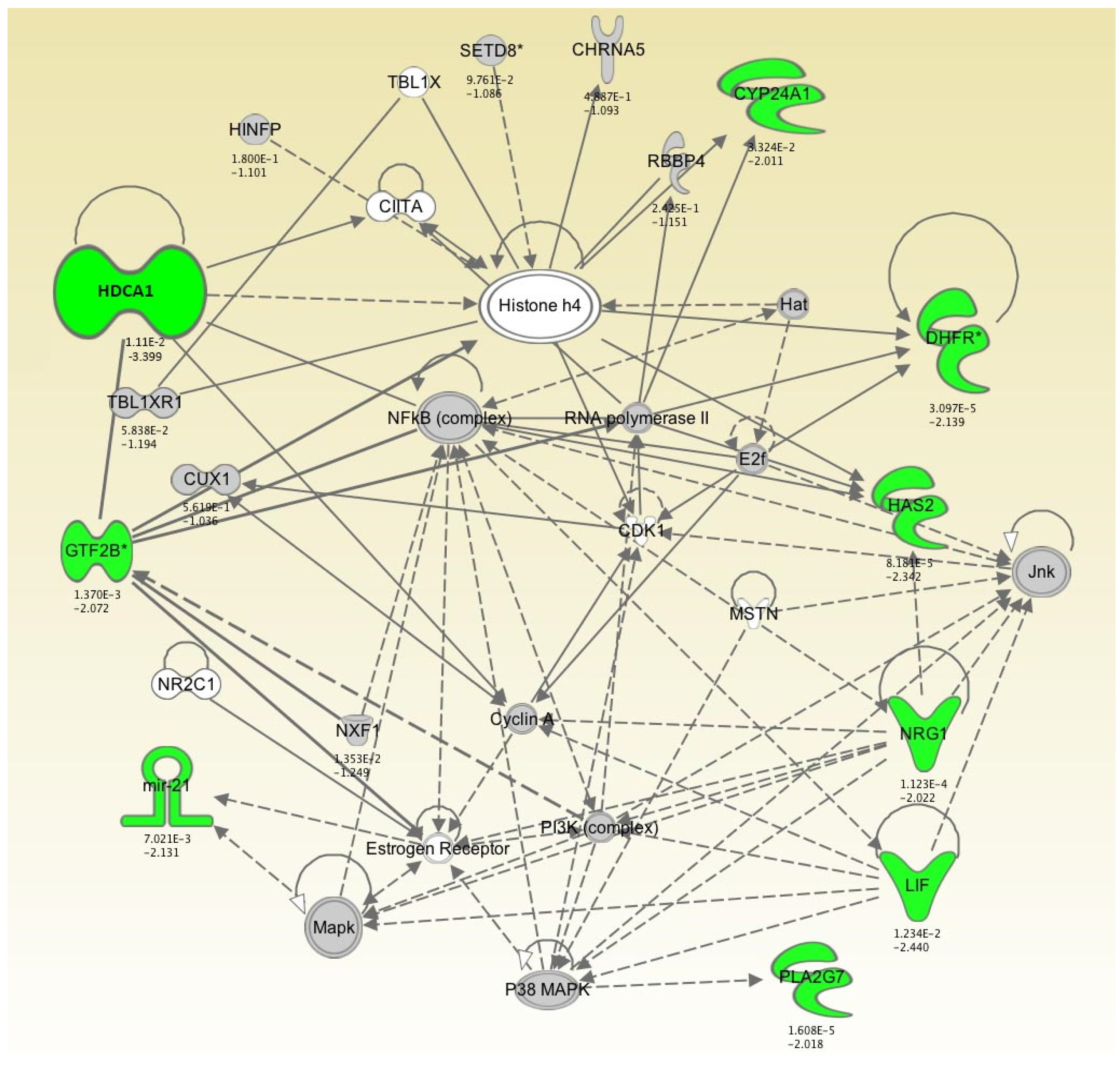
2.2. Genes Exhibiting Lower Levels of Expression Following 2, 4, and 7 Days of Hypothermic Storage Compared to Control
3. Discussion
4. Experimental Section
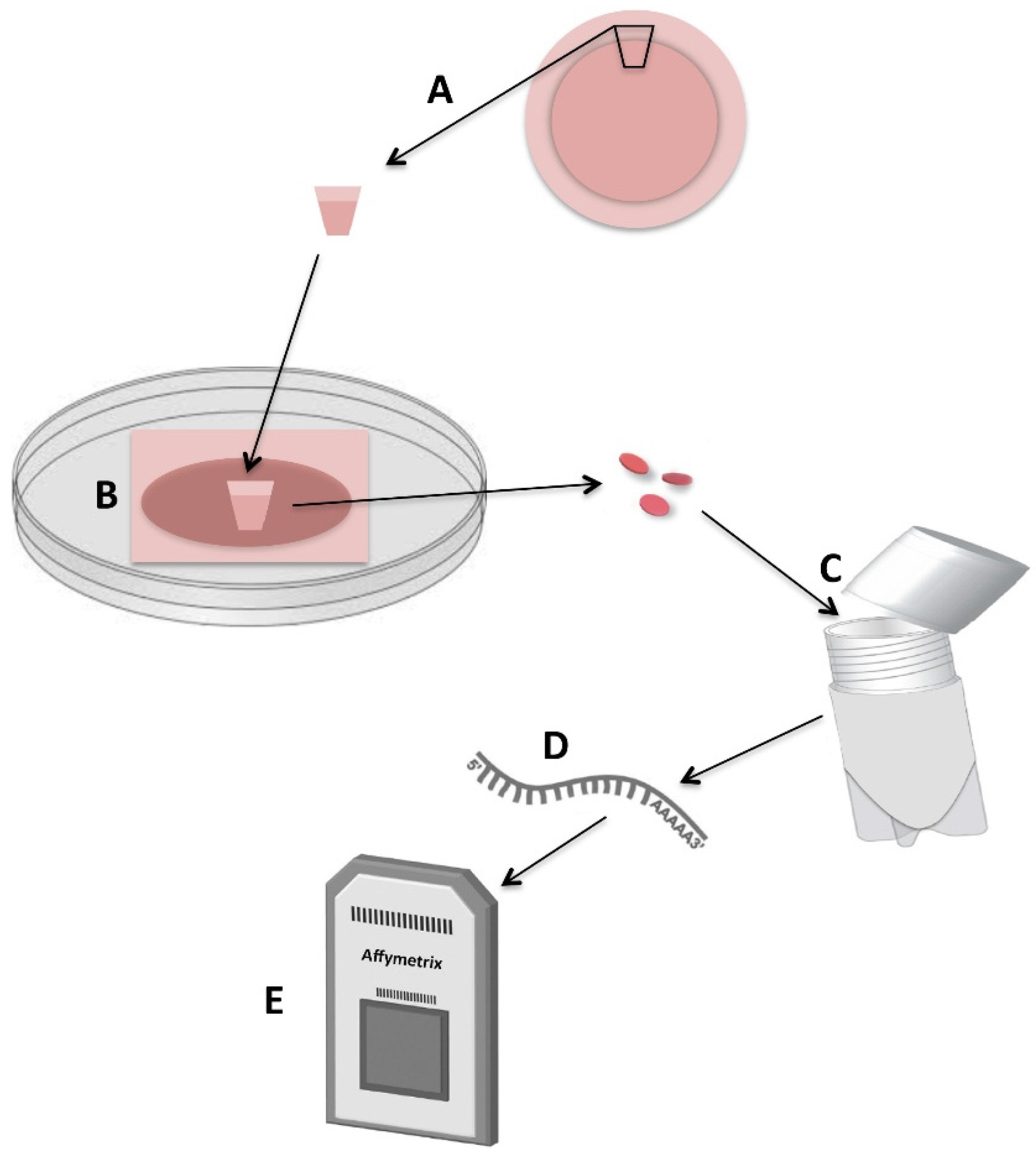
4.1. Human Tissue Preparation
4.2. Human Limbal Explant Cultures on Intact Amniotic Membranes
4.3. Hypothermic Storage of Cultured Human Limbal Epithelial Cells in Optisol-GS
4.4. RNA Isolation
4.5. Microarray Analysis
4.6. Bioinformatic Analysis
Acknowledgments
Author Contributions
Conflicts of interest
References
- Schermer, A.; Galvin, S.; Sun, T.T. Differentiation-related expression of a major 64K corneal keratin in vivo and in culture suggests limbal location of corneal epithelial stem cells. J. Cell Biol. 1986, 103, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Thoft, R.A.; Friend, J. The X, Y, Z hypothesis of corneal epithelial maintenance. Investig. Ophthalmol. Vis. Sci. 1983, 24, 1442–1443. [Google Scholar] [PubMed]
- Ahmad, S.; Osei-Bempong, C.; Dana, R.; Jurkunas, U. The culture and transplantation of human limbal stem cells. J. Cell Physiol. 2010, 225, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Oie, Y.; Nishida, K. Translational research on ocular surface reconstruction using oral mucosal epithelial cell sheets. Cornea 2014, 33, S47–S52. [Google Scholar] [CrossRef] [PubMed]
- Vasania, V.S.; Hari, A.; Tandon, R.; Shah, S.; Haldipurkar, S.; Shah, S.; Sachan, S.; Viswanathan, C. Transplantation of autologous ex vivo expanded human conjunctival epithelial cells for treatment of pterygia: A prospective open-label single arm multicentric clinical trial. J. Ophthalmic Vis. Res. 2014, 9, 407–416. [Google Scholar] [PubMed]
- Raeder, S.; Utheim, T.P.; Utheim, O.A.; Nicolaissen, B.; Roald, B.; Cai, Y.; Haug, K.; Kvalheim, A.; Messelt, E.B.; Drolsum, L.; et al. Effects of organ culture and optisol-GS storage on structural integrity, phenotypes, and apoptosis in cultured corneal epithelium. Invest. Ophthalmol. Vis. Sci. 2007, 48, 5484–5493. [Google Scholar] [CrossRef] [PubMed]
- Utheim, T.P.; Utheim, T.P.; Raeder, S.; Eidet, J.; Stormo, C.; de la Paz, M.; Utheim, O.A. Storage of cultured human limbal epithelial cells in Optisol-GS at 23 °C versus 5 °C. Invest. Ophthalmol. Vis. Sci. 2009, 50, 1778–1779. [Google Scholar]
- Pels, E.; Beele, H.; Claerhout, I. Eye bank issues: II. Preservation techniques: Warm versus cold storage. Int. Ophthalmol. 2008, 28, 155–163. [Google Scholar] [PubMed]
- Abrahamse, S.L.; van Runnard Heimel, P.; Hartman, R.J.; Chamuleau, R.A.; van Gulik, T.M. Induction of necrosis and DNA fragmentation during hypothermic preservation of hepatocytes in UW, HTK, and Celsior solutions. Cell Transplant. 2003, 12, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Komuro, A.; Hodge, D.O.; Gores, G.J.; Bourne, W.M. Cell death during corneal storage at 4 °C. Invest. Ophthalmol. Vis. Sci. 1999, 40, 2827–2832. [Google Scholar]
- Rauen, U.; de Groot, H. Mammalian cell injury induced by hypothermia—The emerging role for reactive oxygen species. Biol. Chem. 2002, 383, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Fitton, T.P.; Wei, C.; Lin, R.; Bethea, B.T.; Barreiro, C.J.; Amado, L.; Gage, F.; Hare, J.; Baumgartner, W.A.; Conte, J.V. Impact of 24 h continuous hypothermic perfusion on heart preservation by assessment of oxidative stress. Clin. Transplant. 2004, 18, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Utheim, T.P.; Raeder, S.; Olstad, O.K.; Utheim, O.A.; de La Paz, M.; Cheng, R.; Huynh, T.T.; Messelt, E.; Roald, B.; Lyberg, T. Comparison of the histology, gene expression profile, and phenotype of cultured human limbal epithelial cells from different limbal regions. Invest. Ophthalmol. Vis. Sci. 2009, 50, 5165–5172. [Google Scholar] [CrossRef] [PubMed]
- Frausto, R.F.; Wang, C.; Aldave, A.J. Transcriptome analysis of the human corneal endothelium. Investig. Ophthalmol. Vis. Sci. 2014, 55, 7821–7830. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Li, X.; Qian, S.W.; Guo, L.; Huang, H.Y.; He, Q.; Liu, Y.; Ma, C.G.; Tang, Q.Q. Transcriptional activation of histone H4 by C/EBPβ during the mitotic clonal expansion of 3T3-L1 adipocyte differentiation. Mol. Biol. Cell 2011, 22, 2165–2174. [Google Scholar] [CrossRef] [PubMed]
- Bongiorno-Borbone, L.; de Cola, A.; Barcaroli, D.; Knight, R.A.; di Ilio, C.; Melino, G.; de Laurenzi, V. FLASH degradation in response to UV-C results in histone locus bodies disruption and cell-cycle arrest. Oncogene 2010, 29, 802–810. [Google Scholar] [CrossRef] [PubMed]
- Kratzmeier, M.; Albig, W.; Meergans, T.; Doenecke, D. Changes in the protein pattern of H1 histones associated with apoptotic DNA fragmentation. Biochem. J. 1999, 337, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Sokol, A.; Kwiatkowska, A.; Jerzmanowski, A.; Prymakowska-Bosak, M. Up-regulation of stress-inducible genes in tobacco and arabidopsis cells in response to abiotic stresses and aba treatment correlates with dynamic changes in histone H3 and H4 modifications. Planta 2007, 227, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Kaygun, H.; Marzluff, W.F. Translation termination is involved in histone mRNA degradation when DNA replication is inhibited. Mol. Cell Biol. 2005, 25, 6879–6888. [Google Scholar] [CrossRef] [PubMed]
- Strahl, B.D.; Allis, C.D. The language of covalent histone modifications. Nature 2000, 403, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Turner, B.M. Histone acetylation and an epigenetic code. Bioessays 2000, 22, 836–845. [Google Scholar] [CrossRef]
- Fullgrabe, J.; Hajji, N.; Joseph, B. Cracking the death code: Apoptosis-related histone modifications. Cell Death Differ. 2010, 17, 1238–1243. [Google Scholar] [CrossRef] [PubMed]
- Lehrmann, H.; Pritchard, L.L.; Harel-Bellan, A. Histone acetyltransferases and deacetylases in the control of cell proliferation and differentiation. Adv. Cancer Res. 2002, 86, 41–65. [Google Scholar] [PubMed]
- Mai, A.; Massa, S.; Rotili, D.; Cerbara, I.; Valente, S.; Pezzi, R.; Simeoni, S.; Ragno, R. Histone deacetylation in epigenetics: An attractive target for anticancer therapy. Med. Res. Rev. 2005, 25, 261–309. [Google Scholar] [CrossRef] [PubMed]
- Gui, C.Y.; Ngo, L.; Xu, W.S.; Richon, V.M.; Marks, P.A. Histone deacetylase (HDAC) inhibitor activation of p21WAF1 involves changes in promoter-associated proteins, including HDAC1. Proc. Natl. Acad. Sci. USA 2004, 101, 1241–1246. [Google Scholar] [CrossRef] [PubMed]
- Bolden, J.E.; Peart, M.J.; Johnstone, R.W. Anticancer activities of histone deacetylase inhibitors. Nat. Rev. Drug Discov. 2006, 5, 769–784. [Google Scholar] [CrossRef] [PubMed]
- Marks, P.A.; Dokmanovic, M. Histone deacetylase inhibitors: Discovery and development as anticancer agents. Expert. Opin. Investig. Drugs 2005, 14, 1497–1511. [Google Scholar] [CrossRef] [PubMed]
- Rosato, R.R.; Grant, S. Histone deacetylase inhibitors: Insights into mechanisms of lethality. Expert. Opin. Ther. Targets 2005, 9, 809–824. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Wang, Y.; Yang, L.; Wang, Y.; Chen, P.; Wang, Y.; Dong, X.; Xie, L. Histone deacetylase inhibitors blocked activation and caused senescence of corneal stromal cells. Mol. Vis. 2008, 14, 2556–2565. [Google Scholar] [PubMed]
- Hammond, S.M. An overview of microRNAs. Adv. Drug Deliv. Rev. 2015, 87, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Asangani, I.A.; Rasheed, S.A.; Nikolova, D.A.; Leupold, J.H.; Colburn, N.H.; Post, S.; Allgayer, H. MicroRNA-21 (miR-21) post-transcriptionally downregulates tumor suppressor Pdcd4 and stimulates invasion, intravasation and metastasis in colorectal cancer. Oncogene 2008, 27, 2128–2136. [Google Scholar] [CrossRef] [PubMed]
- Kadera, B.E.; Li, L.; Toste, P.A.; Wu, N.; Adams, C.; Dawson, D.W.; Donahue, T.R. MicroRNA-21 in pancreatic ductal adenocarcinoma tumor-associated fibroblasts promotes metastasis. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, N.; Mandusic, V.; Stanojevic, B.; Lukic, S.; Todorovic, L.; Roganovic, J.; Dimitrijevic, B. The difference in miR-21 expression levels between invasive and non-invasive breast cancers emphasizes its role in breast cancer invasion. Med. Oncol. 2014, 31, 867. [Google Scholar] [CrossRef] [PubMed]
- Harmalkar, M.; Upraity, S.; Kazi, S.; Shirsat, N.V. Tamoxifen-induced cell death of malignant glioma cells is brought about by oxidative-stress-mediated alterations in the expression of BCL2 family members and is enhanced on miR-21 inhibition. J. Mol. Neurosci. 2015, 57, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Meller, D.; Pires, R.T.; Tseng, S.C. Ex vivo preservation and expansion of human limbal epithelial stem cells on amniotic membrane cultures. Br. J. Ophthalmol. 2002, 86, 463–471. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Symbol | 2 Days vs. Ctr | 4 Days vs. Ctr | 7 Days vs. Ctr | |||
|---|---|---|---|---|---|---|
| P-value | Fold Change | P-value | Fold Change | P-value | Fold Change | |
| CD177 | 1.41 × 10−1 | 1.981 | 5.31 × 10−2 | 2.050 | 2.71 × 10−2 | 3.053 |
| FMO1 | 2.53 × 10−1 | 1.368 | 3.75 × 10−1 | 1.273 | 1.49 × 10−2 | 2.108 |
| GLUD1 | 3.61 × 10−2 | 1.644 | 6.07 × 10−3 | 1.975 | 3.32 × 10−4 | 2.746 |
| GSTM2 | 2.59 × 10−4 | 2.387 | 6.38 × 10−6 | 3.312 | 1.30 × 10−7 | 5.251 |
| HIST1H3A | 3.84 × 10−2 | 1.333 | 1.75 × 10−5 | 2.528 | 2.80 × 10−8 | 4.660 |
| HIST4H4 | 9.95 × 10−2 | 1.112 | 5.19 × 10−8 | 2.655 | 2.49 × 10−10 | 2.164 |
| RNU11 | 4.65 × 10−5 | 1.963 | 2.32 × 10−5 | 2.045 | 5.64 × 10−6 | 2.316 |
| RNU4-1 | 1.02 × 10−1 | 1.365 | 1.51 × 10−2 | 1.624 | 6.75 × 10−4 | 2.163 |
| SLC27A2 | 4.71 × 10−1 | 1.161 | 2.44 × 10−3 | 2.033 | 7.55 × 10−1 | 1.070 |
| Functions | P-Value | No of Genes |
|---|---|---|
| Cell death | 1.47 × 10−7 | 20 |
| Necrosis | 3.06 × 10−7 | 18 |
| Transcription of RNA | 4.12 × 10−4 | 17 |
| Binding of DNA | 5.11 × 10−4 | 17 |
| Transcription of RNA | 7.22 × 10−4 | 12 |
| Cellular assembly and organization | 1.23 × 10−6 | 9 |
| Transcription of DNA | 5.63 × 10−7 | 8 |
| Activation of DNA endogenous promotor | 2.14 × 10−6 | 5 |
| Cell-cycle progression | 2.27 × 10−3 | 4 |
| Gene expression | 1.35 × 10−4 | 2 |
| Symbol | 2 Days vs. Ctr | 4 Days vs. Ctr | 7 Days vs. Ctr | |||
|---|---|---|---|---|---|---|
| P-Value | Fold Change | P-Value | Fold Change | P-Value | Fold Change | |
| ANKRD50 | 2.82 × 10−2 | −1.323 | 2.32 × 10−6 | −2.191 | 2.35 × 10−4 | −1.749 |
| ANKRD36B | 4.65 × 10−3 | −2.292 | 1.85 × 10−3 | −2.546 | 9.24 × 10−3 | −1.979 |
| C9orf3 | 7.91 × 10−4 | −3.622 | 5.04 × 10−4 | −3.860 | 6.56 × 10−2 | −1.938 |
| CCDC88C | 1.82 × 10−1 | −1.363 | 1.51 × 10−1 | −1.398 | 6.70 × 10−3 | −2.041 |
| CYP24A1 | 9.28 × 10−1 | −1.027 | 8.44 × 10−1 | −1.060 | 3.32 × 10−2 | −2.011 |
| DGKH | 3.48 × 10−5 | −1.619 | 8.80 × 10−8 | −2.114 | 2.51 × 10−5 | −1.681 |
| DHFR | 3.77 × 10−4 | −1.779 | 6.90 × 10−4 | −1.717 | 3.10 × 10−5 | −2.139 |
| FAP | 2.07 × 10−1 | −1.562 | 4.72 × 10−1 | −1.284 | 3.01 × 10−2 | −2.314 |
| GTF2B | 2.33 × 10−2 | −1.580 | 6.39 × 10−4 | −2.131 | 1.37 × 10−3 | −2.072 |
| HDCA1 | 8.97 × 10−1 | −1.015 | 2.43 × 10−1 | −1.147 | 1.11 × 10−2 | −3.399 |
| HAS2 | 6.50 × 10−2 | −1.375 | 7.49 × 10−4 | −1.920 | 8.18 × 10−5 | −2.342 |
| LIF | 4.97 × 10−2 | −1.906 | 2.85 × 10−1 | −1.403 | 1.23 × 10−2 | −2.440 |
| LRRN1 | 2.41 × 10−1 | −1.422 | 1.78 × 10−1 | −1.501 | 1.71 × 10−2 | −2.217 |
| mir-21 | 5.77 × 10−4 | −2.677 | 6.96 × 10−5 | −3.347 | 7.02 × 10−3 | −2.131 |
| MPLKIP | 2.86 × 10−4 | −1.829 | 1.59 × 10−4 | −1.983 | 4.89 × 10−6 | −2.618 |
| NPIPL3 | 3.40 × 10−3 | −2.249 | 2.27 × 10−3 | −2.348 | 5.88 × 10−2 | −1.398 |
| NRG1 | 1.57 × 10−3 | −1.666 | 4.65 × 10−5 | −2.067 | 1.12 × 10−4 | −2.022 |
| PLA2G7 | 6.14 × 10−1 | −1.062 | 3.73 × 10−2 | −1.299 | 1.61 × 10−5 | −2.018 |
| PSD3 | 1.43 × 10−1 | −1.141 | 1.78 × 10−5 | −2.162 | 9.70 × 10−5 | −2.012 |
| RNF152 | 4.83 × 10−1 | −1.118 | 1.16 × 10−4 | −2.119 | 6.23 × 10−2 | −1.381 |
| SESN3 | 6.85 × 10−2 | −1.206 | 5.50 × 10−7 | −2.044 | 6.02 × 10−5 | −1.684 |
| SLC7A11 | 1.55 × 10−1 | −1.279 | 3.03 × 10−4 | −2.078 | 1.02 × 10−3 | −1.963 |
| SMAD2 | 4.59 × 10−3 | −1.526 | 9.51 × 10−6 | −2.195 | 2.76 × 10−4 | −1.848 |
| SMG1 | 1.43 × 10−4 | −2.030 | 2.45 × 10−5 | −2.503 | 4.67 × 10−4 | −1.395 |
| TAF1D | 7.99 × 10−3 | −1.961 | 2.85 × 10−3 | −2.177 | 2.71 × 10−2 | −1.770 |
| TRA2A | 1.38 × 10−3 | −2.456 | 3.45 × 10−3 | −2.229 | 6.73 × 10−4 | −1.357 |
| Functions | P-Value | No of Genes |
|---|---|---|
| Cellular assembly and organization | 4.28 × 10−5 | 19 |
| Differentiation of cells | 1.51 × 10−2 | 19 |
| DNA repair | 6.17 × 10−3 | 16 |
| Cellular function and maintenance | 1.31 × 10−2 | 13 |
| Transactivation of RNA | 4.60 × 10−4 | 10 |
| Binding of DNA | 1.86 × 10−3 | 9 |
| Activation of DNA endogenous promotor | 3.45 × 10−3 | 8 |
| G1/S phase transition | 9.14 × 10−3 | 6 |
| Cell-cycle progression | 3.23 × 10−3 | 2 |
| Transcription of DNA | 1.12 × 10−2 | 3 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paaske Utheim, T.; Salvanos, P.; Aass Utheim, Ø.; Ræder, S.; Pasovic, L.; Olstad, O.K.; Fideliz de la Paz, M.; Sehic, A. Transcriptome Analysis of Cultured Limbal Epithelial Cells on an Intact Amniotic Membrane following Hypothermic Storage in Optisol-GS. J. Funct. Biomater. 2016, 7, 4. https://doi.org/10.3390/jfb7010004
Paaske Utheim T, Salvanos P, Aass Utheim Ø, Ræder S, Pasovic L, Olstad OK, Fideliz de la Paz M, Sehic A. Transcriptome Analysis of Cultured Limbal Epithelial Cells on an Intact Amniotic Membrane following Hypothermic Storage in Optisol-GS. Journal of Functional Biomaterials. 2016; 7(1):4. https://doi.org/10.3390/jfb7010004
Chicago/Turabian StylePaaske Utheim, Tor, Panagiotis Salvanos, Øygunn Aass Utheim, Sten Ræder, Lara Pasovic, Ole Kristoffer Olstad, Maria Fideliz de la Paz, and Amer Sehic. 2016. "Transcriptome Analysis of Cultured Limbal Epithelial Cells on an Intact Amniotic Membrane following Hypothermic Storage in Optisol-GS" Journal of Functional Biomaterials 7, no. 1: 4. https://doi.org/10.3390/jfb7010004