
In Vitro Activities of Oxazolidinone Antibiotics Alone and in Combination with C-TEMPO against Methicillin-Resistant Staphylococcus aureus Biofilms
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. The Oxazolidinones Have Inherent Biofilm Activity against S. aureus Biofilms
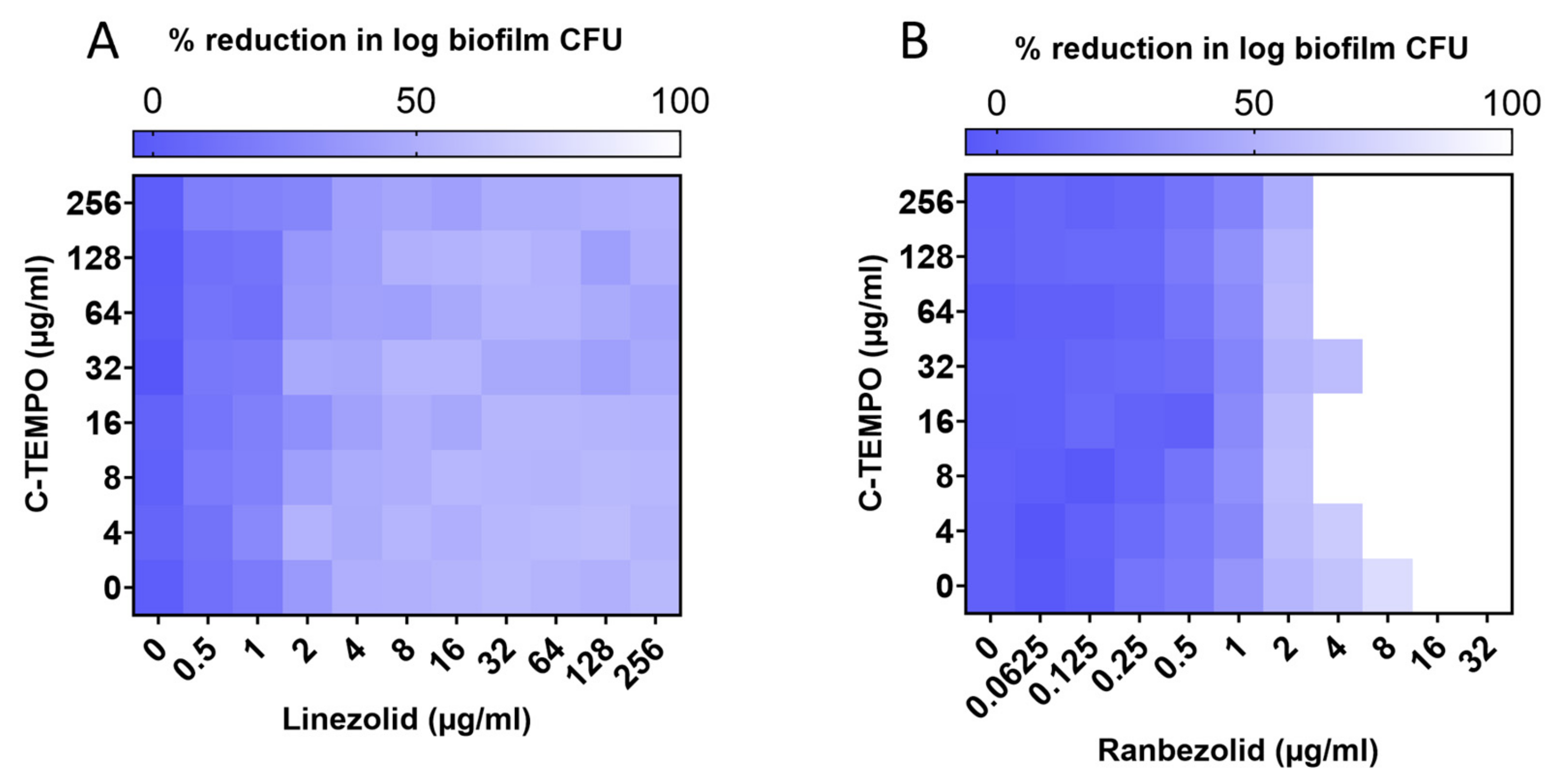
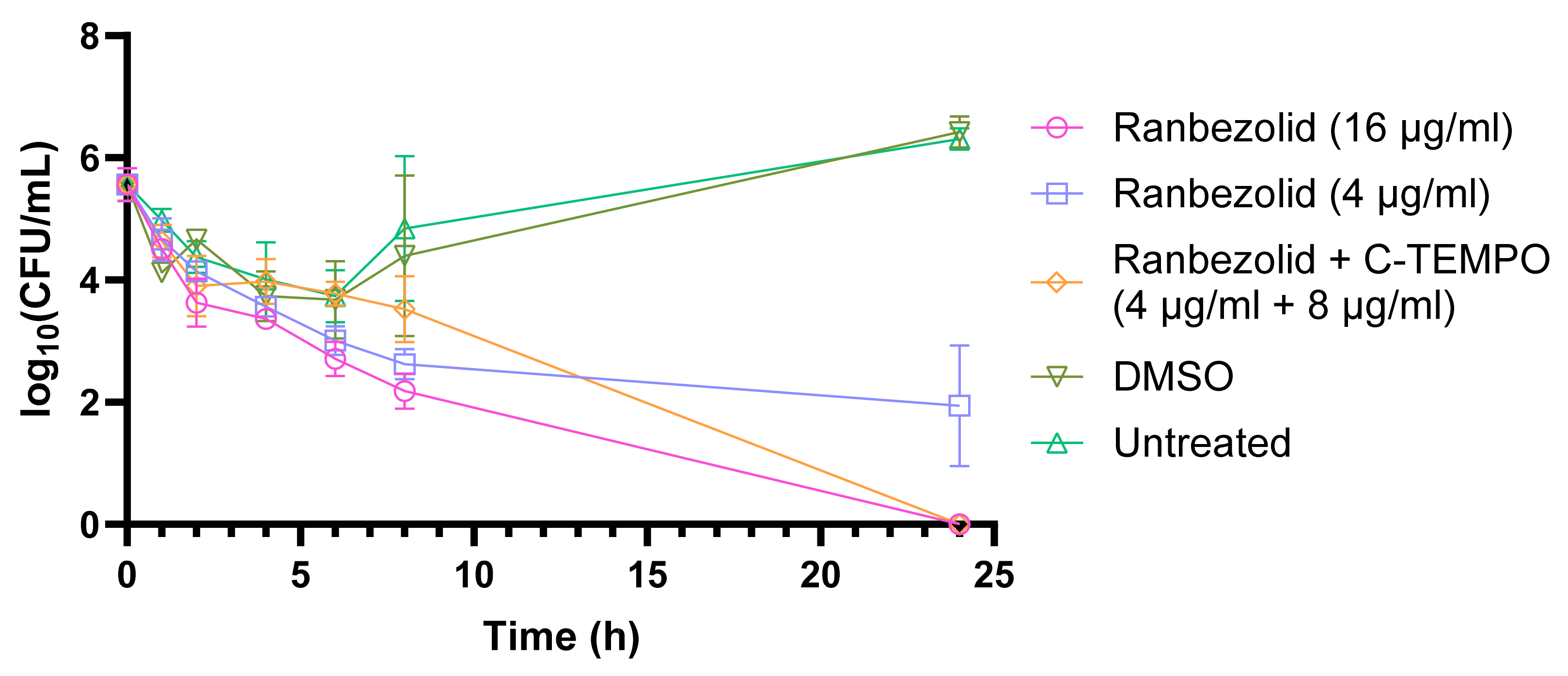
2.2. C-TEMPO Coadministration Improves the Susceptibility of S. aureus Biofilms to Ranbezolid
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Nitroxide and Antibiotics
4.3. Nitroxide and Antibiotic MIC Assay
4.4. Biofilm Culture Using the Minimum Biofilm Eradication Concentration Device
4.5. Minimum Biofilm Eradication Concentration (MBEC) Assays
4.6. Checkerboard MBEC Assay
4.7. Biofilm Time Kill Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MacKenzie, F.M.; Bruce, J.; Struelens, M.J.; Goossens, H.; Mollison, J.; Gould, I.M. Antimicrobial drug use and infection control practices associated with the prevalence of methicillin-resistant Staphylococcus aureus in European hospitals. Clin. Microbiol. Infect. 2007, 13, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, K.T.; Calderon, L.E.; Saman, D.M.; Abusalem, S.K. The use of surveillance and preventative measures for methicillin-resistant Staphylococcus aureus infections in surgical patients. Antimicrob. Resist. Infect. Control 2014, 3, 18. [Google Scholar] [CrossRef] [PubMed]
- Hassoun, A.; Linden, P.K.; Friedman, B. Incidence, prevalence, and management of MRSA bacteremia across patient populations—A review of recent developments in MRSA management and treatment. Crit. Care 2017, 21, 211. [Google Scholar] [CrossRef] [PubMed]
- Appelbaum, P.C. MRSA—The tip of the iceberg. Clin. Microbiol. Infect. 2006, 12, 3–10. [Google Scholar] [CrossRef]
- Noskin, G.A.; Rubin, R.J.; Schentag, J.J.; Kluytmans, J.; Hedblom, E.C.; Smulders, M.; Lapetina, E.; Gemmen, E. The Burden of Staphylococcus aureus Infections on Hospitals in the United States: An Analysis of the 2000 and 2001 Nationwide Inpatient Sample Database. Arch. Intern. Med. 2005, 165, 1756–1761. [Google Scholar] [CrossRef] [PubMed]
- Shoaib, M.; Aqib, A.I.; Muzammil, I.; Majeed, N.; Bhutta, Z.A.; Kulyar, M.F.; Fatima, M.; Zaheer, C.F.; Muneer, A.; Murtaza, M.; et al. MRSA compendium of epidemiology, transmission, pathophysiology, treatment, and prevention within one health framework. Front. Microbiol. 2022, 13, 1067284. [Google Scholar] [CrossRef] [PubMed]
- Pichereau, S.; Rose, W.E. Invasive community-associated MRSA infections: Epidemiology and antimicrobial management. Expert Opin. Pharmacother. 2010, 11, 3009–3025. [Google Scholar] [CrossRef] [PubMed]
- Cascioferro, S.; Carbone, D.; Parrino, B.; Pecoraro, C.; Giovannetti, E.; Cirrincione, G.; Diana, P. Therapeutic Strategies to Counteract Antibiotic Resistance in MRSA Biofilm-Associated Infections. ChemMedChem 2021, 16, 65–80. [Google Scholar] [CrossRef]
- Stewart, P.S.; William Costerton, J. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Stoodley, P. Evolving concepts in biofilm infections. Cell. Microbiol. 2009, 11, 1034–1043. [Google Scholar] [CrossRef]
- Silva, V.; Almeida, L.; Gaio, V.; Cerca, N.; Manageiro, V.; Caniça, M.; Capelo, J.L.; Igrejas, G.; Poeta, P. Biofilm Formation of Multidrug-Resistant MRSA Strains Isolated from Different Types of Human Infections. Pathogens 2021, 10, 970. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, P. Extracellular polymeric substances, a key element in understanding biofilm phenotype. AIMS Microbiol. 2018, 4, 274–288. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente-Núñez, C.; Reffuveille, F.; Fairfull-Smith, K.E.; Hancock, R.E.W. Effect of Nitroxides on Swarming Motility and Biofilm Formation, Multicellular Behaviors in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2013, 57, 4877–4881. [Google Scholar] [CrossRef] [PubMed]
- Fleming, D.; Rumbaugh, K. The Consequences of Biofilm Dispersal on the Host. Sci. Rep. 2018, 8, 10738. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation. Prioritization of Pathogens to Guide Discovery, Research and Development of New Antibiotics for Drug Resistant Bacterial Infections, Including Tuberculosis; World Health Organisation: Geneva, Switzerland, 2017; p. 88. [Google Scholar]
- Rybak, M.J.; Hershberger, E.; Moldovan, T.; Grucz, R.G. In Vitro Activities of Daptomycin, Vancomycin, Linezolid, and Quinupristin-Dalfopristin against Staphylococci and Enterococci, Including Vancomycin- Intermediate and -Resistant Strains. Antimicrob. Agents Chemother. 2000, 44, 1062–1066. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.M.; Jarvis, B. Linezolid. Drugs 2001, 61, 525–551. [Google Scholar] [CrossRef] [PubMed]
- Ford, W.C.; Zurenko, E.G.; Barbachyn, R.M. The Discovery of Linezolid, the First Oxazolidinone Antibacterial Agent. Curr. Drug Targets Infect. Disord. 2001, 1, 181–199. [Google Scholar] [CrossRef] [PubMed]
- Barman, T.K.; Kumar, M.; Mathur, T.; Chaira, T.; Ramkumar, G.; Kalia, V.; Rao, M.; Pandya, M.; Yadav, A.S.; Das, B.; et al. In Vitro and In Vivo Activities of a Bi-Aryl Oxazolidinone, RBx 11760, against Gram-Positive Bacteria. Antimicrob. Agents Chemother. 2016, 60, 7134–7145. [Google Scholar] [CrossRef]
- Douros, A.; Grabowski, K.; Stahlmann, R. Drug–drug interactions and safety of linezolid, tedizolid, and other oxazolidinones. Expert Opin. Drug Metab. Toxicol. 2015, 11, 1849–1859. [Google Scholar] [CrossRef]
- Furin, J.J.; Du Bois, J.; van Brakel, E.; Chheng, P.; Venter, A.; Peloquin, C.A.; Alsultan, A.; Thiel, B.A.; Debanne, S.M.; Boom, W.H.; et al. Early Bactericidal Activity of AZD5847 in Patients with Pulmonary Tuberculosis. Antimicrob. Agents Chemother. 2016, 60, 6591–6599. [Google Scholar] [CrossRef]
- LegoChem Biosciences. A Phase II Clinical Study of LCB01-0371 to Evaluate the EBA, Safety and PK (Identifier: NCT02836483). 2016. Available online: https://clinicaltrials.gov/ct2/show/NCT02836483 (accessed on 18 June 2023).
- Melinta Therapeutics. Safety and Efficacy Study of Oxazolidinones to Treat Uncomplicated Skin Infections (Identifier: NCT00646958). 2008. Available online: https://clinicaltrials.gov/ct2/show/NCT00646958 (accessed on 18 June 2023).
- Melinta Therapeutics. Safety and Efficacy Study of Oxazolidinone to Treat Pneumonia (Identifier: NCT00640926). 2008. Available online: https://clinicaltrials.gov/ct2/show/NCT00640926 (accessed on 18 June 2023).
- MicuRx. Contezolid Acefosamil Versus Linezolid for the Treatment of Acute Bacterial Skin and Skin Structure Infection Identifier: (NCT03747497). 2018. Available online: https://clinicaltrials.gov/ct2/show/NCT03747497 (accessed on 18 June 2023).
- Actelion. Efficacy and Safety of Cadazolid Versus Vancomycin in Subjects with Clostridium Difficile-Associated Diarrhea (Identifier: NCT01983683). 2013. Available online: https://clinicaltrials.gov/ct2/show/NCT01983683 (accessed on 18 June 2023).
- LegoChem Biosciences. PanACEA DElpazolid Dose-Finding and COmbination DEvelopment (DECODE) (Identifier: NCT04550832). 2020. Available online: https://clinicaltrials.gov/ct2/show/NCT04550832 (accessed on 18 June 2023).
- University of Munich; Radboud University Medical Center; University of California; University College London. PanACEA–STEP2C-01 (NCT05807399). 2023. Available online: https://clinicaltrials.gov/ct2/show/NCT05807399 (accessed on 18 June 2023).
- Yuan, S.; Shen, D.-D.; Bai, Y.-R.; Zhang, M.; Zhou, T.; Sun, C.; Zhou, L.; Wang, S.-Q.; Liu, H.-M. Oxazolidinone: A promising scaffold for the development of antibacterial drugs. Eur. J. Med. Chem. 2023, 250, 115239. [Google Scholar] [CrossRef]
- LegoChem Biosciences. A Phase 2a Study, Effect of Vancomycin with vs. without Delpazolid (LCB01-0371) in Patients with MRSA Bacteremia (NCT05225558). 2022. Available online: https://clinicaltrials.gov/ct2/show/NCT05225558 (accessed on 18 June 2023).
- Andrews, J. To be or not to be exclusive: The sutezolid story. Lancet Glob. Health 2016, 4, e89–e90. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Kim, Y.-H.; Lee, S.H.; Kim, Y.H.; Kim, J.-W.; Kang, J.Y.; Kim, S.K.; Kim, S.J.; Kang, Y.-S.; Kim, T.-H.; et al. Early Bactericidal Activity of Delpazolid (LCB01-0371) in Patients with Pulmonary Tuberculosis. Antimicrob. Agents Chemother. 2022, 66, e01684-21. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.-W.; Jung, S.-J.; Lee, H.-H.; Kim, Y.-Z.; Park, T.-K.; Cho, Y.-L.; Chae, S.-E.; Baek, S.-Y.; Woo, S.-H.; Lee, H.-S.; et al. In Vitro and In Vivo Activities of LCB01-0371, a New Oxazolidinone. Antimicrob. Agents Chemother. 2010, 54, 5359–5362. [Google Scholar] [CrossRef] [PubMed]
- Shaw, K.J.; Barbachyn, M.R. The oxazolidinones: Past, present, and future. Ann. N. Y. Acad. Sci. 2011, 1241, 48–70. [Google Scholar] [CrossRef] [PubMed]
- Vastra Gotaland Region. Susceptibility Testing of Biofilm to Guide Treatment of Periprosthetic Joint Infections (Identifier: NCT04488458). 2021. Available online: https://clinicaltrials.gov/study/NCT04488458 (accessed on 18 June 2023).
- Gordon, R.J.; Lowy, F.D. Pathogenesis of Methicillin-Resistant Staphylococcus aureus Infection. Clin. Infect. Dis. 2008, 46, S350–S359. [Google Scholar] [CrossRef] [PubMed]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef]
- Luppens, S.B.I.; Reij, M.W.; van der Heijden, R.W.L.; Rombouts, F.M.; Abee, T. Development of a Standard Test To Assess the Resistance of Staphylococcus aureus Biofilm Cells to Disinfectants. Appl. Environ. Microbiol. 2002, 68, 4194–4200. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M. Biofilms and device-associated infections. Emerg. Infect. Dis. 2001, 7, 277–281. [Google Scholar] [CrossRef]
- Assefa, M.; Amare, A. Biofilm-Associated Multi-Drug Resistance in Hospital-Acquired Infections: A Review. Infect. Drug. Resist. 2022, 15, 5061–5068. [Google Scholar] [CrossRef]
- Loffler, C.A.; MacDougall, C. Update on prevalence and treatment of methicillin-resistant Staphylococcus aureus infections. Expert Rev. Anti Infect. Ther. 2007, 5, 961–981. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, M.; Frees, D.; Ingmer, H. Antibiotic Resistance and the MRSA Problem. Microbiol. Spectr. 2019, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Pillai, S.K.; Sakoulas, G.; Wennersten, C.; Eliopoulos, G.M.; Moellering, R.C., Jr.; Ferraro, M.J.; Gold, H.S. Linezolid Resistance in Staphylococcus aureus: Characterization and Stability of Resistant Phenotype. J. Infect. Dis. 2002, 186, 1603–1607. [Google Scholar] [CrossRef]
- Farrell, D.J.; Mendes, R.E.; Ross, J.E.; Jones, R.N. Linezolid surveillance program results for 2008 (LEADER Program for 2008). Diagn. Microbiol. Infect. Dis. 2009, 65, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Markwart, R.; Willrich, N.; Eckmanns, T.; Werner, G.; Ayobami, O. Low Proportion of Linezolid and Daptomycin Resistance Among Bloodborne Vancomycin-Resistant Enterococcus faecium and Methicillin-Resistant Staphylococcus aureus Infections in Europe. Front. Microbiol 2021, 12, 664199. [Google Scholar] [CrossRef] [PubMed]
- Katayama, Y.; Ito, T.; Hiramatsu, K. A new class of genetic element, staphylococcus cassette chromosome mec, encodes methicillin resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 2000, 44, 1549–1555. [Google Scholar] [CrossRef]
- Wali, M.; Shah, M.S.; Rehman, T.U.; Wali, H.; Hussain, M.; Zaman, L.; Khan, F.U.; Mangi, A.H. Detection of linezolid resistance cfr gene among MRSA isolates. J. Infect. Public Health 2022, 15, 1142–1146. [Google Scholar] [CrossRef] [PubMed]
- AbdAlhafiz, A.I.; Elleboudy, N.S.; Aboshanab, K.M.; Aboulwafa, M.M.; Hassouna, N.A. Phenotypic and genotypic characterization of linezolid resistance and the effect of antibiotic combinations on methicillin-resistant Staphylococcus aureus clinical isolates. Ann. Clin. Microbiol. Antimicrob. 2023, 22, 23. [Google Scholar] [CrossRef]
- Abad, L.; Tafani, V.; Tasse, J.; Josse, J.; Chidiac, C.; Lustig, S.; Ferry, T.; Diot, A.; Laurent, F.; Valour, F. Evaluation of the ability of linezolid and tedizolid to eradicate intraosteoblastic and biofilm-embedded Staphylococcus aureus in the bone and joint infection setting. J. Antimicrob. Chemother. 2019, 74, 625–632. [Google Scholar] [CrossRef]
- Wu, S.; Yang, T.; Luo, Y.; Li, X.; Zhang, X.; Tang, J.; Ma, X.; Wang, Z. Efficacy of the novel oxazolidinone compound FYL-67 for preventing biofilm formation by Staphylococcus aureus. J. Antimicrob. Chemother. 2014, 69, 3011–3019. [Google Scholar] [CrossRef]
- Svensson Malchau, K.; Tillander, J.; Zaborowska, M.; Hoffman, M.; Lasa, I.; Thomsen, P.; Malchau, H.; Rolfson, O.; Trobos, M. Biofilm properties in relation to treatment outcome in patients with first-time periprosthetic hip or knee joint infection. J. Orthop. Transl. 2021, 30, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Ndukwe, A.R.N.; Wiedbrauk, S.; Boase, N.R.B.; Fairfull-Smith, K.E. Strategies to Improve the Potency of Oxazolidinones towards Bacterial Biofilms. Chem. Asian J. 2022, 17, e202200201. [Google Scholar] [CrossRef] [PubMed]
- Martínez, S.R.; Rocca, D.M.; Aiassa, V.; Becerra, M.C. Linezolid as an eradication agent against assembled methicillin-resistant Staphylococcus aureus biofilms. RSC Adv. 2016, 6, 101023–101028. [Google Scholar] [CrossRef]
- Guo, P.; Buttaro, B.A.; Xue, H.Y.; Tran, N.T.; Wong, H.L. Lipid-polymer hybrid nanoparticles carrying linezolid improve treatment of methicillin-resistant Staphylococcus aureus (MRSA) harbored inside bone cells and biofilms. Eur. J. Pharm. Biopharm. 2020, 151, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Brackman, G.; Breyne, K.; De Rycke, R.; Vermote, A.; Van Nieuwerburgh, F.; Meyer, E.; Van Calenbergh, S.; Coenye, T. The Quorum Sensing Inhibitor Hamamelitannin Increases Antibiotic Susceptibility of Staphylococcus aureus Biofilms by Affecting Peptidoglycan Biosynthesis and eDNA Release. Sci. Rep. 2016, 6, 20321. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, B.K.; Soni, R.; Patel, J.Z.; Solanki, M.; Valani, D.; Gupta, S.; Mishra, B.; Takale, V.; Pandya, P.; Jain, M.R.; et al. Synthesis and in vitro antibacterial activity of novel methylamino piperidinyl oxazolidinones. Bioorganic Med. Chem. Lett. 2007, 17, 5227–5232. [Google Scholar] [CrossRef] [PubMed]
- Roch, M.; Varela, M.C.; Taglialegna, A.; Rosato, A.E. Tedizolid is a promising antimicrobial option for the treatment of Staphylococcus aureus infections in cystic fibrosis patients. J. Antimicrob. Chemother. 2019, 75, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, S.; Kosowska-Shick, K.; Appelbaum, P.C.; Verween, G.; Tulkens, P.M.; Bambeke, F.V. Cellular Pharmacodynamics of the Novel Biaryloxazolidinone Radezolid: Studies with Infected Phagocytic and Nonphagocytic cells, Using Staphylococcus aureus, Staphylococcus epidermidis, Listeria monocytogenes, and Legionella pneumophila. Antimicrob. Agents Chemother. 2010, 54, 2549–2559. [Google Scholar] [CrossRef]
- Ndukwe, A.R.N.; Hawas, S.; Qin, J.; Wiedbrauk, S.; Totsika, M.; Boase, N.R.B.; Fairfull-Smith, K.E. Simple and Efficient Synthesis of 3-Aryl-2-oxazolidinone Scaffolds Enabling Increased Potency toward Biofilms. Mol. Pharm. 2023, 20, 3484–3493. [Google Scholar] [CrossRef]
- Verderosa, A.D.; Hawas, S.; Harris, J.; Totsika, M.; Fairfull-Smith, K.E. Isothiazolone–Nitroxide Hybrids with Activity against Antibiotic-Resistant Staphylococcus aureus Biofilms. ACS Omega 2022, 7, 5300–5310. [Google Scholar] [CrossRef]
- Nasr, R.A.; AbuShady, H.M.; Hussein, H.S. Biofilm formation and presence of icaAD gene in clinical isolates of staphylococci. Egypt. J. Med. Hum. Genet. 2012, 13, 269–274. [Google Scholar] [CrossRef]
- Tuon, F.F.; Suss, P.H.; Telles, J.P.; Dantas, L.R.; Borges, N.H.; Ribeiro, V.S. Antimicrobial Treatment of Staphylococcus aureus Biofilms. Antibiotics 2023, 12, 87. [Google Scholar] [CrossRef] [PubMed]
- Yazdani, R.; Oshaghi, M.; Havaei, A.; Pishva, E.; Salehi, R.; Sadeghizadeh, M.; Forouhesh, H. Detection of icaAD gene and biofilm formation in Staphylococcus aureus isolates from wound infections. Iran. J. Public Health 2006, 35, 25–28. [Google Scholar]
- Mohammad, G.J. Expression of icaA, B, D, R and ebps biofilm-associated genes in methicillin-resistant-Staphylococcus aureus in exposure to curcumin. Gene Rep. 2022, 27, 101616. [Google Scholar] [CrossRef]
- Götz, F. Staphylococcus and biofilms. Mol. Microbiol. 2002, 43, 1367–1378. [Google Scholar] [CrossRef] [PubMed]
- Arciola, C.R.; Baldassarri, L.; Montanaro, L. Presence of icaA and icaD genes and slime production in a collection of staphylococcal strains from catheter-associated infections. J. Clin. Microbiol. 2001, 39, 2151–2156. [Google Scholar] [CrossRef]
- Singh, R.; Ray, P.; Das, A.; Sharma, M. Penetration of antibiotics through Staphylococcus aureus and Staphylococcus epidermidis biofilms. J. Antimicrob. Chemother. 2010, 65, 1955–1958. [Google Scholar] [CrossRef]
- Mathur, T.; Singhal, S.; Khan, S.; Upadhyay, D.; Fatma, T.; Rattan, A. Adverse effect of staphylococci slime on in vitro activity of glycopeptides. Jpn J. Infect. Dis. 2005, 58, 353. [Google Scholar]
- Liu, J.; Madec, J.Y.; Bousquet-Mélou, A.; Haenni, M.; Ferran, A.A. Destruction of Staphylococcus aureus biofilms by combining an antibiotic with subtilisin A or calcium gluconate. Sci. Rep. 2021, 11, 6225. [Google Scholar] [CrossRef]
- Mugunthan, S.; Wong, L.L.; Winnerdy, F.R.; Summers, S.; Bin Ismail, M.H.; Foo, Y.H.; Jaggi, T.K.; Meldrum, O.W.; Tiew, P.Y.; Chotirmall, S.H.; et al. RNA is a key component of extracellular DNA networks in Pseudomonas aeruginosa biofilms. Nat. Commun. 2023, 14, 7772. [Google Scholar] [CrossRef]
- Chiba, A.; Seki, M.; Suzuki, Y.; Kinjo, Y.; Mizunoe, Y.; Sugimoto, S. Staphylococcus aureus utilizes environmental RNA as a building material in specific polysaccharide-dependent biofilms. Npj Biofilms Microbiomes 2022, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A.; Gollan, B.; Helaine, S. Persistent bacterial infections and persister cells. Nat. Rev. Microbiol. 2017, 15, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Schilcher, K.; Horswill, A.R. Staphylococcal Biofilm Development: Structure, Regulation, and Treatment Strategies. Microbiol. Mol. Biol. Rev. 2020, 84, e00026. [Google Scholar] [CrossRef] [PubMed]
- Karimaei, S.; Kazem Aghamir, S.M.; Foroushani, A.R.; Pourmand, M.R. Antibiotic tolerance in biofilm persister cells of Staphylococcus aureus and expression of toxin-antitoxin system genes. Microb. Pathog. 2021, 159, 105126. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed]
- Guay, D.R. An Update on the Role of Nitrofurans in the Management of Urinary Tract Infections. Drugs 2001, 61, 353–364. [Google Scholar] [CrossRef]
- McOsker, C.C.; Fitzpatrick, P.M. Nitrofurantoin: Mechanism of action and implications for resistance development in common uropathogens. J. Antimicrob. Chemother. 1994, 33, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Le, V.V.H.; Rakonjac, J. Nitrofurans: Revival of an “old” drug class in the fight against antibiotic resistance. PLOS Pathog. 2021, 17, e1009663. [Google Scholar] [CrossRef]
- Rice, A.M.; Long, Y.; King, S.B. Nitroaromatic Antibiotics as Nitrogen Oxide Sources. Biomolecules 2021, 11, 267. [Google Scholar] [CrossRef]
- Choi, M.; Hasan, N.; Cao, J.; Lee, J.; Hlaing, S.P.; Yoo, J.-W. Chitosan-based nitric oxide-releasing dressing for anti-biofilm and in vivo healing activities in MRSA biofilm-infected wounds. Int. J. Biol. Macromol. 2020, 142, 680–692. [Google Scholar] [CrossRef]
- Ghaffari, A.; Miller, C.C.; McMullin, B.; Ghahary, A. Potential application of gaseous nitric oxide as a topical antimicrobial agent. Nitric Oxide 2006, 14, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Worley, B.V.; Schilly, K.M.; Schoenfisch, M.H. Anti-Biofilm Efficacy of Dual-Action Nitric Oxide-Releasing Alkyl Chain Modified Poly(amidoamine) Dendrimers. Mol. Pharm. 2015, 12, 1573–1583. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, C.; dos Santos, L.S.; Franco, O.L. Fosfomycin and nitrofurantoin: Classic antibiotics and perspectives. J. Antibiot. 2021, 74, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Koenig, H.N.; Durling, G.M.; Walsh, D.J.; Livinghouse, T.; Stewart, P.S. Novel Nitro-Heteroaromatic Antimicrobial Agents for the Control and Eradication of Biofilm-Forming Bacteria. Antibiotics 2021, 10, 855. [Google Scholar] [CrossRef] [PubMed]
- Mahdizade Ari, M.; Dashtbin, S.; Ghasemi, F.; Shahroodian, S.; Kiani, P.; Bafandeh, E.; Darbandi, T.; Ghanavati, R.; Darbandi, A. Nitrofurantoin: Properties and potential in treatment of urinary tract infection: A narrative review. Front. Cell. Infect. Microbiol. 2023, 13, 1148603. [Google Scholar] [CrossRef] [PubMed]
- Esposito, E.; Cuzzocrea, S. Role of nitroso radicals as drug targets in circulatory shock. Br. J. Pharmacol. 2009, 157, 494–508. [Google Scholar] [CrossRef] [PubMed]
- Verderosa, A.D.; Harris, J.; Dhouib, R.; Totsika, M.; Fairfull-Smith, K.E. Eradicating uropathogenic Escherichia coli biofilms with a ciprofloxacin-dinitroxide conjugate. MedChemComm 2019, 10, 699–711. [Google Scholar] [CrossRef]
- Das, B.; Rudra, S.; Yadav, A.; Ray, A.; Rao, A.V.S.R.; Srinivas, A.S.S.V.; Soni, A.; Saini, S.; Shukla, S.; Pandya, M.; et al. Synthesis and SAR of novel oxazolidinones: Discovery of ranbezolid. Bioorganic Med. Chem. Lett. 2005, 15, 4261–4267. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2018. [Google Scholar]
- Ceri, H.; Olson, M.E.; Stremick, C.; Read, R.R.; Morck, D.; Buret, A. The Calgary Biofilm Device: New technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J. Clin. Microbiol. 1999, 37, 1771–1776. [Google Scholar] [CrossRef]
- Innovotech. MBEC Assay® Procedural Manual for High-Throughput Antimicrobial Susceptibility Testing of Biofilms; Version 2.2; Innovotech: Edmonton, AB, Canada, 2021; p. 14. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | MIC (µg/mL) | MBEC99.9 a (µg/mL) | MBECC b (µg/mL) |
|---|---|---|---|
| Linezolid | 2 | 8 | >1024 |
| Tedizolid | 0.5 | 4 | >512 |
| Ranbezolid | 1 | 2 | 16 |
| Radezolid | 1 | 1 | >1024 |
| Deacetyl Linezolid Thioacetamide | 1 | 1 | >1024 |
| C-TEMPO | 4096 | >2048 | >2048 |
| Compound | MBECC (µg/mL) | FBEC | DRI |
|---|---|---|---|
| Ranbezolid | 16 | 0.25 | 4 |
| C-TEMPO | >2048 | 0.0039 | 256 |
| Ranbezolid + C-TEMPO | - | 0.2539 a | - |
| Compound | MW a (g/mol) | logP b | H Bond Donors c | H Bond Acceptors d | Rotatable Bonds e | TPSA f (Å2) |
|---|---|---|---|---|---|---|
| Linezolid | 337.35 | 1.26 | 5 | 1 | 5 | 71.11 |
| Tedizolid | 370.34 | 1.47 | 8 | 1 | 4 | 106.26 |
| Ranbezolid | 461.44 | 1.24 | 8 | 1 | 8 | 124.08 |
| Radezolid | 438.45 | 2.07 | 7 | 3 | 9 | 112.24 |
| Deacetyl Linezolid Thioacetamide | 353.41 | 1.95 | 4 | 1 | 5 | 86.13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndukwe, A.R.N.; Qin, J.; Wiedbrauk, S.; Boase, N.R.B.; Fairfull-Smith, K.E.; Totsika, M. In Vitro Activities of Oxazolidinone Antibiotics Alone and in Combination with C-TEMPO against Methicillin-Resistant Staphylococcus aureus Biofilms. Antibiotics 2023, 12, 1706. https://doi.org/10.3390/antibiotics12121706
Ndukwe ARN, Qin J, Wiedbrauk S, Boase NRB, Fairfull-Smith KE, Totsika M. In Vitro Activities of Oxazolidinone Antibiotics Alone and in Combination with C-TEMPO against Methicillin-Resistant Staphylococcus aureus Biofilms. Antibiotics. 2023; 12(12):1706. https://doi.org/10.3390/antibiotics12121706
Chicago/Turabian StyleNdukwe, Audrey R. N., Jilong Qin, Sandra Wiedbrauk, Nathan R. B. Boase, Kathryn E. Fairfull-Smith, and Makrina Totsika. 2023. "In Vitro Activities of Oxazolidinone Antibiotics Alone and in Combination with C-TEMPO against Methicillin-Resistant Staphylococcus aureus Biofilms" Antibiotics 12, no. 12: 1706. https://doi.org/10.3390/antibiotics12121706