Production of Antagonistic Compounds by Bacillus sp. with Antifungal Activity against Heritage Contaminating Fungi

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism and Culture Media
2.2. Bioactive Compound Production Study
2.2.1. Bacillus Growth Monitoring
2.2.2. Spore Germination Assay
2.3. Antifungal Inhibitory Capacity Assessment
2.3.1. Antifungal Paper Disk Diffusion Assay
2.3.2. In Vitro Assay with Marble Slabs
2.3.3. DNA Analysis
3. Results and Discussion
3.1. Biocompound Production Study
3.1.1. Cell Growth Dynamics of Bacillus sp. CCLBH 1053
3.1.2. Spore Germination Assessment
3.2. Antimicrobial Effect in Cultural Heritge Assets
3.2.1. Antifungal Inhibitory Capacity Assessment
3.2.2. Simulation Assays in Marble Materials
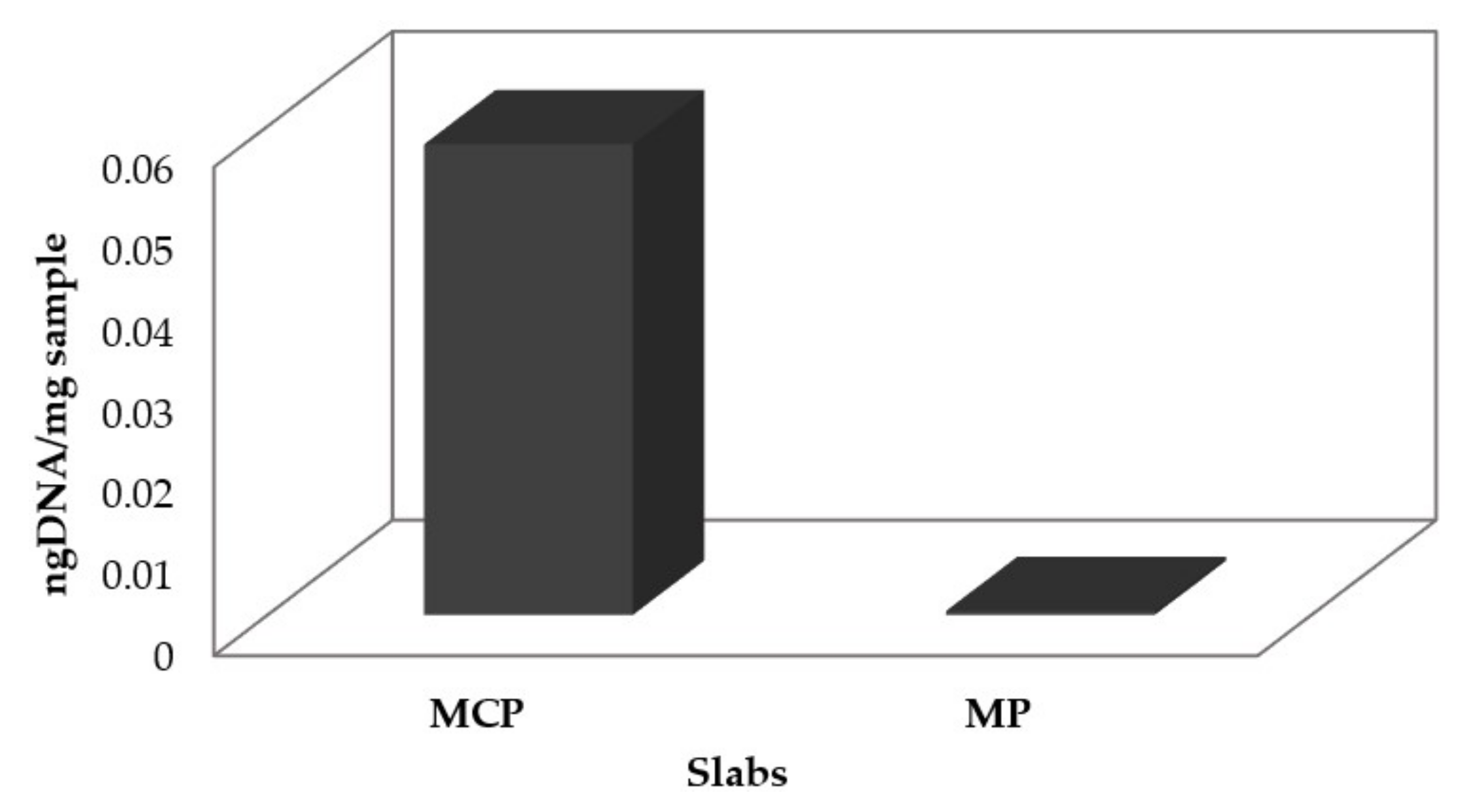
3.2.3. Marble Slab DNA
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Urzì, C.; De Leo, F. Evaluation of the efficiency of water-repellent and biocide compounds against microbial colonization of mortars. Int. Biodeterior. Biodegrad. 2007, 60, 25–34. [Google Scholar] [CrossRef]
- McNamara, C.J.; Mitchell, R. Microbial deterioration of historic stone. Front. Ecol. Environ. 2005, 3, 445–451. [Google Scholar] [CrossRef]
- Sterflinger, K.; Piñar, G. Microbial deterioration of cultural heritage and works of art—Tilting at windmills? Appl. Microbiol. Biotechnol. 2013, 97, 9637–9646. [Google Scholar] [CrossRef] [PubMed]
- Scheerer, S.; Ortega-Morales, O.; Gaylarde, C. Microbial deterioration of stone monuments—An updated overview. Adv. Appl. MIicrobiol. 2009, 66, 97–139. [Google Scholar]
- Rosado, T.; Gil, M.; Mirão, J.; Candeias, A.; Caldeira, A.T. Oxalate biofilm formation in mural paintings due to microorganisms—A comprehensive study. Int. Biodeterior. Biodegrad. 2013, 85, 1–7. [Google Scholar] [CrossRef]
- Silva, M.; Rosado, T.; Teixeira, D.; Candeias, A.; Caldeira, A.T. Production of green biocides for cultural heritage. Novel biotechnological solutions. Int. J. Conserv. Sci. 2015, 6, 519–530. [Google Scholar]
- Hegstad, K.; Langsrud, S.; Lunestad, B.T.; Scheie, A.A.; Sunde, M.; Yazdankhah, S.P. Does the wide use of quaternary ammonium compounds enhance the selection and spread of antimicrobial resistance and thus threaten our health? Microb. Drug Resist. 2010, 16, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Buffet-Bataillon, S.; Tattevin, P.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Emergence of resistance to antibacterial agents: The role of quaternary ammonium compounds—A critical review. Int. J. Antimicrob. Agents 2012, 39, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Caldeira, A.T.; Feio, S.S.; Santos Arteiro, J.M.; Roseiro, J.C. Bacillus amyloliquefaciens CCMI 1051 in vitro activity against wood contaminant fungi. Ann. Microbiol. 2007, 57, 29–33. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.F.; Gudiña, E.J.; Costa, R.; Vitorino, R.; Teixeira, J.A.; Coutinho, J.A.; Rodrigues, L.R. Optimization and characterization of biosurfactant production by Bacillus subtilis isolates towards microbial enhanced oil recovery applications. Fuel 2013, 111, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Raaijmakers, J.M.; De Bruijn, I.; Nybroe, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef] [PubMed]
- Ruangwong, O.-U.; Chang, C.-I.; Lamine, S.A.; Liang, W.-J. Identification of antifungal compound produced by Bacillus subtilis LB5 with ability to control anthracnose disease caused by Colletotrichum gloeosporioides. Afr. J. Microbiol. Res. 2012, 6, 3732–3738. [Google Scholar] [CrossRef]
- Rahman, M.S.; Ano, T.; Shoda, M. Second stage production of iturin A by induced germination of Bacillus subtilis RB14. J. Biotechnol. 2006, 125, 513–515. [Google Scholar] [CrossRef] [PubMed]
- Engelberg-Kulka, H.; Hazan, R. Cannibals defy starvation and avoid sporulation. Science 2003, 301, 467–468. [Google Scholar] [CrossRef] [PubMed]
- González-Pastor, J.E.; Hobbs, E.C.; Losick, R. Cannibalism by sporulating bacteria. Science 2003, 301, 510–513. [Google Scholar] [CrossRef] [PubMed]
- Piggot, P.J.; Hilbert, D.W. Sporulation of Bacillus subtilis. Curr. Opin. Microbiol. 2004, 7, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Caldeira, A.T.; Feio, S.S.; Arteiro, J.M.; Coelho, A.V.; Roseiro, J.C. Environmental dynamics of Bacillus amyloliquefaciens CCMI 1051 antifungal activity under different nitrogen patterns. J. Appl. Microbiol. 2008, 104, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, B.R.; Chakraborty, R.; Chaudhuri, U.R. Validity of modified Gompertz and Logistic models in predicting cell growth of Pediococcus acidilactici H during the production of bacteriocin pediocin AcH. J. Food Eng. 2007, 80, 1171–1175. [Google Scholar] [CrossRef]
- Zwietering, M.; Jongenburger, I.; Rombouts, F.; Van’t Riet, K. Modeling of the bacterial growth curve. Appl. Environ. Microbiol. 1990, 56, 1875–1881. [Google Scholar] [PubMed]
- Silva, M.; Pereira, A.; Teixeira, D.; Candeias, A.; Caldeira, A.T. Combined use of NMR, LC-ESI-MS and antifungal tests for rapid detection of bioactive lipopeptides produced by Bacillus. Adv. Microbiol. 2016, 6, 788–796. [Google Scholar] [CrossRef]
- Caldeira, A.T.; Santos Arteiro, J.M.; Coelho, A.V.; Roseiro, J.C. Combined use of LC–ESI-MS and antifungal tests for rapid identification of bioactive lipopeptides produced by Bacillus amyloliquefaciens CCMI 1051. Process. Biochem. 2011, 46, 1738–1746. [Google Scholar] [CrossRef]
- Dieckmann, R.; Pavela-Vrancic, M.; von Döhren, H. Synthesis of (di) adenosine polyphosphates by non-ribosomal peptide synthetases (NRPS). Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzymol. 2001, 1546, 234–241. [Google Scholar] [CrossRef]
- Yazgan, A.; Özcengiz, G.; Özcengiz, E.; Kılınç, K.; Marahiel, M.; Alaeddinoğlu, N.G. Bacilysin biosynthesis by a partially-purified enzyme fraction from Bacillus subtilis. Enzym. Microb. Technol. 2001, 29, 400–406. [Google Scholar] [CrossRef]
- Chevanet, C.; Besson, F.; Michel, G. Effect of various growth conditions on spore formation and bacillomycin L production in Bacillus subtilis. Can. J. Microbiol. 1986, 32, 254–258. [Google Scholar] [CrossRef] [PubMed]

 ) Normal growth in NB culture medium; (
) Normal growth in NB culture medium; (  ) NBPSHA assay; (
) NBPSHA assay; (  ) NBPS assay. I—Spore concentration baseline, II—Time with high spore concentration alteration.
) Normal growth in NB culture medium; ( ) NBPSHA assay; ( ) NBPS assay. I—Spore concentration baseline, II—Time with high spore concentration alteration.
) NBPS assay. I—Spore concentration baseline, II—Time with high spore concentration alteration.
) Normal growth in NB culture medium; ( ) NBPSHA assay; ( ) NBPS assay. I—Spore concentration baseline, II—Time with high spore concentration alteration.
 ) NBPSHA; (
) NBPSHA; (  ) NBPS assay.
) NBPSHA; ( ) NBPS assay.
) NBPS assay.
) NBPSHA; ( ) NBPS assay.




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, M.; Rosado, T.; Gonzalez-Pérez, M.; Gobbo, D.; Teixeira, D.; Candeias, A.; Caldeira, A.T. Production of Antagonistic Compounds by Bacillus sp. with Antifungal Activity against Heritage Contaminating Fungi. Coatings 2018, 8, 123. https://doi.org/10.3390/coatings8040123
Silva M, Rosado T, Gonzalez-Pérez M, Gobbo D, Teixeira D, Candeias A, Caldeira AT. Production of Antagonistic Compounds by Bacillus sp. with Antifungal Activity against Heritage Contaminating Fungi. Coatings. 2018; 8(4):123. https://doi.org/10.3390/coatings8040123
Chicago/Turabian StyleSilva, Mara, Tânia Rosado, Marina Gonzalez-Pérez, Débora Gobbo, Dora Teixeira, António Candeias, and Ana Teresa Caldeira. 2018. "Production of Antagonistic Compounds by Bacillus sp. with Antifungal Activity against Heritage Contaminating Fungi" Coatings 8, no. 4: 123. https://doi.org/10.3390/coatings8040123