
Use of the Naturally Occurring Bacteriophage Grouping Model for the Design of Potent Therapeutic Cocktails

 , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Isolation of Phages
2.2. Phage Plaque Purification and Morphology Characterization
2.3. Phage Host Range Study
2.4. Appelmans Assay
2.5. Lytic Activity of Phages and Phage Cocktails
2.5.1. Design of Phage Cocktails
2.5.2. Selection of Bacterial Strains for Testing the Activity of Single Phages and Cocktails
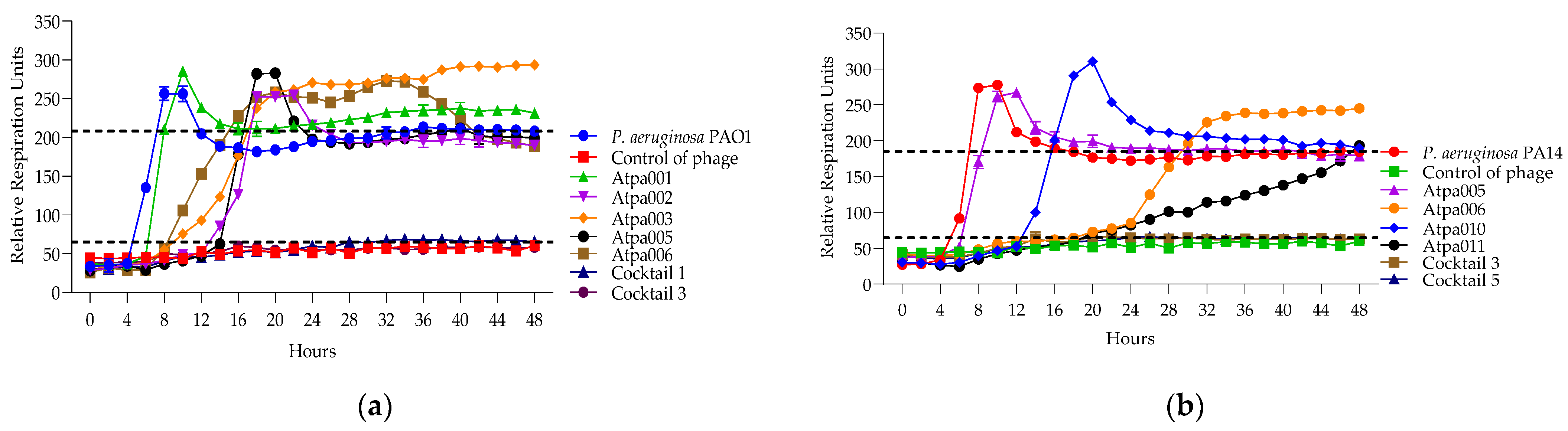
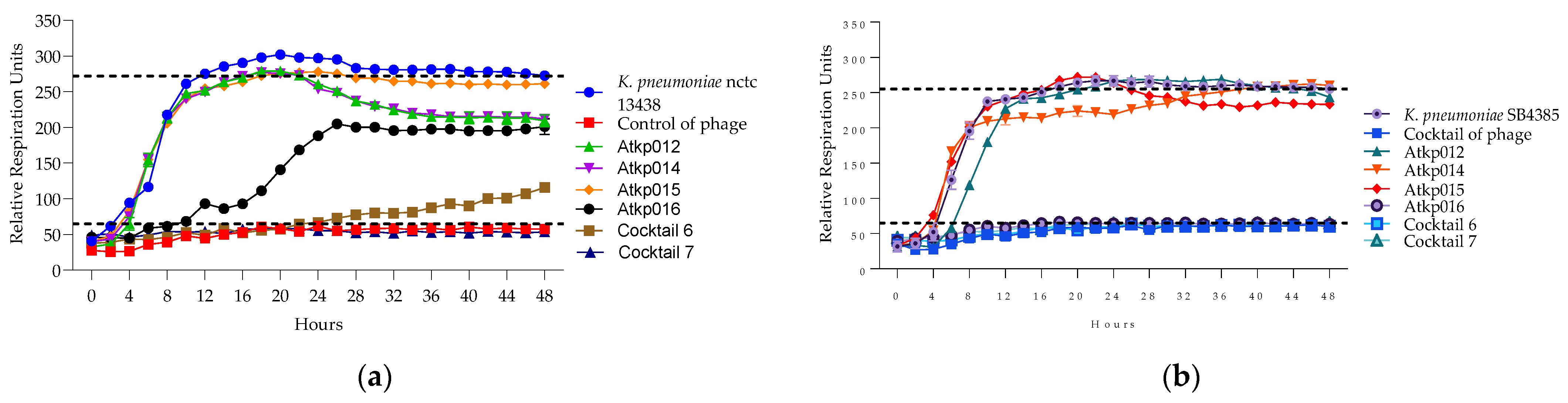
2.5.3. Developing a Model to Interpret Bacterial Proliferation and Phage Activity Curves
2.5.4. Lytic Activity of Individual Phages and Phage Cocktails
2.5.5. Bacteria and Phage Enumeration
3. Discussion
- The natural groups (cocktails) of phages isolated from the same environmental source (urban and hospital sewage or rivers) are quite different. For P. aeruginosa, 6/7 phage cocktails were composed of phages belonging to different genera, while for K. pneumoniae, this was the case for 3/7 phage cocktails.
- The natural groups (cocktails) of phages have relatively broad host range, as at least two of them showed in vitro clearing effect on P. aeruginosa strains of PAO1K, CN573, PAV237, PAO1, A11, Is573 and Is580 with the different virulence factors, including three resistotypes of PAO1, PA7 and PA14 and on K. pneumoniae strains with five different capsular serotypes and one with KPC-3.
- The natural groups (cocktails) of phages largely performed well, inhibiting PRM growth either in synergy or in proto-cooperation. Each cocktail showed a killing effect against at least two non-propagating strains.
- —
- P. aeruginosa phages largely suppress in vitro PRM (phage-resistant mutant) growth, either synergistically or in proto-cooperation.
- —
- K. pneumoniae phages overcome the inhibitory effect of single phages, resulting in synergy or proto-cooperation.
4. Materials and Methods
4.1. Bacterial growth
4.1.1. Bacterial Growth Media
4.1.2. Bacterial Growth Methods and Conditions
4.1.3. Biological Materials
4.2. Bacterial Enumeration (cfu) and Adjustment to OD
4.3. Phage Isolation
4.4. Phage Detection—Preliminary Test
4.5. Phage Activity Detection—Phage Plaque Formation and Enumeration
4.6. Confirmatory Test for Phage Plaque Formation and Enumeration
4.7. Phage Host Range and Efficiency of Plating (EOP)
4.8. Phage Plaque Purification
4.9. Preparation of High Concentrated Phage Using Web Pattern Plates [27,28]
4.10. Phage DNA Isolation
4.11. Phage Genome Sequencing
4.12. Phage Liquid Culturing (PLC)
4.12.1. Appelmans’ Method
4.12.2. Phage (or Mixture) Lytic Activity Evaluation Using Phage Liquid Culturing (PLC)
4.12.3. Enumeration of Bacteria and Phage upon Completion of the OmniLog Experiments (Figure 18)
4.13. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haines, M.E.K.; Hodges, F.E.; Nale, J.Y.; Mahony, J.; van Sinderen, D.; Kaczorowska, J.; Alrashid, B.; Akter, M.; Brown, N.; Sauvageau, D.; et al. Analysis of Selection Methods to Develop Novel Phage Therapy Cocktails Against Antimicrobial Resistant Clinical Isolates of Bacteria. Front. Microbiol. 2021, 12, 613529. [Google Scholar] [CrossRef] [PubMed]
- WHO Regional Office for Europe/European Centre for Disease Prevention and Control. Antimicrobial Resistance Surveillance in Europe 2022–2020 Data. Copenhagen: WHO Regional Office for Europe. 2022. Available online: https://www.ecdc.europa.eu/en/publications-data/antimicrobial-resistance-surveillance-europe-2022-2020-data (accessed on 17 April 2024).
- European Centre for Disease Prevention and Control. Surveillance of antimicrobial resistance in Europe 2018. Stockholm: ECDC. 2019. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/surveillance-antimicrobial-resistance-Europe-2018.pdf (accessed on 17 April 2024).
- Nikolich, M.P.; Filippov, A.A. Bacteriophage Therapy: Developments and Directions. Antibiotics 2020, 9, 135. [Google Scholar] [CrossRef] [PubMed]
- Pirnay, J.P.; Djebara, S.; Steurs, G.; Griselain, J.; Cochez, C.; De Soir, S.; Glonti, T.; Spiessens, A.; Vanden Berghe, E.; Green, S.; et al. Retrospective, observational analysis of the first one hundred consecutive cases of personalized bacteriophage therapy of difficult-to-treat infections facilitated by a Belgian consortium. medRxiv 2023. [Google Scholar] [CrossRef]
- Chan, B.K.; Turner, P.E.; Kim, S.; Mojibian, H.R.; Elefteriades, J.A.; Narayan, D. Phage treatment of an aortic graft infected with Pseudomonas aeruginosa. Evolut. Med. Public Health 2018, 2018, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Forti, F.; Roach, D.R.; Cafora, M.; Pasini, M.E.; Horner, D.S.; Fiscarelli, E.V.; Rossitto, M.; Cariani, L.; Briani, F.; Debarbieux, L.; et al. Design of a Broad-Range Bacteriophage Cocktail That Reduces Pseudomonas aeruginosa Biofilms and Treats Acute Infections in Two Animal Models. Antimicrob. Agents Chemother. 2018, 62, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Martins, W.M.B.S.; Li, M.; Sands, K.; Lenzi, M.H.; Portal, E.; Mathias, J.; Dantas, P.P.; Migliavacca, R.; Hunter, J.R.; Medeiros, E.A.; et al. Effective phage cocktail to combat the rising incidence of extensively drug-resistant Klebsiella pneumoniae sequence type 16. Emerg. Microbes Infect. 2022, 11, 1015–1023. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Liang, B.; Han, B.; Shi, Y.; Li, X.; Zhao, W.; Kastelic, J.; Gao, J. Effective of phage cocktail against Klebsiella pneumoniae infection of murine mammary glands. Microb. Pathog. 2023, 182, 106218. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.M.; Ribeiro, J.M.; Pereira, G.N.; Pereira, G.N.; Junior, I.D.; Junior, I.D.; Teixeira, G.M.; Teixeira, G.M.; Bertozzi, M.M.; Bertozzi, M.M.; et al. Comparative analysis of effectiveness for phage cocktail development against multiple Salmonella serovars and its biofilm control activity. Sci. Rep. 2023, 13, 13054. [Google Scholar] [CrossRef]
- Guerrero-Bustamante, C.A.; Dedrick, R.M.; Garlena, R.A.; Russell, D.A.; Hatfull, G.F. Toward a Phage Cocktail for Tuberculosis: Susceptibility and Tuberculocidal Action of Mycobacteriophages against Diverse Mycobacterium tuberculosis Strains. mBio 2021, 12, e00973-21. [Google Scholar] [CrossRef]
- Naknaen, A.; Samernate, T.; Wannasrichan, W.; Surachat, K.; Nonejuie, P.; Chaikeeratisak, V. Combination of genetically diverse Pseudomonas phages enhances the cocktail efficiency against bacteria. Sci. Rep. 2023, 13, 8921. [Google Scholar] [CrossRef]
- Abedon, S.T.; Danis-Wlodarczyk, K.M.; Wozniak, D.J. Phage Cocktail Development for Bacteriophage Therapy: Toward Improving Spectrum of Activity Breadth and Depth. Pharmaceuticals 2021, 14, 1019. [Google Scholar] [CrossRef]
- Markwitz, P.; Lood, C.; Olszak, T.; van Noort, V.; Lavigne, R.; Drulis-Kawa, Z. Genome-driven elucidation of phage-host interplay and impact of phage resistance evolution on bacterial fitness. ISME J. 2022, 16, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Behzadi, P.; Baráth, Z.; Gajdács, M. It’s Not Easy Being Green: A Narrative Review on the Microbiology, Virulence and Therapeutic Prospects of Multidrug-Resistant Pseudomonas aeruginosa. Antibiotics 2021, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Reboud, E.; Basso, P.; Maillard, A.P.; Huber, P.; Attrée, I. Exolysin Shapes the Virulence of Pseudomonas aeruginosa Clonal Outliers. Toxins 2017, 9, 364. [Google Scholar] [CrossRef] [PubMed]
- Pirnay, J.-P.; Bilocq, F.; Pot, B.; Cornelis, P.; Zizi, M.; Van Eldere, J.; Deschaght, P.; Vaneechoutte, M.; Jennes, S.; Pitt, T.; et al. Pseudomonas aeruginosa Population Structure Revisited. PLoS ONE 2009, 4, e7740. [Google Scholar] [CrossRef] [PubMed]
- Grace, A.; Sahu, R.; Owen, D.R.; Dennis, V.A. Pseudomonas aeruginosa reference strains PAO1 and PA14: A genomic, phenotypic, and therapeutic review. Front. Microbiol. 2022, 13, 1023523. [Google Scholar] [CrossRef] [PubMed]
- Imbert, P.R.; Louche, A.; Luizet, J.; Grandjean, T.; Bigot, S.; Wood, T.E.; Gagné, S.; Blanco, A.; Wunderley, L.; Terradot, L.; et al. A Pseudomonas aeruginosa TIR effector mediates immune evasion by targeting UBAP1 and TLR adaptors. EMBO J. 2017, 36, 1869–1887. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Stapleton, F.; Summers, S.; Rice, S.A.; Willcox, M.D.P. Antibiotic Resistance Characteristics of Pseudomonas aeruginosa Isolated from Keratitis in Australia and India. Antibiotics 2020, 9, 600. [Google Scholar] [CrossRef] [PubMed]
- Subedi, D.; Vijay, A.K.; Kohli, G.S.; Rice, S.A.; Willcox, M. Association between possession of ExoU and antibiotic resistance in Pseudomonas aeruginosa. PLoS ONE 2018, 13, e0204936. [Google Scholar] [CrossRef]
- Ammann, C.G.; Nagl, M.; Nogler, M.; Coraça-Huber, D.C. Pseudomonas aeruginosa outcompetes other bacteria in the manifestation and maintenance of a biofilm in polyvinylchloride tubing as used in dental devices. Arch. Microbiol. 2016, 198, 389–391. [Google Scholar] [CrossRef]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.L.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M.; et al. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Chibeu, A.; Ceyssens, P.-J.; Hertveldt, K.; Volckaert, G.; Cornelis, P.; Matthijs, S.; Lavigne, R. The adsorption of Pseudomonas aeruginosa bacteriophage phiKMV is dependent on expression regulation of type IV pili genes. FEMS Microbiol. Lett. 2009, 296, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Antoine, C.; Laforêt, F.; Blasdel, B.; Glonti, T.; Kutter, E.; Pirnay, J.; Mainil, J.; Delcenserie, V.; Thiry, D. Efficacy assessment of PEV2 phage on Galleria mellonella larvae infected with a Pseudomonas aeruginosa dog otitis isolate. Res. Veter Sci. 2021, 136, 598–601. [Google Scholar] [CrossRef] [PubMed]
- Hazan, R.; Que, Y.-A.; Maura, D.; Rahme, L.G. A method for high throughput determination of viable bacteria cell counts in 96-well plates. BMC Microbiol. 2012, 12, 259. [Google Scholar] [CrossRef] [PubMed]
- Glonti, T.; Pirnay, J.-P. In Vitro Techniques and Measurements of Phage Characteristics That Are Important for Phage Therapy Success. Viruses 2022, 14, 1490. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.D. Bacteriophages; Interscience Publishers: New York, NY, USA, 1959. [Google Scholar]
- Jakočiūnė, D.; Moodley, A. A Rapid Bacteriophage DNA Extraction Method. Methods Protoc. 2018, 1, 27. [Google Scholar] [CrossRef]
- Eskenazi, A.; Lood, C.; Wubbolts, J.; Hites, M.; Balarjishvili, N.; Leshkasheli, L.; Askilashvili, L.; Kvachadze, L.; van Noort, V.; Wagemans, J.; et al. Combination of pre-adapted bacteriophage therapy and antibiotics for treatment of fracture-related infection due to pandrug-resistant Klebsiella pneumoniae. Nat. Commun. 2022, 13, 302. [Google Scholar] [CrossRef]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phage Name | Isolation Host Strains (P. aeruginosa) | Isolation Source | Plaque Morphology | Genome Size (bp) | Genus |
|---|---|---|---|---|---|
| Atpa001 | PAO1K ATCC 15692 DSM 22644 | Brussels International Airport sewage water | 2 mm Ø (diameter) plaque | 66,468 | Hollowayvirus |
| Atpa002 | PAO1K ATCC 15692 DSM 22644 | Brussels International Airport sewage water | 2 mm Ø plaque | 65,922 | Pbunavirus |
| Atpa003 | PAO1K ATCC 15692 DSM 22644 | Brussels International Airport sewage water | 1 mm Ø clear plaque with halo | 60,912 | Yuavirus |
| Atpa004 | PAO1K ATCC 15692 DSM 22644 | Aquafin, Brussels, Belgium | 4 mm Ø plaque with size polymorphism, clear center and irregular margin of 2.5 mm | 72,290 | Litunavirus |
| Atpa005 | PAO1K ATCC 15692 DSM 22644 | Academic Hospital Monika, Brussels, Belgium | 4 mm Ø plaque with size polymorphism, clear center and irregular margin of 2.5 mm | 72,348 | Litunavirus |
| Atpa006 | PAO1K ATCC 15692 DSM 22644 | Academic Hospital Monika, Brussels, Belgium | 1 mm Ø translucent plaque with irregular margin | 45,608 | Bruynoghevirus |
| Atpa008 | PAO1K ATCC 15692 DSM 22644 | Academic Hospital Monika, Brussels, Belgium | 2 mm Ø shiny turbid plaque with irregular margin | 93,506 | Samunavirus |
| Atpa009 | CN573 DSM 22644 Eliava IBMV Collection CN573 | Saint-Jean hospital, Brussels, Belgium | 3 mm Ø plaque with clear center, size polymorphism and an irregular margin of 2 mm width | 71,982 | Litunavirus |
| Atpa010 | PAO1K ATCC 15692 DSM 22644 | River water from Nepal | 6–7 mm Ø clear plaque with clear center and an irregular margin of 1 mm width | 43,381 | Phikmvvirus |
| Atpa011 | PAO1K ATCC 15692 DSM 22644 | River water from Nepal | 6 mm Ø clear plaque with clear center and an irregular margin | Contig 1, 283,284 Contig 2, 43,363 | Phikzvirus Phikmvvirus |
| Atpa012 | PAO1K ATCC 15692 DSM 22644 | River water from Congo | 1 mm Ø plaque with size polymorphism | 61,551 | Yuavirus |
| Atpa013 | Eliava IBMV Collection CN573 | River water from Congo | 0.85 mm Ø plaque | 61,750 | Yuavirus |
| Atpa014 | CN573 DSM 22644 Eliava IBMV Collection CN573 | River water from Congo | 4 mm Ø plaque with 0.85 mm Ø clear center | 45,479 | Bruynoghevirus |
| Qatpa008 | PAV237 Olympia, WA, USA | QAMH sewage water | 2 mm Ø translucent plaque with size polymorphism and an irregular margin | 63,083 | New genus (unclassified) |
| Qatpa009 | PAV237 Olympia, WA, USA | QAMH sewage water | Clear 1 mm Ø plaque with size polymorphism and an irregular margin | 58,469 | Maxdohrnvirus |
| Qatpa0010 | CN573 DSM 22644 Eliava IBMV Collection CN573 | QAMH sewage water | 1 mm Ø clear plaque | Contig 11, cov 6.4, 87,990 | Pakpunavirus |
| Phage Name | Isolation Host Strains (K. pneumoniae) | Isolation Source | Plaque Morphology | Genome Size (bp) | Genus |
|---|---|---|---|---|---|
| Atkp01 | LabMCT0682 | Brussels International Airport sewage water | 4 mm Ø plaque with size polymorphism, clear center and a margin of 3 mm width | 52,977 | New genus (Drexlerviridae) |
| Atkp04 | LabMCT0682 | Aquafin, Brussels, Belgium | 3 mm Ø plaque with clear center and an irregular margin of 2 mm width | 52,586 | New genus (Drexlerviridae) |
| Atkp06 | LabMCT0682 | Aquafin, Brussels, Belgium | 1 mm Ø translucent plaque | 110,813 | Sugarlandvirus |
| Atkp07 | LabMCT0682 | Academic Hospital Monika, Brussels, Belgium | 1.5 mm Ø plaque with clear center | 115,044 | Sugarlandvirus |
| Atkp08 | LabMCT0682 | Academic Hospital Monika, Brussels, Belgium | 1.5 mm Ø plaque with clear center and a margin of 0.5 mm width | 115,044 | Sugarlandvirus |
| Atkp09 | NCTC 13438 | Saint-Jean hospital, Brussels, Belgium | 1.5 mm Ø plaque with clear center and a margin of 0.5 mm width | 106,874 | Sugarlandvirus |
| Atkp10 | SB4385 | Saint-Jean hospital, Brussels, Belgium | 1 mm Ø plaque with clear center | 108,212 | Sugarlandvirus |
| Atkp12 | ATCC 27736 | River water from Nepal | 3 mm Ø plaque with clear center and an irregular margin of 2 mm width | 49,740 | Webervirus |
| Atkp14 | LabMCT0682 | River water from Congo | Ø1 mm clear plaque | 175,285 | Marfavirus |
| Atkp15 | LabMCT0682 | River water from Congo | Ø1 mm clear plaque | 176,764 | Pseudotevenvirus |
| Atkp16 | ATCC 27736 | River water from Congo | Ø1 mm with twisted undulate margin | 176,276 | Slopekvirus |
| Cocktail Number | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| Phage components and their genera | Atpa001 Hollowayvirus | Atpa001 | Atpa005 Litunavirus | Atpa008 Samunavirus | Atpa010 Phikmvvirus | Atpa012 Yuavirus | Qatpa008 New genus (unclassified) |
| Atpa002 Pbunavirus | Atpa003 | Atpa006 Bruynoghevirus | Atpa009 Litunavirus | Atpa011 Phikzvirus | Atpa013 Yuavirus | Qatpa009 Maxdohrnvirus | |
| Atpa003 Yuavirus | Atpa004 Litunavirus | n/a | n/a | n/a | Atpa014 Bruynoghevirus | Qatpa010 Pakpunavirus | |
| Isolation sources | Brussels International Airport sewage water (BIA) | BIA and Aquafin (A), Brussels, Belgium | Academic Hospital Monika, Brussels, Belgium | Saint-Jean hospital, Brussels, Belgium | River water from Nepal | River water from Congo | Queen Astrid Military Hospital sewage water |
| Cocktail Number | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| Phage component and their genera | Atkp001 New genus (Drexlerviridae) | Atkp001 | Atkp004 New genus (Drexlerviridae) | Atkp007 Sugarlandvirus | Atkp009 | Atkp012 Webervirus | Atkp014 Marfavirus |
| Atkp008 Sugarlandvirus | Atkp009 Sugarlandvirus | Atkp006 Sugarlandvirus | Atkp008 | Atkp010 Sugarlandvirus | Atkp016 Slopekvirus | Atkp015 Pseudotevenvirus | |
| n/a | Atpa004 | n/a | n/a | n/a | n/a | Atkp016 | |
| Isolation source | BIA and Academic hospital Monika, Brussels, Belgium | BIA and Saint-Jean hospital, Brussels, Belgium | Aquafin, Brussels, Belgium | Academic hospital Monika, Brussels, Belgium | Saint-Jean hospital, Brussels, Belgium | River waters from Nepal and Congo | River waters from Congo |
| P. aeruginosa Strain rru Characteristics/ Parameters | Cocktail 1 Phage Components | Cocktail 2 Phage Components | Cocktail 3 Phage Components | Cocktail 4 Phage Components | Cocktail 5 Phage Components | Cocktail 6 Phage Components | Cocktail 7 Phage Components |
|---|---|---|---|---|---|---|---|
| PAO1K Control—221 rru | Cl-32 h, 161 rru -- | Cl-42 h, 135 rru -- | Cl-48 h, --, -- | In-48 h, 135 rru -- | Cl-48 h, --, -- | Cl-48 h, --, -- | Cl-30 h, 235 rru PRM |
| O5-serotype T3SS ExoS, ExoY oprI A1 oprL B11 | Atpa001, Cl-12 h PRM, 319 rru (EOP 0.002) | Atpa001, Cl-12 h PRM, 319 rru (EOP 0.002) | Atpa005, Cl-28 h PRM, 283 rru (EOP 0.03) | Atpa008, Cl-10 h --, 208 rru (EOP 0.2) | Atpa010, Cl-48 h --, --, (EOP 0.06) | Atpa012, Cl-8 h PRM, 302 rru (EOP 0.007) | Qatpa008, Cl-30 h PRM, 245 rru (EOP 7.8) |
| Atpa002, Cl-24 h PRM, 265 rru (EOP 0.0005) | Atpa003, Cl-12 h PRM, 248 rru (EOP 0.01) | Atpa006, Cl-28 h --, 237 rru (EOP 0.002) | Atpa009, Cl-24 h PRM, 307 rru (EOP 0.22) | Atpa011, Cl-48 h --, --, (EOP 0.002) | Atpa013, Cl-12 h PRM, 285 rru (EOP 0.003) | Qatpa009, Cl-12 h PRM, 301 rru (EOP 0.32) | |
| Atpa003, Cl-12 h PRM, 248 rru (EOP 0.01) | Atpa004, Cl-28 h PRM, 341 rru (EOP 0.03) | n/a | n/a | n/a | Atpa014, Cl-16 h --, 235 rru (EOP 1.9) | Qatpa010, Cl-16 h PRM, 307 rru (EOP 0.022) | |
| Interpretation | Synergy | Synergy | Synergy | Antagonism | Proto- cooperation | Synergy | Proto- cooperation |
| PAO1 Control—208 rru | Cl-48 h, --, -- | n/a | Cl-48 h, --, -- | n/a | n/a | n/a | n/a |
| O5-serotype T3SS ExoS, ExoY oprI A1 oprL B11 | Atpa001, Cl-6 h --, 231 rru (EOP 0.002) | n/a | Atpa005, Cl-15 h --, 199 rru (EOP 0.03) | n/a | n/a | n/a | n/a |
| Atpa002, Cl-12 h --, 188 rru (EOP 0.0005) | n/a | Atpa006, Cl-8 h PRM, 188 rru (EOP 0.002) | n/a | n/a | n/a | n/a | |
| Atpa003, Cl-8 h PRM, 294 rru (EOP 0.01) | n/a | n/a | n/a | n/a | n/a | n/a | |
| Interpretation | Synergy | - | Synergy | n/a | n/a | n/a | n/a |
| CN573 Control—222 rru | Cl-48 h, --, -- | Cl-48 h, --, -- | Cl-48 h, --, -- | Cl-48 h, --, -- | Cl-48 h, --, -- | Cl-48 h, --, -- | Cl-20 h, 187 rru -- |
| O1-serotype oprI B1 oprL B09 ExoS | Atpa001, Cl-28 h --, 234 rru (EOP 0.002) | Atpa001, Cl-28 h --, 234 rru (EOP 0.002) | Atpa005m Cl-48 h --, --, (EOP 0.03) | Atpa008, Cl-18 h --, 189 rru (EOP 0.2) | Atpa010, Cl-48 h --, --, (EOP 0.06) | Atpa012, ln-36 h --, 204 rru (EOP 0.007) | Qatpa008, Cl-28 h --, 122 rru (EOP 7.8) |
| Atpa002, Cl-36 h --, 225 rru (EOP 0.0005) | Atpa003, Cl-28 h PRM, 265 rru (EOP 0.01) | Atpa006, Cl-48 h --, --, (EOP 0.002) | Atpa009, Cl-48 h --, --, (EOP 0.22) | Atpa011, Cl-48 h --, --, (EOP 0.002) | Atpa013, Cl-30 h PRM, 248 rru (EOP 0.003) | Qatpa009, Cl-12 h PRM, 252 rru (EOP 0.32) | |
| Atpa003, Cl-28 h PRM, 265 rru (EOP 0.01) | Atpa004, Cl-48 h --, --, (EOP 0.03) | n/a | n/a | n/a | Atpa014, Cl-14 h --, 265 rru (EOP 1.9) | Qatpa010, Cl-48 h --, --, (EOP 0.022) | |
| Interpretation | Synergy | Proto-cooperation | Proto- cooperation | Proto- cooperation | Proto- cooperation | Synergy | Antagonism |
| PAV237 Control—228 rru | Cl-20 h, 255 rru PRM | Cl-48 h, --, -- | Cl-48 h, --, -- | Cl-48 h, --, -- | Cl-48 h, --, -- | Cl-48 h, --, -- | Cl-8 h, 296 rru PRM |
| lasB gene coding for elastase B | Atpa001, Cl-20 h --, 224 rru (EOP 0.002) | Atpa001, Cl-20 h --, 224 rru (EOP 0.002) | Atpa005, Cl-48 h --, (EOP 0.03) | Atpa008, Cl-12 h --, 213 rru (EOP 0.2) | Atpa010, Cl-14 h --, 209 rru (EOP 0.06) | Atpa012, Cl-6 h --, 220 rru (EOP 0.007) | Qatpa008, Cl-20 h PRM, 271 rru (EOP 7.8) |
| Atpa002, Cl-10 h PRM, 263 rru (EOP 0.0005) | Atpa003, Cl-10 h PRM, 285 rru (EOP 0.01) | Atpa006, Cl-12 h --, 193 rru (EOP 0.002) | Atpa009, Cl-48 h --, --, (EOP 0.22) | Atpa011, Cl-48 h --, --, (EOP 0.002) | Atpa013, Cl-10 h --, 179 rru (EOP 0.003) | Qatpa009, Cl-8 h PRM, 295 rru (EOP0.32) | |
| Atpa003, Cl-10 h PRM, 285 rru (EOP 0.01) | Atpa004, Cl-48 h --, --, (EOP 0.03) | n/a | n/a | n/a | Atpa014, Cl-48 h --, --, (EOP 1.9) | Qatpa010, Cl-14 h --, 209 rru (EOP 0.022) | |
| Interpretation | Proto- cooperation | Proto- cooperation | Proto-cooperation | Proto- cooperation | Proto- cooperation | Proto- cooperation | Antagonism |
| PA14 Control—185 rru | n/a | n/a | Cl-48 h, --, -- | n/a | Cl-48 h, --, -- | n/a | n/a |
| O10-serotype, oprI B1 oprL A05 T3SS ExoU, ExoY | n/a | n/a | Atpa005, Cl-6 h --, 180 rru (EOP 0.03) | n/a | Atpa010, Cl-12 h --, 193 rru (EOP 0.06) | n/a | n/a |
| - | - | Atpa006, Cl-24 h PRM, 245 rru (EOP 0.002) | - | Atpa011, Cl-26 h PRM, 193 rru (EOP 0.002) | - | - | |
| Interpretation | - | - | Synergy | - | Synergy | - | - |
| Is573 Control—248 rru | n/a | n/a | n/a | n/a | n/a | Cl-48 h, --, -- | Cl-30 h, 205 rru PRM |
| O11-serotype, oprI B1 oprL B02 ExoU | n/a | n/a | n/a | n/a | n/a | Atpa012, Cl-6 h --, 239 rru (EOP 0.007) | Qatpa008, Cl-16 h --, 225 rru (EOP 7.8) |
| n/a | n/a | n/a | n/a | n/a | Atpa013, Cl-8 h --, 257 rru (EOP 0.003) | Qatpa009, Cl-14 h --, 242 rru (EOP 0.32) | |
| n/a | n/a | n/a | n/a | n/a | Atpa014, Cl-48 h --, --, (EOP 1.9) | Qatpa010, Cl-8 h --, 249 rru (EOP 0.022) | |
| Interpretation | - | - | - | - | - | Proto- cooperation | Synergy |
| A11 Control—241 rru | Cl-28 h, 216 rru -- | n/a | n/a | n/a | Cl-30 h, 210 rru -- | n/a | n/a |
| oprI B1 oprL B12 ExoS | Atpa001, --, --, 250 rru (EOP 0.002) | n/a | n/a | n/a | Atpa010, Cl-6 h --, 250 rru (EOP 0.06) | n/a | n/a |
| Atpa002, Cl-6 h --, 198 rru (EOP 0.0005) | n/a | n/a | n/a | Atpa011, Cl-26 h --, 252 rru (EOP 0.002) | n/a | n/a | |
| Atpa003, Cl-8 h --, 197 rru (EOP 0.01) | n/a | n/a | n/a | n/a | n/a | n/a | |
| Interpretation | Synergy | n/a | n/a | n/a | Synergy | n/a | n/a |
| Is580 Control—222 rru | n/a | n/a | n/a | Cl-44 h, 148 rru | Cl-48 h | n/a | n/a |
| O3-serotype oprI B1 oprL B03 ExoS | n/a | n/a | n/a | Atpa008, Cl-6 h --, 188 rru (EOP 0.2) | Atpa010, Cl-10 h --, 213 rru (EOP 0.06) | n/a | n/a |
| n/a | n/a | n/a | Atpa009, Cl-10 h --, 193 rru (EOP 0.22) | Atpa011, Cl-48 h --, --, (EOP 0.002) | n/a | n/a | |
| Interpretation | n/a | n/a | n/a | Synergy | Proto- cooperation | n/a | n/a |
| PA7 Control—261 rru | Cl-10 h, 246 rru -- | --, 298 rru PRM | --, 212 rru -- | --, 283 rru PRM | Cl-14 h, 296 rru PRM | Cl-8 h, 247 rru -- | Cl-10 h, 297 rru PRM |
| MDR O12-serotype TPS ExlA oprI F1 oprL E03 | Atpa001, --, PRM, 280 rru (EOP 0.002) | Atpa001, --, --, 280 rru (EOP 0.002) | Atpa005 --, 256 rru (EOP 0.03) | Atpa008, --, --, 236 rru (EOP 0.2) | Atpa010, --, PRM, 275 rru (EOP 0.06) | Atpa012, --, PRM, 287 rru (EOP 0.007) | Qatpa008, Cl-20 h PRM, 275 rru (EOP 7.8) |
| Atpa002, --, --, 255 rru (EOP 0.0005) | Atpa003, --, PRM, 284 rru (EOP 0.01) | Atpa006, --, --, 208 rru (EOP 0.002) | Atpa009, --, --, 227 rru (EOP 0.22) | Atpa011, Cl-14 h PRM, 305 rru (EOP 0.002) | Atpa013, --, PRM, 288 rru (EOP 0.003) | Qatpa009, --, --, 236 rru (EOP 0.32) | |
| Atpa003, --, --, 284 rru (EOP 0.01) | Atpa004, --, --, 265 rru (EOP 0.03) | n/a | n/a | n/a | Atpa014, Cl-8 h --, 274 rru (EOP 1.9) | Qatpa010, Cl-10 h --, 297 rru (EOP 0.022) | |
| Interpretation | Synergy | Antagonism | - | Antagonism | Proto-cooperation | Proto- cooperation | Antagonism |
| K. pneumoniae Strains/ Capsule Type | Cocktail 1 Phage Components | Cocktail 2 Phage Components | Cocktail 3 Phage Components | Cocktail 4 Phage Components | Cocktail 5 Phage Components | Cocktail 6 Phage Components | Cocktail 7 Phage Components |
|---|---|---|---|---|---|---|---|
| LabMCT 0682 Control—192 rru | Cl-48 h, --, -- | Cl-48 h, --, -- | Cl-48 h, --, -- | Cl-48 h, --, -- | Cl-48 h, --, -- | Cl-48 h, --, -- | Cl-48 h, --, -- |
| Capsular serotype 81 | Atkp001, In-48 h --, 163 rru (EOP 0.08) | Atkp001, In-48 h --, 163 rru (EOP 0.08) | Atkp004, Cl-48 h --, --, (EOP 0.8) | Atkp007, ln-48 h --, 136 rru (EOP 0.08) | Atkp00,9 CL-48 h --, --, (EOP 0.013) | Atkp012, Cl-48 h --, --, (EOP 0.5) | Atkp014, Cl-48 h --, --, EOP 0.011) |
| Atkp008, In-48 h --, 16 rru (EOP 0.07) | Atkp009, In-48 h --, 106 rru (EOP 0.013) | Atkp006, In-48 h --, 180 rru (EOP 0.09) | Atkp008, In-48 h --, 116 rru (EOP 0.07) | Atkp010, Cl-48 h --, --, (EOP 1.02) | Atkp016, Cl-48 h --, --, (EOP 0.015) | Atkp015, In-48 h --, 112 rru (EOP 0.09) | |
| n/a | n/a | n/a | n/a | n/a | n/a | Atkp016, Cl-48 h --, --, (EOP 0.015) | |
| Interpretation | Synergy | Synergy | Proto-cooperation | Synergy | Proto-cooperation | Proto-cooperation | Proto-cooperation |
| Atcc 27736 Control—250 rru | n/a | Cl-8 h, 178 rru --, | n/a | n/a | n/a | Cl-48 h, --, -- | Cl-48 h, --, -- |
| Capsular serotype 30 | n/a | Atkp001, --, --, 250 rru (EOP 0.08) | n/a | n/a | n/a | Atkp012, Cl-48 h --, --, (EOP 0.5) | Atkp014, Cl-12 h --, 218 rru (EOP 0.011) |
| n/a | Atkp009, --, --, 250 rru (EOP 0.013) | n/a | n/a | n/a | Atkp016, Cl-48 h --, --, EOP 0.015 | Atkp015, --, --, --, (EOP 0.09) | |
| n/a | n/a | n/a | n/a | n/a | n/a | Atkp016, Cl-48 h --, --, (EOP 0.015) | |
| Interpretation | n/a | Synergy | n/a | n/a | n/a | Proto-cooperation | Proto-cooperation |
| nctc 13438 Control—272 rru | n/a | n/a | n/a | n/a | n/a | Cl-48 h, --, --, --, | Cl-48 h, --, --, --, |
| carbapenemase KPC-3 | n/a | n/a | n/a | n/a | n/a | Atkp012, --, --, --, (EOP 0.5) | Atkp014, --, --, --, (EOP 0.011) |
| n/a | n/a | n/a | n/a | n/a | Atkp016, Cl-16 h --, 201 rru (EOP 0.015) | Atkp015, --, --, --, (EOP 0.09) | |
| n/a | n/a | n/a | n/a | n/a | n/a | Atkp016, Cl-16 h --, 201 rru (EOP 0.015) | |
| interpretation | n/a | n/a | n/a | n/a | n/a | Synergy | Synergy |
| SB4385 Control—255 rru | n/a | n/a | n/a | n/a | n/a | Cl-48 h, --, --, | Cl-48 h, --, --, |
| Capsular serotype 1 | n/a | n/a | n/a | n/a | n/a | Atkp012, Cl-6 h --, --, (EOP 0.5) | Atkp014, --, --, --, (EOP 0.011) |
| n/a | n/a | n/a | n/a | n/a | Atkp016, Cl-48 h --, --, (EOP 0.015) | Atkp015, --, --, --, (EOP 0.09) | |
| n/a | n/a | n/a | n/a | n/a | n/a | Atkp016 Cl-48 h EOP 0.015 | |
| interpretation | n/a | n/a | n/a | n/a | n/a | Proto-cooperation | Proto-cooperation |
| 10394 Control—265 rru | n/a | n/a | n/a | n/a | n/a | In-48 h, 206 rru -- | In-48 h, 185 rru -- |
| Capsular serotype 62 | n/a | n/a | n/a | n/a | n/a | Atkp012, --, PRM, 341 rru (EOP 0.5) | Atkp014, --, --, 258 rru (EOP 0.011) |
| n/a | n/a | n/a | n/a | n/a | Atkp016, --, --, 266 rru (EOP 0.015) | Atkp015, --, --, 272 rru (EOP 0.09) | |
| n/a | n/a | n/a | n/a | n/a | n/a | Atkp016, --, --, 266 rru (EOP 0.015) | |
| n/a | n/a | n/a | n/a | n/a | Synergy | Synergy | |
| 70165 Control—065 rru | n/a | ln-48 h, 194 rru -- | ln-48 h, 202 rru -- | Cl-10 h, ln-48 h, --, 216 rru | In-48 h, 185 rru --, | n/a | n/a |
| Capsular serotype 2 | n/a | Atkp001, --, --, 224 rru (EOP 0.08) | Atkp004, ln-48 h --, 201 rru (EOP0.8) | Atkp007, Cl-8 h --, 257 rru (EOP 0.08) | Atkp010, In-48 h --, 185 rru (EOP 1.02) | n/a | n/a |
| n/a | Atkp009, --, --, 246 rru (EOP 0.013) | Atkp006, --, --, 246 rru (EOP 0.08) | Atkp008, Cl-8 h --, 243 rru (EOP 0.07) | Atkp009, --, --, 246 rru (EOP 0.013) | n/a | n/a | |
| interpretation | n/a | Synergy | Proto-cooperation | Synergy | Proto-cooperation | n/a | n/a |
| VPKP389 Control—294 rru | Cl-48 h, --, -- | Cl-28 h, --, -- | n/a | n/a | Cl-48 h, --, -- | n/a | n/a |
| Capsular serotype 27 | Atkp001, --, ---, --, (EOP 0.08) | Atkp001, --, --, --, (EOP0.08) | n/a | n/a | Atkp009, Cl-48 h --, --, (EOP0.013) | n/a | n/a |
| Atkp008 Cl-12 h EOP 0.08 | Atkp009 In-48 h EOP 0.013 | n/a | n/a | Atkp010, --, --, 275 rru (EOP 0.5) | n/a | n/a | |
| Interpretation | Synergy | Synergy | n/a | n/a | Proto-cooperation | n/a | n/a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glonti, T.; Goossens, M.; Cochez, C.; Green, S.; Gorivale, S.; Wagemans, J.; Lavigne, R.; Pirnay, J.-P. Use of the Naturally Occurring Bacteriophage Grouping Model for the Design of Potent Therapeutic Cocktails. Antibiotics 2024, 13, 385. https://doi.org/10.3390/antibiotics13050385
Glonti T, Goossens M, Cochez C, Green S, Gorivale S, Wagemans J, Lavigne R, Pirnay J-P. Use of the Naturally Occurring Bacteriophage Grouping Model for the Design of Potent Therapeutic Cocktails. Antibiotics. 2024; 13(5):385. https://doi.org/10.3390/antibiotics13050385
Chicago/Turabian StyleGlonti, Tea, Michael Goossens, Christel Cochez, Sabrina Green, Sayali Gorivale, Jeroen Wagemans, Rob Lavigne, and Jean-Paul Pirnay. 2024. "Use of the Naturally Occurring Bacteriophage Grouping Model for the Design of Potent Therapeutic Cocktails" Antibiotics 13, no. 5: 385. https://doi.org/10.3390/antibiotics13050385