
Dissecting Holistic Metabolic Acclimatization of Mucor circinelloides WJ11 Defective in Carotenoid Biosynthesis

 ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
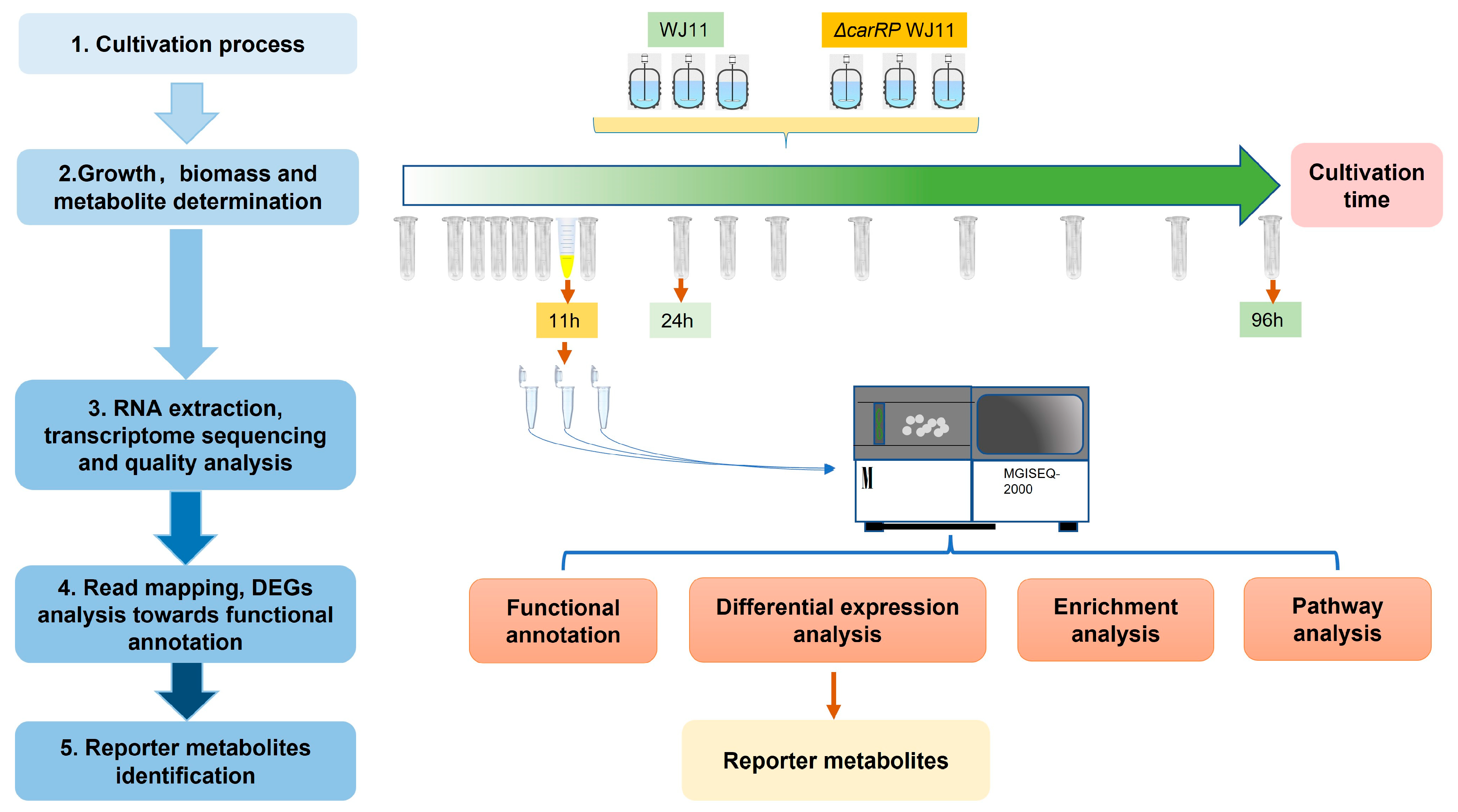
2.1. Fungal Strains and Cultivations
2.2. Biomass and Metabolite Determination
2.3. RNA Extraction, Transcriptome Sequencing, and Quality Analysis
2.4. Read Mapping and DEG Analysis for Functional Annotation
2.5. Reporter Metabolites Identification Based on the Genome-Scale Metabolic Model of M. circinelloides WJ11 (iNI1159)
3. Results and Discussion
3.1. Comparative Growth Profiles and Targeted Metabolite Traits of WJ11 and ΔcarRP WJ11 Strains
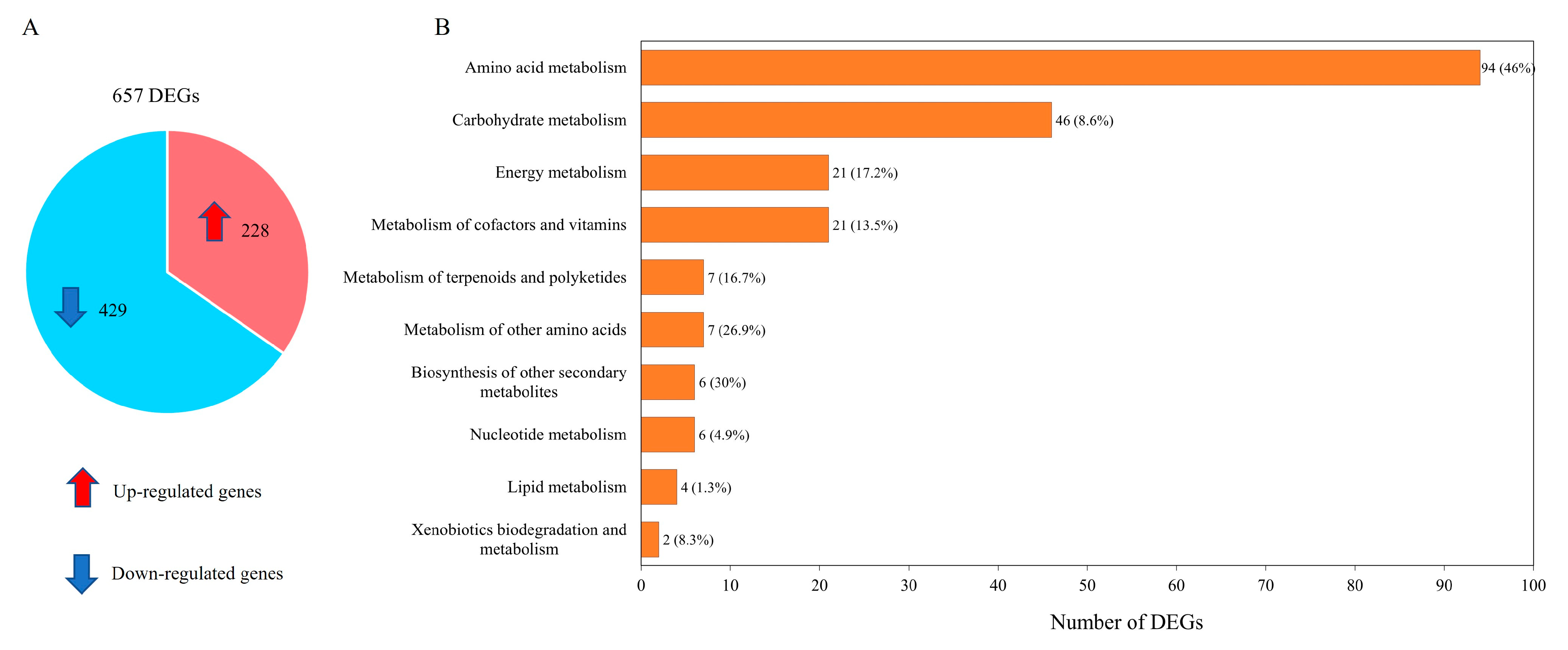
3.2. Transcriptome Data and DEGs across Pairwise Comparisons of WJ11 and ΔcarRP WJ11 Cultures
3.3. Metabolic Responses of Lipid and β-Carotene Biosynthetic Pathways in WJ11 and ΔcarRP WJ11 Strains
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ratledge, C. Fatty acid biosynthesis in microorganisms being used for Single Cell Oil production. Biochimie 2004, 86, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Lopes, H.J.S.; Bonturi, N.; Kerkhoven, E.J.; Miranda, E.A.; Lahtvee, P.J. C/N ratio and carbon source-dependent lipid production profiling in Rhodotorula toruloides. Appl. Microbiol. Biotechnol. 2020, 104, 2639–2649. [Google Scholar] [CrossRef] [PubMed]
- Moguel, I.S.; Yamakawa, C.K.; Pessoa, A., Jr.; Mussatto, S.I. L-asparaginase Production by Leucosporidium scottii in a Bench-Scale Bioreactor with Co-production of Lipids. Front. Bioeng. Biotechnol. 2020, 8, 576511. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.G.; Cohen, J.L.; Ozturk, G.; Hennebelle, M.; Taha, A.Y.; JM, L.N.d.M.B. Bioconversion of cheese whey permeate into fungal oil by Mucor circinelloides. J. Biol. Eng. 2018, 12, 25. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.Q.; Pratomo Juwono, N.K.; Leong, S.S.; Chang, M.W. Production of Fatty Acid-derived valuable chemicals in synthetic microbes. Front. Bioeng. Biotechnol. 2014, 2, 78. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Chen, H.; Chen, Y.Q.; Chen, W.; Garre, V.; Song, Y.; Ratledge, C. Comparison of biochemical activities between high and low lipid-producing strains of Mucor circinelloides: An explanation for the high oleaginicity of strain WJ11. PLoS ONE 2015, 10, e0128396. [Google Scholar] [CrossRef] [PubMed]
- Ratledge, C. Regulation of lipid accumulation in oleaginous micro-organisms. Biochem. Soc. Trans. 2002, 30, 1047–1050. [Google Scholar] [CrossRef] [PubMed]
- Naz, T.; Nosheen, S.; Li, S.; Nazir, Y.; Mustafa, K.; Liu, Q.; Garre, V.; Song, Y. Comparative Analysis of beta-Carotene Production by Mucor circinelloides Strains CBS 277.49 and WJ11 under Light and Dark Conditions. Metabolites 2020, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Strebanska, S.; Sajbidor, J. Oligounsaturated fatty acid production by selected strains of Micromycetes. Folia Microbiol. 1992, 37, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Luan, X.; Zhang, H.; Garre, V.; Song, Y.; Ratledge, C. Improved γ-linolenic acid production in Mucor circinelloides by homologous overexpressing of delta-12 and delta-6 desaturases. Microb Cell Fact. 2017, 16, 113. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Wang, W.; Yarbrough, J.M.; Baker, J.O.; Laurens, L.; Van Wychen, S.; Chen, X.; Taylor, L.E., 2nd; Xu, Q.; Himmel, M.E.; et al. Genomic, proteomic, and biochemical analyses of oleaginous Mucor circinelloides: Evaluating its capability in utilizing cellulolytic substrates for lipid production. PLoS ONE 2013, 8, e71068. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues Reis, C.E.; Bento, H.B.S.; Carvalho, A.K.F.; Rajendran, A.; Hu, B.; De Castro, H.F. Critical applications of Mucor circinelloides within a biorefinery context. Crit. Rev. Biotechnol. 2019, 39, 555–570. [Google Scholar] [CrossRef] [PubMed]
- Kot, A.M.; Blazejak, S.; Kurcz, A.; Gientka, I.; Kieliszek, M. Rhodotorula glutinis-potential source of lipids, carotenoids, and enzymes for use in industries. Appl. Microbiol. Biotechnol. 2016, 100, 6103–6117. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Jackson, E.N. Metabolic engineering of Yarrowia lipolytica for industrial applications. Curr. Opin. Biotechnol. 2015, 36, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Zhao, L.; Chen, H.; Chen, Y.Q.; Chen, W.; Song, Y.; Ratledge, C. Complete genome sequence of a high lipid-producing strain of Mucor circinelloides WJ11 and comparative genome analysis with a low lipid-producing strain CBS 277.49. PLoS ONE 2015, 10, e0137543. [Google Scholar] [CrossRef] [PubMed]
- Isarankura Na Ayudhya, N.; Laoteng, K.; Song, Y.; Meechai, A.; Vongsangnak, W. Metabolic traits specific for lipid-overproducing strain of Mucor circinelloides WJ11 identified by genome-scale modeling approach. PeerJ 2019, 7, e7015. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Zan, X.; Zhao, L.; Chen, H.; Chen, Y.Q.; Chen, W.; Song, Y.; Ratledge, C. Proteomics analysis of high lipid-producing strain Mucor circinelloides WJ11: An explanation for the mechanism of lipid accumulation at the proteomic level. Microb. Cell Fact. 2016, 15, 35. [Google Scholar] [CrossRef] [PubMed]
- Navarro, E.; Lorca-Pascual, J.M.; Quiles-Rosillo, M.D.; Nicolas, F.E.; Garre, V.; Torres-Martinez, S.; Ruiz-Vazquez, R.M. A negative regulator of light-inducible carotenogenesis in Mucor circinelloides. Mol. Genet. Genom. 2001, 266, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Iturriaga, E.A.; Alvarez, M.I.; Eslava, A.P.; Papp, T. Expression vectors and gene fusions for the directed modification of the carotenoid biosynthesis pathway in Mucor circinelloides. Methods Mol. Biol. 2018, 1852, 239–256. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, X.; Tan, T. Lipid and carotenoid production by Rhodotorula glutinis under irradiation/high-temperature and dark/low-temperature cultivation. Bioresour. Technol. 2014, 157, 149–153. [Google Scholar] [CrossRef]
- Tkáčová, J.; Čaplová, J.; Klempová, T.; Čertík, M. Correlation between lipid and carotenoid synthesis in torularhodin-producing Rhodotorula glutinis. Ann. Microbiol. 2017, 67, 541–551. [Google Scholar] [CrossRef]
- Somashekar, D.; Joseph, R. Inverse relationship between carotenoid and lipid formation in Rhodotorula gracilis according to the C/N ratio of the growth medium. World J. Microbiol. Biotechnol. 2000, 16, 491–493. [Google Scholar] [CrossRef]
- Zhekisheva, M.; Boussiba, S.; Khozin-Goldberg, I.; Zarka, A.; Cohen, Z. Accumulation of oleic acid in Haematococcus pluvialis (Chlorophyceae) under nitrogen starvation or high light is correlated with that of astaxanthin esters. J. Phycol. 2002, 38, 325–331. [Google Scholar] [CrossRef]
- Minhas, A.K.; Hodgson, P.; Barrow, C.J.; Adholeya, A. A Review on the Assessment of Stress Conditions for Simultaneous Production of Microalgal Lipids and Carotenoids. Front. Microbiol. 2016, 7, 546. [Google Scholar] [CrossRef] [PubMed]
- Velayos, A.; Eslava, A.P.; Iturriaga, E.A. A bifunctional enzyme with lycopene cyclase and phytoene synthase activities is encoded by the carRP gene of Mucor circinelloides. Eur. J. Biochem. 2000, 267, 5509–5519. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.; Navarro, E.; Lopez-Garcia, S.; Chen, Y.Q.; Zhang, H.; Chen, W.; Garre, V. Generation of lycopene-overproducing strains of the fungus Mucor circinelloides reveals important aspects of lycopene formation and accumulation. Biotechnol. Lett. 2017, 39, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Vongsangnak, W.; Kingkaw, A.; Yang, J.; Song, Y.; Laoteng, K. Dissecting metabolic behavior of lipid over-producing strain of Mucor circinelloides through genome-scale metabolic network and multi-level data integration. Gene 2018, 670, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Xin, F.; Wang, R.; Chang, Y.; Gao, M.; Xie, Z.; Yang, W.; Chen, M.; Zhang, H.; Song, Y. Homologous overexpression of diacylglycerol acyltransferase in oleaginous fungus Mucor circinelloides WJ11 enhances lipid accumulation under static solid cultivation. J. Agric. Food Chem. 2022, 70, 9073–9083. [Google Scholar] [CrossRef]
- Kendrick, A.; Ratledge, C. Desaturation of polyunsaturated fatty acids in Mucor circinelloides and the involvement of a novel membrane-bound malic enzyme. Eur. J. Biochem. 1992, 209, 667–673. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A Simple Method for the Isolation and Purification of Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Kroll, K.W.; Mokaram, N.E.; Pelletier, A.R.; Frankhouser, D.E.; Westphal, M.S.; Stump, P.A.; Stump, C.L.; Bundschuh, R.; Blachly, J.S.; Yan, P. Quality control for RNA-Seq (QuaCRS): An integrated quality control pipeline. Cancer Inform. 2014, 13, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, Y.; Shi, C.; Huang, Z.; Zhang, Y.; Li, S.; Li, Y.; Ye, J.; Yu, C.; Li, Z.; et al. SOAPnuke: A MapReduce acceleration-supported software for integrated quality control and preprocessing of high-throughput sequencing data. Gigascience 2018, 7, gix120. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Hermoso, D.; Criscuolo, A.; Lee, S.C.; Legrand, M.; Chaouat, M.; Denis, B.; Lafaurie, M.; Rouveau, M.; Soler, C.; Schaal, J.V.; et al. Outbreak of invasive wound mucormycosis in a burn unit due to multiple strains of Mucor circinelloides f. circinelloides resolved by whole-genome sequencing. mBio 2018, 9, e00573-18. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Billmyre, R.B.; Li, A.; Carson, S.; Sykes, S.M.; Huh, E.Y.; Mieczkowski, P.; Ko, D.C.; Cuomo, C.A.; Heitman, J. Analysis of a food-borne fungal pathogen outbreak: Virulence and genome of a Mucor circinelloides isolate from yogurt. mBio 2014, 5, e01390-14. [Google Scholar] [CrossRef] [PubMed]
- Findley, K.; Oh, J.; Yang, J.; Conlan, S.; Deming, C.; Meyer, J.A.; Schoenfeld, D.; Nomicos, E.; Park, M.; Program, N.I.H.I.S.C.C.S.; et al. Topographic diversity of fungal and bacterial communities in human skin. Nature 2013, 498, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M. Enzyme Annotation and Metabolic Reconstruction Using KEGG. Methods Mol. Biol. 2017, 1611, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating viruses and cellular organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef] [PubMed]
- Consortium, T.G.O. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar] [CrossRef]
- Pruitt, K.D.; Tatusova, T.; Maglott, D.R. NCBI reference sequences (RefSeq): A curated non-redundant sequence database of genomes, transcripts and proteins. Nucleic Acids Res. 2007, 35, D61–D65. [Google Scholar] [CrossRef]
- Patil, K.R.; Nielsen, J. Uncovering transcriptional regulation of metabolism by using metabolic network topology. Proc. Natl. Acad. Sci. USA 2005, 102, 2685–2689. [Google Scholar] [CrossRef] [PubMed]
- Varemo, L.; Nielsen, J.; Nookaew, I. Enriching the gene set analysis of genome-wide data by incorporating directionality of gene expression and combining statistical hypotheses and methods. Nucleic Acids Res. 2013, 41, 4378–4391. [Google Scholar] [CrossRef] [PubMed]
- Laoteng, K.; Jitsue, S.; Dandusitapunth, Y.; Cheevadhanarak, S. Ethanol-induced changes in expression profiles of cell growth, fatty acid and desaturase genes of Mucor rouxii. Fungal Genet. Biol. 2008, 45, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Iturriaga, E.A.; Velayos, A.; Eslava, A.P. Structure and function of the genes involved in the biosynthesis of carotenoids in the Mucorales. Biotechnol. Bioprocess. Eng. 2000, 5, 263–274. [Google Scholar] [CrossRef]
- Stajich, J.E. Fungal Evolution: Mucor and Phycomyces See Double. Curr. Biol. 2016, 26, R775–R777. [Google Scholar] [CrossRef] [PubMed]
- Thananusak, R.; Laoteng, K.; Raethong, N.; Koffas, M.; Vongsangnak, W. Dissecting Metabolic Regulation in Mycelial Growth and Fruiting Body Developmental Stages of Cordyceps militaris through Integrative Transcriptome Analysis. Biotechnol. Bioprocess. Eng. 2023, 28, 406–418. [Google Scholar] [CrossRef]
- Harris, M.A.; Clark, J.; Ireland, A.; Lomax, J.; Ashburner, M.; Foulger, R.; Eilbeck, K.; Lewis, S.; Marshall, B.; Mungall, C.; et al. The Gene Ontology (GO) database and informatics resource. Nucleic Acids Res. 2004, 32, D258–D261. [Google Scholar] [CrossRef] [PubMed]
- Aleksander, S.A.; Balhoff, J.; Carbon, S.; Cherry, J.M.; Drabkin, H.J.; Ebert, D.; Feuermann, M.; Gaudet, P.; Harris, N.L.; Hill, D.P.; et al. The Gene Ontology knowledgebase in 2023. Genetics 2023, 224, iyad031. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [PubMed]
- Bueno, E.; Sit, B.; Waldor, M.K.; Cava, F. Anaerobic nitrate reduction divergently governs population expansion of the enteropathogen Vibrio cholerae. Nat. Microbiol. 2018, 3, 1346–1353. [Google Scholar] [CrossRef]
- Marmagne, A.; Jasinski, S.; Fagard, M.; Bill, L.; Guerche, P.; Masclaux-Daubresse, C.; Chardon, F. Post-flowering biotic and abiotic stresses impact nitrogen use efficiency and seed filling in Arabidopsis thaliana. J. Exp. Bot. 2020, 71, 4578–4590. [Google Scholar] [CrossRef] [PubMed]
- Harding, C.O.; Winn, S.R.; Gibson, K.M.; Arning, E.; Bottiglieri, T.; Grompe, M. Pharmacologic inhibition of L-tyrosine degradation ameliorates cerebral dopamine deficiency in murine phenylketonuria (PKU). J. Inherit. Metab. Dis. 2014, 37, 735–743. [Google Scholar] [CrossRef]
- Panda, S. Circadian physiology of metabolism. Science 2016, 354, 1008–1015. [Google Scholar] [CrossRef]
- Ruenwai, R.; Cheevadhanarak, S.; Laoteng, K. Overexpression of acetyl-CoA carboxylase gene of Mucor rouxii enhanced fatty acid content in Hansenula polymorpha. Mol. Biotechnol. 2009, 42, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Tai, M.; Stephanopoulos, G. Engineering the push and pull of lipid biosynthesis in oleaginous yeast Yarrowia lipolytica for biofuel production. Metab. Eng. 2013, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.T.; Tu, B.P.; Tang, Y. Eight Kinetically Stable but Thermodynamically Activated Molecules that Power Cell Metabolism. Chem. Rev. 2018, 118, 1460–1494. [Google Scholar] [CrossRef] [PubMed]
- Luengo, A.; Gui, D.Y.; Vander Heiden, M.G. Targeting Metabolism for Cancer Therapy. Cell Chem. Biol. 2017, 24, 1161–1180. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, J.B.; Haigis, M.C. The multifaceted contributions of mitochondria to cellular metabolism. Nat. Cell Biol. 2018, 20, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Chalishazar, M.D.; Wait, S.J.; Huang, F.; Ireland, A.S.; Mukhopadhyay, A.; Lee, Y.; Schuman, S.S.; Guthrie, M.R.; Berrett, K.C.; Vahrenkamp, J.M.; et al. MYC-Driven Small-Cell Lung Cancer is Metabolically Distinct and Vulnerable to Arginine Depletion. Clin. Cancer Res. 2019, 25, 5107–5121. [Google Scholar] [CrossRef] [PubMed]
- El-Hattab, A.W.; Hsu, J.W.; Emrick, L.T.; Wong, L.J.; Craigen, W.J.; Jahoor, F.; Scaglia, F. Restoration of impaired nitric oxide production in MELAS syndrome with citrulline and arginine supplementation. Mol. Genet. Metab. 2012, 105, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Bok, R.; Guerra, D.D.; Lorca, R.A.; Wennersten, S.A.; Harris, P.S.; Rauniyar, A.K.; Stabler, S.P.; MacLean, K.N.; Roede, J.R.; Brown, L.D.; et al. Cystathionine gamma-lyase promotes estrogen-stimulated uterine artery blood flow via glutathione homeostasis. Redox Biol. 2021, 40, 101827. [Google Scholar] [CrossRef] [PubMed]
- Amengual, J.; Lobo, G.P.; Golczak, M.; Li, H.N.; Klimova, T.; Hoppel, C.L.; Wyss, A.; Palczewski, K.; von Lintig, J. A mitochondrial enzyme degrades carotenoids and protects against oxidative stress. FASEB J. 2011, 25, 948–959. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenotypic Characteristics | M. circinelloides WJ11 | M. circinelloides ΔcarRP WJ11 |
|---|---|---|
| Maximum specific growth rate, µmax (h−1) | 0.765 ± 0.5 a | 0.665 ± 0.17 a |
| Biomass productivity (gDCW L−1) | 12.95 ± 0.6 a | 8.83 ± 1.03 b |
| Fatty acids in DCW (g gDCW−1, %) | 35.18 ± 2.18 a | 31.45 ± 6.36 a |
| Carotenoid content (μg gDCW−1) | 208.99 ± 7.24 a | 2.47 ± 0.4 b |
| Features | WJ11 | ΔcarRP WJ11 |
|---|---|---|
| Sequencing depth (million reads) | 43.82 | 43.82 |
| Total clean reads (million reads) | 43.18 | 43.12 |
| Sequencing quality (%) | 96.78 | 96.93 |
| Total mapped reads for genome (%) | 93.49 | 93.86 |
| Total mapped reads for gene (%) | 68.84 | 70.04 |
| Number of expressed genes | 10,063 | 10,162 |
| Total number of expressed genes | 10,287 | |
| Gene Name | Functions (EC Number) | Metabolic Pathway | Log2FC |
|---|---|---|---|
| HMPREF1544_11519 | Ornithine carbamoyltransferase (EC: 2.1.3.3) | Arginine and proline metabolism | 5.59 |
| HMPREF1544_06810 | Argininosuccinate synthase (EC: 6.3.4.5) | Arginine and proline metabolism | 3.28 |
| HMPREF1544_01650 | Argininosuccinate lyase (EC: 4.3.2.1) | Arginine and proline metabolism | 3.29 |
| HMPREF1544_11380 | Pyrroline-5-carboxylate reductase (EC: 1.5.1.2) | Arginine and proline metabolism | 1.47 |
| HMPREF1544_09829 | Acetylglutamate kinase (EC: 2.7.2.8) | Arginine and proline metabolism | 1.50 |
| HMPREF1544_09419 | Arginase (EC: 3.5.3.1) | Arginine and proline metabolism | 1.83 |
| HMPREF1544_10960 | glutamate N-acetyltransferase (EC: 2.3.1.35) | Arginine biosynthesis | 2.88 |
| HMPREF1544_01946 | Carbamoyl-phosphate synthase (EC:6.3.5.5) | Alanine, aspartate, and glutamate metabolism | 3.14 |
| HMPREF1544_07542 | Phosphoheptulonate synthase (EC:2.5.1.54) | Phenylalanine, tyrosine, and tryptophan biosynthesis | 3.26 |
| HMPREF1544_09131 | Hydroxyphenylpyruvate dioxygenase (EC:1.13.11.27) | Phenylalanine, tyrosine, and tryptophan biosynthesis | 2.11 |
| HMPREF1544_03914 | Glutaryl-CoA dehydrogenase (EC: 1.3.8.6) | Lysine degradation | 1.40 |
| HMPREF1544_01100 | Acetolactate synthase (EC:2.2.1.6) | Valine, leucine, and isoleucine metabolism | 2.09 |
| HMPREF1544_06855 | Branched-chain amino acid aminotransferase (EC:2.6.1.42) | Valine, leucine, and isoleucine metabolism | 1.27 |
| HMPREF1544_11990 | Nitrite reductase (NAD(P)H) (EC: 1.7.1.4) | Nitrogen metabolism | 4.35 |
| HMPREF1544_11989 | Nitrate reductase (NAD(P)H) (EC:1.7.1.1; 1.7.1.2; 1.7.1.3) | Nitrogen metabolism | 4.92 |
| HMPREF1544_11064 | Glutamate dehydrogenase (EC:1.4.1.2) | Nitrogen metabolism | 1.96 |
| HMPREF1544_00954 | Glutamine synthetase (EC:6.3.1.2) | Nitrogen metabolism | 1.07 |
| HMPREF1544_09200 | Fructose-bisphosphate aldolase (EC:4.1.2.13) | Glycolysis/gluconeogenesis | −1.33 |
| HMPREF1544_09493 | Pyruvate kinase (EC:2.7.1.40) | Glycolysis/gluconeogenesis | −1.01 |
| HMPREF1544_02579 | Acetyl-CoA synthetase (EC:6.2.1.1) | Pyruvate metabolism | −2.94 |
| HMPREF1544_12198 | Pyruvate decarboxylase (EC: 4.1.1.1) | Pyruvate metabolism | −2.08 |
| HMPREF1544_06544 | Isocitrate dehydrogenase (EC:1.1.1.42) | Tricarboxylic acid cycle | 1.94 |
| HMPREF1544_03574 | ATP citrate lyase (EC:2.3.3.8) | Tricarboxylic acid cycle | −2.68 |
| HMPREF1544_05482 | Fatty acid synthase (EC:2.3.1.86) | Fatty acid biosynthesis | −2.10 |
| HMPREF1544_01855 | Acetyl-CoA carboxylase (EC:6.4.1.2) | Fatty acid biosynthesis | −1.92 |
| HMPREF1544_10598 | Acetyl-CoA carboxylase (EC:6.4.1.2) | Fatty acid biosynthesis | −1.62 |
| HMPREF1544_10223 | Phytoene synthase/lycopene beta-cyclase (EC:2.5.1.32) | Carotenoid biosynthesis | −7.81 |
| Reporter Metabolite | Up-Directional p-Value |
|---|---|
| Glutamate (GLU) | 0.00009999 * |
| Phosphate (PI) | 0.00009999 * |
| Carbamoyl phosphate (CAP) | 0.00009999 * |
| NADPH | 0.00029997 * |
| Arginine succinate (ARGSUCC) | 0.00029997 * |
| Citrulline (CITR) | 0.00029997 * |
| NADP | 0.00059994 * |
| Chorismate (CHOR) | 0.0019998 * |
| ATP | 0.0022998 * |
| Glutamine (GLN) | 0.0031997 * |
| Diphosphate (PPI) | 0.0033997 * |
| Ornithine (ORN) | 0.0033997 * |
| Arginine (ARG) | 0.0037996 * |
| Fumarate (FUM) | 0.0062994 * |
| HCO3 | 0.0062994 * |
| α-Ketoglutarate (AKG) | 0.0074993 * |
| 5-Phospho-alpha-D-ribose 1-diphosphate (PRPP) | 0.009799 * |
| CO2 | 0.012199 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Thananusak, R.; Raethong, N.; Yang, J.; Wei, M.; Zhao, X.; Laoteng, K.; Song, Y.; Vongsangnak, W. Dissecting Holistic Metabolic Acclimatization of Mucor circinelloides WJ11 Defective in Carotenoid Biosynthesis. Biology 2024, 13, 276. https://doi.org/10.3390/biology13040276
Li F, Thananusak R, Raethong N, Yang J, Wei M, Zhao X, Laoteng K, Song Y, Vongsangnak W. Dissecting Holistic Metabolic Acclimatization of Mucor circinelloides WJ11 Defective in Carotenoid Biosynthesis. Biology. 2024; 13(4):276. https://doi.org/10.3390/biology13040276
Chicago/Turabian StyleLi, Fanyue, Roypim Thananusak, Nachon Raethong, Junhuan Yang, Mingyue Wei, Xingtang Zhao, Kobkul Laoteng, Yuanda Song, and Wanwipa Vongsangnak. 2024. "Dissecting Holistic Metabolic Acclimatization of Mucor circinelloides WJ11 Defective in Carotenoid Biosynthesis" Biology 13, no. 4: 276. https://doi.org/10.3390/biology13040276