How to Address the Adjuvant Effects of Nanoparticles on the Immune System

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
2.2. Characterization of aSNP
2.3. Endotoxin Detection
2.4. Generation of Primary Cultures of Human Dendritic Cells
2.5. Phenotypic Analysis
2.6. Co-Culture of MoDCs and CD4+ T-Cells
3. Results and Discussion
3.1. Characterization of Amorphous Silica Nanoparticles
3.2. Assessing the Effects of Amorphous Silica Nanoparticules on the Expression of Major Histocompatibility Complex Class II and co-stimulatory Molecules in Human Dendritic Cells
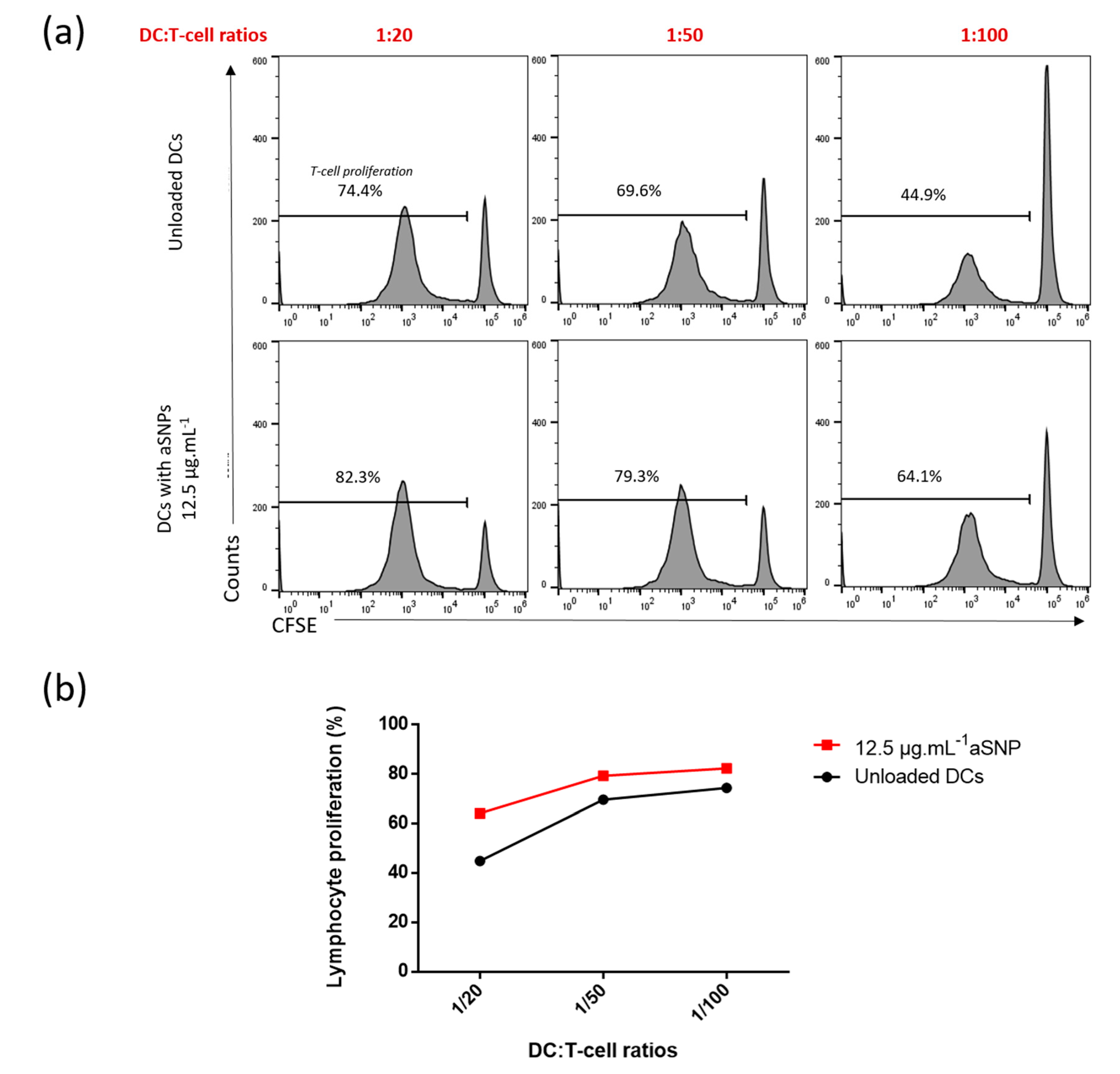
3.3. Assessing the Effect of Dendritic Cells Treated with Amorphous Silica Nanoparticules on Allogeneic T-Cell Proliferation
3.3.1. Influence of DC:T-Cell Ratios on T-Cell Proliferation
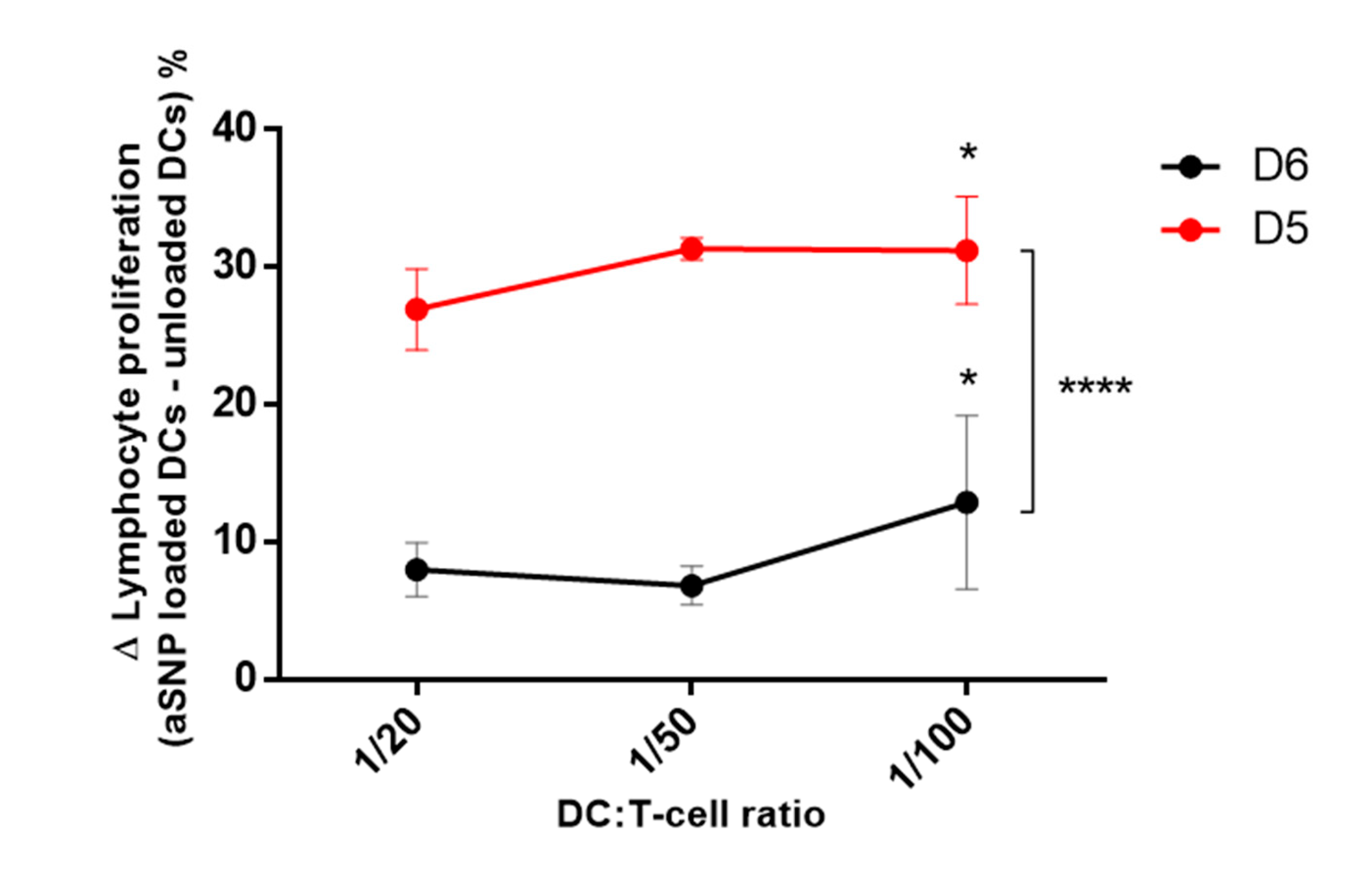
3.3.2. Influence of the co-Culture Duration on T-Cell Proliferation and Assessment of Nanoparticles Effects
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shannahan, J.H.; Brown, J.M. Engineered nanomaterial exposure and the risk of allergic disease. Curr. Opin. Allergy Clin. Immunol. 2014, 14, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Oostingh, G.J.; Casals, E.; Italiani, P.; Colognato, R.; Stritzinger, R.; Ponti, J.; Pfaller, T.; Kohl, Y.; Ooms, D.; Favilli, F.; et al. Problems and challenges in the development and validation of human cell-based assays to determine nanoparticle-induced immunomodulatory effects. Part Fibre Toxicol. 2011, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boraschi, D.; Italiani, P.; Palomba, R.; Decuzzi, P.; Duschl, A.; Fadeel, B.; Moghimi, S.M. Nanoparticles and innate immunity: New perspectives on host defence. Semin. Immunol. 2017, 34, 33–51. [Google Scholar] [CrossRef] [PubMed]
- Drasler, B.; Sayre, P.; Steinhauser, K.G.; Petri-Fink, A.; Rothen-Rutishauser, B. In vitro approaches to assess the hazard of nanomaterials. NanoImpact 2017, 8, 99–116. [Google Scholar] [CrossRef]
- Najafi-Hajivar, S.; Zakeri-Milani, P.; Mohammadi, H.; Niazi, M.; Soleymani-Goloujeh, M.; Baradaran, B.; Valizadeh, H. Overview on experimental models of interactions between nanoparticles and the immune system. Biomed. Pharmacother. 2016, 83, 1365–1378. [Google Scholar] [CrossRef]
- Winter, M.; Beer, H.D.; Hornung, V.; Kramer, U.; Schins, R.P.; Forster, I. Activation of the inflammasome by amorphous silica and TiO2 nanoparticles in murine dendritic cells. Nanotoxicology 2011, 5, 326–340. [Google Scholar] [CrossRef]
- Deville, S.; Bare, B.; Piella, J.; Tirez, K.; Hoet, P.; Monopoli, M.P.; Dawson, K.A.; Puntes, V.F.; Nelissen, I. Interaction of gold nanoparticles and nickel(II) sulfate affects dendritic cell maturation. Nanotoxicology 2016, 10, 1395–1403. [Google Scholar] [CrossRef]
- Pfaller, T.; Colognato, R.; Nelissen, I.; Favilli, F.; Casals, E.; Ooms, D.; Leppens, H.; Ponti, J.; Stritzinger, R.; Puntes, V.; et al. The suitability of different cellular in vitro immunotoxicity and genotoxicity methods for the analysis of nanoparticle-induced events. Nanotoxicology 2010, 4, 52–72. [Google Scholar] [CrossRef]
- Lambrecht, B.N. Dendritic cells and the regulation of the allergic immune response. Allergy 2005, 60, 271–282. [Google Scholar] [CrossRef]
- Oberdorster, G.; Oberdorster, E.; Oberdorster, J. Nanotoxicology: An emerging discipline evolving from studies of ultrafine particles. Environ. Health Perspect. 2005, 113, 823–839. [Google Scholar] [CrossRef]
- Lutz, M.B.; Schuler, G. Immature, semi-mature and fully mature dendritic cells: Which signals induce tolerance or immunity? Trends Immunol. 2002, 23, 445–449. [Google Scholar] [CrossRef]
- Gallo, P.M.; Gallucci, S. The dendritic cell response to classic, emerging, and homeostatic danger signals. Implications for autoimmunity. Front. Immunol. 2013, 4, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matzinger, P. Tolerance, danger, and the extended family. Annu. Rev. Immunol. 1994, 12, 991–1045. [Google Scholar] [CrossRef] [PubMed]
- de Haar, C.; Kool, M.; Hassing, I.; Bol, M.; Lambrecht, B.N.; Pieters, R. Lung dendritic cells are stimulated by ultrafine particles and play a key role in particle adjuvant activity. J. Allergy Clin. Immunol. 2008, 121, 1246–1254. [Google Scholar] [CrossRef] [PubMed]
- Fadeel, B. Clear and present danger? Engineered nanoparticles and the immune system. Swiss Med. Wkly. 2012, 142, w13609. [Google Scholar] [CrossRef]
- Pallardy, M.J.; Turbica, I.; Biola-Vidamment, A. Why the Immune System Should Be Concerned by Nanomaterials? Front. Immunol. 2017, 8, 544. [Google Scholar] [CrossRef]
- Vita, A.A.; Royse, E.A.; Pullen, N.A. Nanoparticles and danger signals: Oral delivery vehicles as potential disruptors of intestinal barrier homeostasis. J. Leukoc. Biol. 2019, 106, 95–103. [Google Scholar] [CrossRef]
- van Helden, S.F.; van Leeuwen, F.N.; Figdor, C.G. Human and murine model cell lines for dendritic cell biology evaluated. Immunol. Lett. 2008, 117, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Koike, E.; Takano, H.; Inoue, K.; Yanagisawa, R.; Kobayashi, T. Carbon black nanoparticles promote the maturation and function of mouse bone marrow-derived dendritic cells. Chemosphere 2008, 73, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Zhu, Y.; Zhang, M.; Xiao, Y.; Du, X.; Liu, H.; Wang, S. The induction of maturation on dendritic cells by TiO2 and Fe3O4@TiO2 nanoparticles via NF-kappaB signaling pathway. Mater. Sci. Eng. C Mater Biol. Appl. 2014, 39, 305–314. [Google Scholar] [CrossRef]
- Vallhov, H.; Gabrielsson, S.; Stromme, M.; Scheynius, A.; Garcia-Bennett, A.E. Mesoporous silica particles induce size dependent effects on human dendritic cells. Nano Lett. 2007, 7, 3576–3582. [Google Scholar] [CrossRef] [PubMed]
- Barillet, S.; Fattal, E.; Mura, S.; Tsapis, N.; Pallardy, M.; Hillaireau, H.; Kerdine-Romer, S. Immunotoxicity of poly (lactic-co-glycolic acid) nanoparticles: Influence of surface properties on dendritic cell activation. Nanotoxicology 2019, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hargadon, K.M. Murine and Human Model Systems for the Study of Dendritic Cell Immunobiology. Int. Rev. Immunol. 2016, 35, 85–115. [Google Scholar] [CrossRef] [PubMed]
- Perkins, D.J.; Vogel, S.N. Inflammation: Species-specific TLR signalling—Insight into human disease. Nat. Rev. 2016, 12, 198–200. [Google Scholar] [CrossRef] [Green Version]
- Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Dusemund, B.; Filipič, M.; Frutos, M.J.; Galtier, P.; Gott, D.; Gundert-Remy, U.; et al. Re-evaluation of silicon dioxide (E 551) as a food additive. EFSA J. 2018, 16. [Google Scholar] [CrossRef] [Green Version]
- Brandenberger, C.; Rowley, N.L.; Jackson-Humbles, D.N.; Zhang, Q.; Bramble, L.A.; Lewandowski, R.P.; Wagner, J.G.; Chen, W.; Kaplan, B.L.; Kaminski, N.E.; et al. Engineered silica nanoparticles act as adjuvants to enhance allergic airway disease in mice. Part Fibre Toxicol. 2013, 10, 26. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Park, Y.H.; Park, H.J.; Lee, K.; Um, K.; Park, J.W.; Lee, J.H. Toxic and adjuvant effects of silica nanoparticles on ovalbumin-induced allergic airway inflammation in mice. Respir. Res. 2016, 17, 60. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.; Lim, J.S. Induction of functional changes of dendritic cells by silica nanoparticles. Immune Netw. 2012, 12, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Winkler, H.C.; Kornprobst, J.; Wick, P.; von Moos, L.M.; Trantakis, I.; Schraner, E.M.; Bathke, B.; Hochrein, H.; Suter, M.; Naegeli, H. MyD88-dependent pro-interleukin-1beta induction in dendritic cells exposed to food-grade synthetic amorphous silica. Part Fibre Toxicol. 2017, 14, 21. [Google Scholar] [CrossRef] [Green Version]
- Schanen, B.C.; Das, S.; Reilly, C.M.; Warren, W.L.; Self, W.T.; Seal, S.; Drake, D.R., 3rd. Immunomodulation and T helper TH1/TH2 response polarization by CeO2 and TiO2 nanoparticles. PLoS ONE 2013, 3, e62816. [Google Scholar] [CrossRef]
- Li, Y.; Fujita, M.; Boraschi, D. Endotoxin Contamination in Nanomaterials Leads to the Misinterpretation of Immunosafety Results. Front. Immunol. 2017, 8, 472. [Google Scholar] [CrossRef]
- Tan, J.K.; O’Neill, H.C. Maturation requirements for dendritic cells in T cell stimulation leading to tolerance versus immunity. J. Leukoc. Biol. 2005, 78, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Kim, J. Properties of immature and mature dendritic cells: Phenotype, morphology, phagocytosis, and migration. Rsc Adv. 2019, 9, 11230–11238. [Google Scholar] [CrossRef] [Green Version]
- Aerts-Toegaert, C.; Heirman, C.; Tuyaerts, S.; Corthals, J.; Aerts, J.L.; Bonehill, A.; Thielemans, K.; Breckpot, K. CD83 expression on dendritic cells and T cells: correlation with effective immune responses. Eur. J. Immunol. 2007, 37, 686–695. [Google Scholar] [CrossRef] [PubMed]
- Hopken, U.E.; Lehmann, I.; Droese, J.; Lipp, M.; Schuler, T.; Rehm, A. The ratio between dendritic cells and T cells determines the outcome of their encounter: proliferation versus deletion. Eur. J. Immunol. 2005, 35, 2851–2863. [Google Scholar] [CrossRef] [PubMed]
- Gallais, Y.; Szely, N.; Legrand, F.X.; Leroy, A.; Pallardy, M.; Turbica, I. Effect of growth hormone and IgG aggregates on dendritic cells activation and T-cells polarization. Immunol Cell Biol 2017, 95, 306–315. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Specific Surface Area 1 | Nominal Primary Particle Diameter 1 | Primary Particle Diameter 2 | DLS Distribution 3 | ζ-Potential Value 3 |
|---|---|---|---|---|---|
| S5505, Sigma-Aldrich (batch SLBR6988V) | 196 m2.g−1 | 14 nm | 14.4 ± 4.3 nm | 201 ± 22 nm 9.0 ± 0.3 µm | −26.5 ± 2.1 mV |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feray, A.; Szely, N.; Guillet, E.; Hullo, M.; Legrand, F.-X.; Brun, E.; Pallardy, M.; Biola-Vidamment, A. How to Address the Adjuvant Effects of Nanoparticles on the Immune System. Nanomaterials 2020, 10, 425. https://doi.org/10.3390/nano10030425
Feray A, Szely N, Guillet E, Hullo M, Legrand F-X, Brun E, Pallardy M, Biola-Vidamment A. How to Address the Adjuvant Effects of Nanoparticles on the Immune System. Nanomaterials. 2020; 10(3):425. https://doi.org/10.3390/nano10030425
Chicago/Turabian StyleFeray, Alexia, Natacha Szely, Eléonore Guillet, Marie Hullo, François-Xavier Legrand, Emilie Brun, Marc Pallardy, and Armelle Biola-Vidamment. 2020. "How to Address the Adjuvant Effects of Nanoparticles on the Immune System" Nanomaterials 10, no. 3: 425. https://doi.org/10.3390/nano10030425