

Different Approaches of Forest Type Classifications for Argentina Based on Functional Forests and Canopy Cover Composition by Tree Species

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
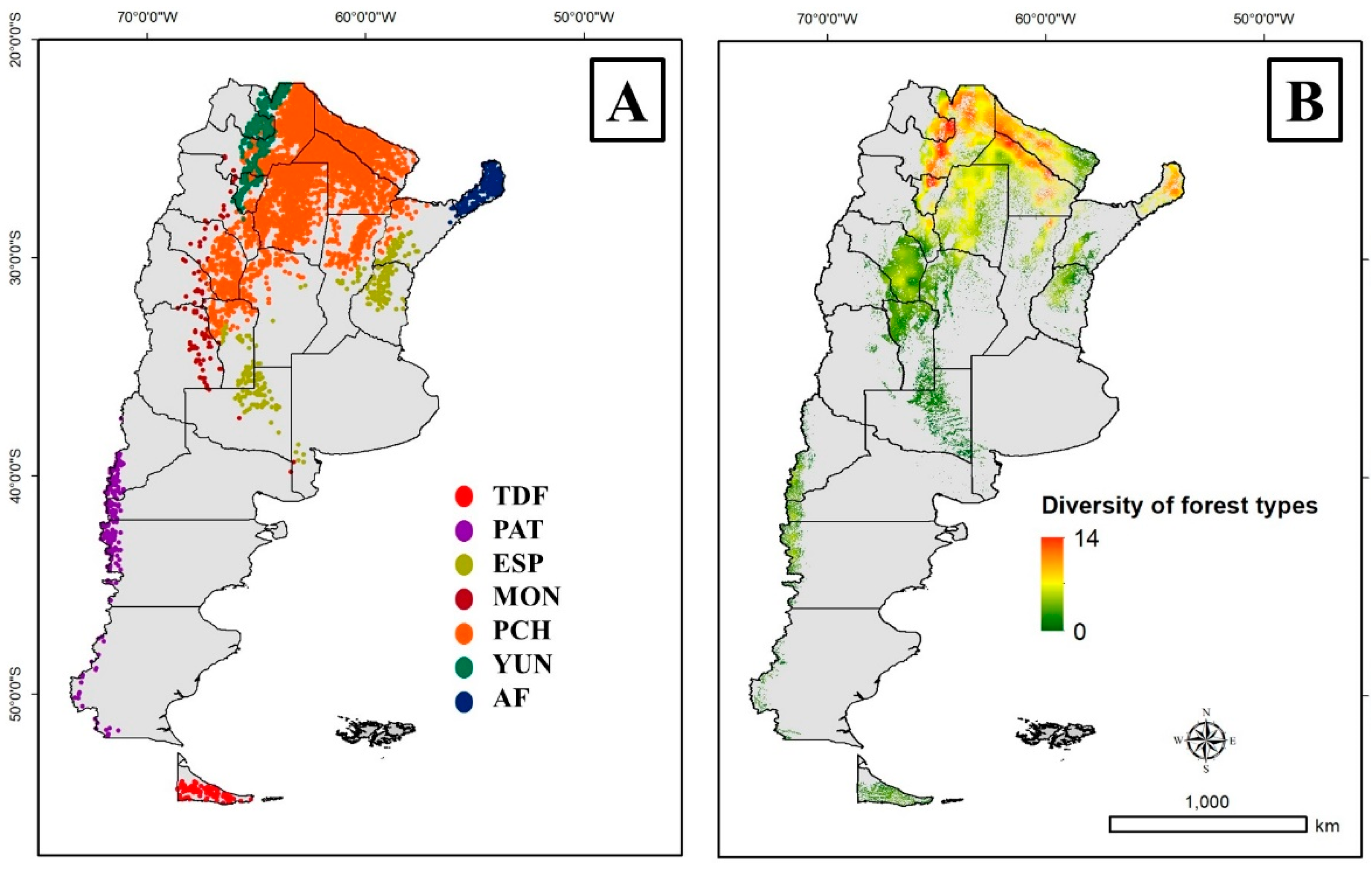
2.1. Study Area
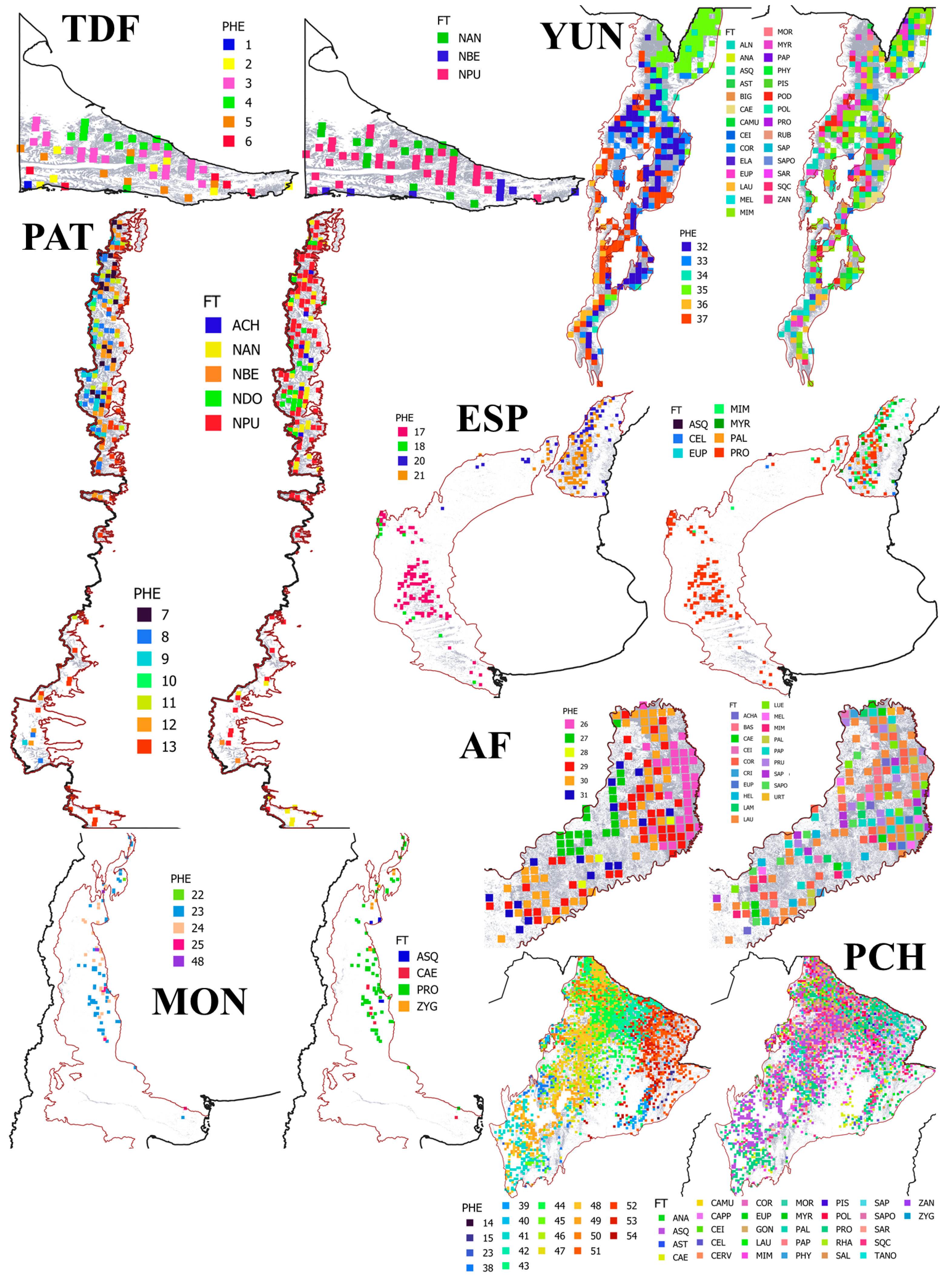
2.2. Forest Type Classification Based on Phenoclusters
2.3. Forest Type Classification Based on Forest Canopy Cover Composition by Tree Species
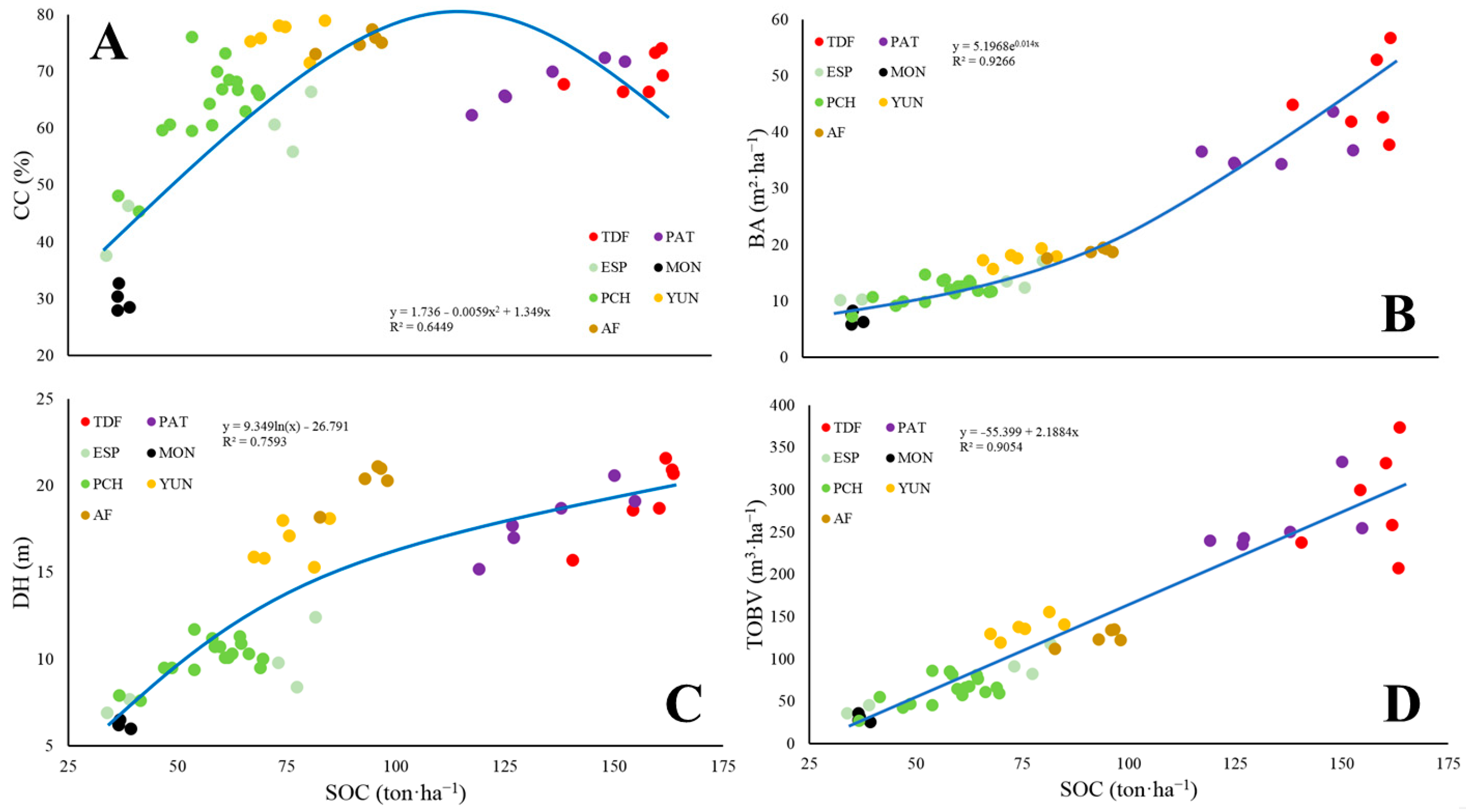
2.4. Statistical Analyses
3. Results
3.1. Forest Type Classification Based on Phenoclusters
3.2. Forest Type Classification Based on Forest Canopy Cover Composition by Tree Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martínez Pastur, G.; Schlichter, T.; Matteucci, S.D.; Gowda, J.; Huertas Herrera, A.; Toro Manríquez, M.; Lencinas, M.V.; Cellini, J.M.; Peri, P.L. Synergies and trade-offs of national conservation policy and agro-forestry management over forest loss in Argentina during the last decade. In Latin America in Times of Global Environmental Change; Lorenzo, C., Ed.; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Koff, H.; Challenger, A.; Portillo, I. Guidelines for operationalizing policy coherence for development (PCD) as a methodology for the design and implementation of sustainable development strategies. Sustainability 2020, 2, 4055. [Google Scholar] [CrossRef]
- Xu, H.; Cao, Y.; Yu, D.; Cao, M.; He, Y.; Gill, M.; Pereira, H. Ensuring effective implementation of the post-2020 global biodiversity targets. Nat. Ecol. Evol. 2021, 5, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Angelstam, P.; Albulescu, C.; Andrianambinina, O.; Aszalós, R.; Borovichev, E.; Cano Cardona, W.; Dobrynin, D.; Fedoriak, M.; Firm, D.; Hunter, M.; et al. Frontiers of protected areas versus forest exploitation: Assessing habitat network functionality in 16 case study regions globally. Ambio 2021, 50, 2286–2310. [Google Scholar] [CrossRef] [PubMed]
- Allan, J.R.; Possingham, H.; Atkinson, S.; Waldron, A.; di Marco, M.; Butchart, S.; Adams, V.; Kissling, W.; Worsdell, T.; Sandbrook, C.; et al. The minimum land area requiring conservation attention to safeguard biodiversity. Science 2022, 376, 1094–1101. [Google Scholar] [CrossRef] [PubMed]
- Peri, P.L.; Martínez Pastur, G.; Schlichter, T. Uso Sustentable del Bosque: Aportes Desde la Silvicultura Argentina; Ministerio de Ambiente y Desarrollo Sostenible de la Nación Argentina: Buenos Aires, Argentina, 2021. [Google Scholar]
- Martinuzzi, S.; Radeloff, V.C.; Martínez Pastur, G.; Rosas, Y.M.; Lizarraga, L.; Politi, N.; Rivera, L.; Huertas Herrera, A.; Silveira, E.M.O.; Olah, A.; et al. Informing forest conservation planning with detailed human footprint data for Argentina. Glob. Ecol. Conserv. 2021, 31, e01787. [Google Scholar] [CrossRef]
- Zhu, L.; Hughes, A.; Zhao, X.; Zhou, L.; Ma, K.; Shen, X.; Li, S.; Liu, M.; Xu, W.; Watson, J. Regional scalable priorities for national biodiversity and carbon conservation planning in Asia. Sci. Adv. 2021, 7, eabe4261. [Google Scholar] [CrossRef] [PubMed]
- Keith, D.A.; Ferrer-Paris, J.R.; Nicholson, E.; Bishop, M.; Polidoro, B.; Ramirez-Llodra, E.; Tozer, M.; Nel, J.; Mac Nally, R.; Gregr, E.; et al. A function-based typology for Earth’s ecosystems. Nature 2022, 610, 513–518. [Google Scholar] [CrossRef]
- Crausbay, S.D.; Sofaer, H.; Cravens, A.; Chaffin, B.; Clifford, K.; Gross, J.; Knapp, C.; Lawrence, D.; Magness, D.; Miller-Rushing, A.; et al. A science agenda to inform natural resource management decisions in an era of ecological transformation. BioScience 2022, 72, 71–90. [Google Scholar] [CrossRef]
- Jansson, G.; Angelstam, P. Threshold levels of habitat composition for the presence of the long-tailed tit (Aegithalos caudatus) in a boreal landscape. Land. Ecol. 1999, 14, 283–290. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, D. Accurate mapping of forest types using dense seasonal Landsat time-series. ISPRS J. Photogram Rem. Sen. 2014, 96, 1–11. [Google Scholar] [CrossRef]
- López, V.L.; Herrera, A.H.; Rosas, Y.M.; Cellini, J.K. Optimal environmental drivers of high-mountains forest: Polylepis tarapacana cover evaluation in their southernmost distribution range of the Andes. Trees For. People 2022, 9, e100321. [Google Scholar] [CrossRef]
- Silveira, E.M.O.; Radeloff, V.C.; Martínez Pastur, G.; Martinuzzi, S.; Politi, N.; Lizarraga, L.; Rivera, L.; Gavier Pizarro, G.; Yin, H.; Rosas, Y.M.; et al. Forest phenoclusters for Argentina based on vegetation phenology and climate. Ecol. Appl. 2022, 32, e2526. [Google Scholar] [CrossRef]
- Liu, X.; Frey, J.; Munteanu, C.; Still, N.; Koch, B. Mapping tree species diversity in temperate montane forests using Sentinel-1 and Sentinel-2 imagery and topography data. Rem. Sen. Environ. 2023, 292, e113576. [Google Scholar] [CrossRef]
- Wadoux, A.; Minasny, B.; McBratney, A. Machine learning for digital soil mapping: Applications, challenges and suggested solutions. Earth-Sci. Rev. 2020, 210, e103359. [Google Scholar] [CrossRef]
- Harris, N.L.; Gibbs, D.; Baccini, A.; Birdsey, R.; de Bruin, S.; Farina, M.; Fatoyinbo, L.; Hansen, M.; Herold, M.; Houghton, R.; et al. Global maps of twenty-first century forest carbon fluxes. Nat. Clim. Chang. 2021, 11, 234–240. [Google Scholar] [CrossRef]
- Portillo-Quintero, C.; Hernández-Stefanoni, J.L.; Reyes-Palomeque, G.; Subedi, M. Novel approaches in tropical forests mapping and monitoring-time for operationalization. Rem. Sen. 2022, 14, 5068. [Google Scholar] [CrossRef]
- Eva, H.D.; Belward, A.; de Miranda, E.; di Bella, C.; Gond, V.; Huber, O.; Jones, S.; Sgrenzaroli, M.; Fritz, S. A land cover map of South America. Glob. Chang. Biol. 2004, 10, 731–744. [Google Scholar] [CrossRef]
- Stanimirova, R.; Graesser, J.; Olofsson, P.; Friedl, M.A. Widespread changes in 21st century vegetation cover in Argentina, Paraguay, and Uruguay. Rem. Sen. Environ. 2022, 282, e113277. [Google Scholar] [CrossRef]
- Silveira, E.M.O.; Radeloff, V.C.; Martinuzzi, S.; Martínez Pastur, G.; Bono, J.; Politi, N.; Lizarraga, L.; Rivera, L.; Ciuffoli, L.; Rosas, Y.M.; et al. Nationwide native forest structure maps for Argentina based on forest inventory data, SAR Sentinel-1 and vegetation metrics from Sentinel-2 imagery. Rem. Sen. Environ. 2023, 285, e113391. [Google Scholar] [CrossRef]
- Martinuzzi, S.; Olah, A.M.; Rivera, L.; Politi, N.; Silveira, E.M.O.; Martínez Pastur, G.; Rosas, Y.M.; Lizarraga, L.; Nazaro, P.; Bardavid, S.; et al. Closing the research-implementation gap: Integrating species and human footprint data into Argentina’s forest planning. Biol. Conserv. 2023, 286, e110257. [Google Scholar] [CrossRef]
- Peri, P.L.; Gaitán, J.; Mastrangelo, M.; Nosetto, M.; Villagra, P.; Balducci, E.; Pinazo, M.; Eclesia, R.; von Wallis, A.; Villarino, S.; et al. Soil organic carbon stocks in native forest of Argentina: A useful surrogate for mitigation and conservation planning under climate variability. Ecol. Proc. 2024, 13, e1. [Google Scholar] [CrossRef]
- Silveira, E.M.O.; Radeloff, V.C.; Martinuzzi, S.; Martínez Pastur, G.; Rivera, L.; Politi, N.; Lizarraga, L.; Farwell, L.S.; Elsen, P.R.; Pidgeon, A.M. Spatio-temporal remotely sensed indices identify hotspots of biodiversity conservation concern. Rem. Sen. Environ. 2021, 258, e112368. [Google Scholar] [CrossRef]
- MAyDS. Primer Inventario Nacional de Bosques Nativos, Informe Nacional; Ministerio de Ambiente y Desarrollo Sostenible: Buenos Aires, Argentina, 2005.
- SGAyDS. Segundo Inventario Nacional de Bosques Nativos: Manual de Campo; Secretaría de Gobierno de Ambiente y Desarrollo Sustentable de la Nación: Buenos Aires, Argentina, 2019. [Google Scholar]
- Matteucci, S.D.; Martinez Pastur, G.; Lencinas, M.V.; Rovere, A.; Amoroso, M.M.; Barberis, I.; Vesprini, J.L.; Galetto, L.; Torres, C.; Villagra, P.E.; et al. Breve descripción de las regiones forestales de la Argentina. In Uso Sustentable del Bosque: Aportes Desde la Silvicultura Argentina; Peri, P.L., Martínez Pastur, G., Schlichter, T., Eds.; Ministerio de Ambiente y Desarrollo Sostenible de la Nación Argentina: Buenos Aires, Argentina, 2021. [Google Scholar]
- Burkart, R.; Bárbaro, N.O.; Sánchez, R.O.; Gómez, D.A. Eco-Regiones de la Argentina; Secretaría de Recursos Naturales y Desarrollo Sustentable, Administración de Parque Nacionales, Gobierno de la República Argentina: Buenos Aires, Argentina, 1999. [Google Scholar]
- Morello, J.; Matteucci, S.; Rodríguez, A.; Silva, M. Ecorregiones y Complejos Ecosistémicos Argentinos; Orientación Gráfica Editora: Buenos Aires, Argentina, 2012. [Google Scholar]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-resolution global maps of 21st-Century forest cover change. Science 2013, 80, 850–853. [Google Scholar] [CrossRef]
- Potapov, P.; Li, X.; Hernandez-Serna, A.; Tyukavina, A.; Hansen, M.C.; Kommareddy, A.; Pickens, A.; Turubanova, S.; Tang, H.; Edibaldo Silva, C.; et al. Mapping global forest canopy height through integration of GEDI and LANDSAT data. Rem. Sen. Environ. 2021, 253, e112165. [Google Scholar] [CrossRef]
- Wilson, K.A.; McBride, M.F.; Bode, M.; Possingham, H.P. Prioritizing global conservation efforts. Nature 2006, 440, 337–340. [Google Scholar] [CrossRef]
- Hmielowski, T.L.; Carter, S.K.; Spaul, H.; Helmers, D.P.; Radeloff, V.C.; Zedler, P.H. Prioritizing land management efforts at a landscape scale: A case study using prescribed fire in Wisconsin. Ecol. Appl. 2015, 26, 1018–1029. [Google Scholar] [CrossRef]
- Potapov, P.; Hansen, M.; Pickens, A.; Hernandez-Serna, A.; Tyukavina, A.; Turubanova, S.; Zalles, V.; Li, X.; Khan, A.; Stolle, F.; et al. The global 2000–2020 land cover and land use change dataset derived from the Landsat archive: First results. Front. Rem. Sen. 2022, 13, e856903. [Google Scholar] [CrossRef]
- Cabrera, A.L. Fitogeografía de la República Argentina. Bol. Soc. Arg. Bot. 1971, 14, 1–2. [Google Scholar]
- Cabrera, A.L. Regiones fitogeográficas argentinas. In Enciclopedia Argentina de Agricultura y Jardinería; Kugler, W.F., Ed.; ACME: Buenos Aires, Argentina, 1976; Volume 2, pp. 1–85. [Google Scholar]
- Cabrera, A.L. Regiones Fitogeográficas Argentinas; ACME: Buenos Aires, Argentina, 1994. [Google Scholar]
- Cabrera, A.L.; Willink, A. Biogeografía de América Latina, Monografía 13, Serie de Biología; Secretaría General de la Organización de los Estados Americanos: Washington, DC, USA, 1973. [Google Scholar]
- Oyarzabal, M.; Clavijo, J.; Oakley, L.; Biganzoli, F.; Tognetti, P.; Barberis, I.; Maturo, H.M.; Aragón, R.; Campanello, P.; Prado, D.; et al. Unidades de vegetación de la Argentina. Ecol. Aust. 2018, 28, 40–63. [Google Scholar] [CrossRef]
- Derguy, M.R.; Frangi, J.L.; Martinuzzi, S. Las regiones forestales de la Argentina en el contexto de zonas de vida de Holdridge. In Uso Sustentable del Bosque: Aportes Desde la Silvicultura Argentina; Peri, P.L., Martínez Pastur, G., Schlichter, T., Eds.; Ministerio de Ambiente y Desarrollo Sostenible de la Nación Argentina: Buenos Aires, Argentina, 2021. [Google Scholar]
- Martínez Pastur, G.; Amoroso, M.M.; Baldi, G.; Barrera, M.D.; Brown, A.D.; Chauchard, L.; Galetto, L.; Garibaldi, L.A.; Gasparri, I.; Kees, S.; et al. ¿Qué es un bosque nativo en Argentina?: Marco conceptual para una correcta definición de acuerdo a las políticas institucionales nacionales y al conocimiento científico disponible. Ecol. Aust. 2023, 33, 152–169. [Google Scholar] [CrossRef]
- Fomin, V.; Mikhailovich, A. Russian approaches to the forest type classification. IOP Conf. Ser. Earth Environ. Sci. 2021, 906, e012023. [Google Scholar] [CrossRef]
- Prodan, M.; Peters, R.; Cox, F.; Real, P. Mensura Forestal; GTZ-IICA: San José, Costa Rica, 1997. [Google Scholar]
- Criteria and Indicators for the Conservation and Sustainable Management of Temperate and Boreal Forests; The Montreal Process: Montreal, QC, Canada, 1998.
- Cajander, A.K. Forest types and their significance. Acta For. Fennica 1949, 56, 1–71. [Google Scholar] [CrossRef]
- Blasi, C.; Carranza, M.L.; Frondoni, R.; Rosati, L. Ecosystem classification and mapping: A proposal for Italian landscapes. Appl. Veg. Sci. 2000, 3, 233–242. [Google Scholar] [CrossRef]
- Larsen, J.B.; Nielsen, A.B. Nature-based forest management: Where are we going? Elaborating forest development types in and with practice. For. Ecol. Manag. 2006, 238, 107–117. [Google Scholar] [CrossRef]
- Barbati, A.; Corona, P.; Marchetti, M. A forest typology for monitoring sustainable forest management: The case of European forest types. Plant Biosyst. 2007, 141, 93–103. [Google Scholar] [CrossRef]
- Huertas Herrera, A.; Toro Manríquez, M.D.R.; Salinas, J.; Rivas Guíñez, F.; Lencinas, M.V.; Martínez Pastur, G. Relationships among livestock, structure, and regeneration in Chilean Austral Macrozone temperate forests. Trees For. People 2023, 13, e100426. [Google Scholar] [CrossRef]
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The shuttle radar topography mission. Rev. Geoph. 2007, 45, RG2004. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climat. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Battersby, S.E.; Strebe, D.D.; Finn, M.P. Shapes on a plane: Evaluating the impact of projection distortion on spatial binning. Cart. Geograp. Inf. Sci. 2017, 44, 410–421. [Google Scholar] [CrossRef]
- Rosas, Y.M.; Peri, P.L.; Benítez, J.; Lencinas, M.V.; Politi, N.; Martínez Pastur, G. Potential biodiversity map of bird species (Passeriformes): Analyses of ecological niche, environmental characterization and identification of priority conservation areas in southern Patagonia. J. Nat. Conserv. 2023, 73, e126413. [Google Scholar] [CrossRef]
- Dirección Nacional de Bosques. Datos del Segundo Inventario Nacional de Bosques Nativos de la República Argentina; Ministerio de Ambiente y Desarrollo Sostenible de la Nación: Buenos Aires, Argentina, 2021. [Google Scholar]
- Zuloaga, F.O.; Belgrano, M.J.; Zanotti, C.A. Actualización del catálogo de las plantas vasculares del cono sur. Darwiniana 2019, 7, 208–278. [Google Scholar] [CrossRef]
- Jones, S.L. Territory size in mixed-grass prairie songbirds. Can. Field Nat. 2011, 125, 5–12. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, X.; Hernandez-Clemente, R.; Lu, W.; Manzanedo, R.D. Global forest types based on climatic and vegetation data. Sustainability 2022, 14, 634. [Google Scholar] [CrossRef]
- Schimper, A.F.W. Plant geography upon a physiological basis. Bot. Gaz. 1904, 37, 392–393. [Google Scholar]
- Costanza, J.K.; Faber-Langendoen, D.; Coulston, J.W.; Wear, D.N. Classifying forest inventory data into species-based forest community types at broad extents: Exploring tradeoffs among supervised and unsupervised approaches. For. Ecosyst. 2018, 5, e8. [Google Scholar] [CrossRef]
- Ersoy Mirici, M.; Satir, O.; Berberoglu, S. Monitoring the Mediterranean type forests and land-use/cover changes using appropriate landscape metrics and hybrid classification approach in Eastern Mediterranean of Turkey. Environ. Earth Sci. 2020, 79, e492. [Google Scholar] [CrossRef]
- Box, E.O. Predicting physiognomic vegetation types with climate variables. Vegetatio 1981, 45, 127–139. [Google Scholar] [CrossRef]
- Holdridge, L.R. Determination of world plant formations from simple climatic data. Science 1947, 105, 367–368. [Google Scholar] [CrossRef]
- DeFries, R.S.; Hansen, M.C.; Townshend, J.R.G.; Janetos, A.C.; Loveland, T.R. A new global 1-km dataset of percentage tree cover derived from remote sensing. Glob. Chang. Biol. 2000, 6, 247–254. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, S.; Yan, X.; Chen, Z. A global classification of vegetation based on NDVI, rainfall and temperature. Int. J. Clim. 2017, 37, 2318–2324. [Google Scholar] [CrossRef]
- Ju, Y.; Bohrer, G. Classification of wetland vegetation based on NDVI time series from the HLS dataset. Rem. Sen. 2022, 14, 2107. [Google Scholar] [CrossRef]
- Beck, P.S.; Juday, G.P.; Alix, C.; Barber, V.A.; Winslow, S.E.; Sousa, E.E.; Heiser, P.; Herriges, J.D.; Goetz, S. Changes in forest productivity across Alaska consistent with biome shift. Ecol. Let. 2011, 14, 373–379. [Google Scholar] [CrossRef]
- Metzger, M.J.; Bunce, R.G.H.; Jongman, R.H.G.; Sayre, R.; Trabucco, A.; Zomer, R. A high-resolution bioclimate map of the world: A unifying framework for global biodiversity research and monitoring. Glob. Ecol. Biogeogr. 2013, 22, 630–638. [Google Scholar] [CrossRef]
- Fomin, V.; Mikhailovich, A.; Zalesov, S.; Popov, A.; Terekhov, G. Development of ideas within the framework of the genetic approach to the classification of forest types. Balt. For. 2021, 27, e466. [Google Scholar] [CrossRef]
- Ivanova, N.; Fomin, V.; Kusbach, A. Experience of forest ecological classification in assessment of vegetation dynamics. Sustainability 2022, 14, 3384. [Google Scholar] [CrossRef]
- Fang, F.; McNeil, B.E.; Warner, T.A.; Maxwell, A.E.; Dahle, G.A.; Eutsler, E.; Li, J. Discriminating tree species at different taxonomic levels using multi-temporal WorldView-3 imagery in Washington DC, USA. Rem. Sen. Environ. 2020, 246, e111811. [Google Scholar] [CrossRef]
- Zhang, B.; Zhao, L.; Zhang, X. Three-dimensional convolutional neural network model for tree species classification using airborne hyperspectral images. Rem. Sen. Environ. 2020, 247, e111938. [Google Scholar] [CrossRef]
- Erinjery, J.J.; Singh, M.; Kent, R. mapping and assessment of vegetation types in the tropical rainforests of the western Ghats using multispectral Sentinel-2 and SAR Sentinel-1 satellite imagery. Rem. Sen. Environ. 2018, 216, 345–354. [Google Scholar] [CrossRef]
- Peri, P.L.; Lasagno, R.; Martínez Pastur, G.; Atkinson, R.; Thomas, E.; Ladd, B. Soil carbon is a useful surrogate for conservation planning in developing nations. Sci. Rep. 2019, 9, 3905. [Google Scholar] [CrossRef]
- Canedoli, C.; Ferrè, C.; Abu El Khair, D.; Comolli, R.; Liga, C.; Mazzucchelli, F.; Proietto, A.; Rota, N.; Colombo, G.; Bassano, B.; et al. Evaluation of ecosystem services in a protected mountain area: Soil organic carbon stock and biodiversity in alpine forests and grasslands. Ecosyst. Ser. 2020, 44, e101135. [Google Scholar] [CrossRef]
- Edmonds, R.L.; Chappell, H.N. Relationships between soil organic matter and forest productivity in western Oregon and Washington. Can. J. For. Res. 1994, 24, 1101–1106. [Google Scholar] [CrossRef]
- Grigal, D.F.; Vance, E.D. Influence of soil organic matter on forest productivity. N. Z. J. For. Sci. 2000, 30, 169–205. [Google Scholar]
- Hoagland, S.J.; Beier, P.; Lee, D. Using MODIS NDVI phenoclasses and phenoclusters to characterize wildlife habitat: Mexican spotted owl as a case study. For. Ecol. Manag. 2018, 412, 80–93. [Google Scholar] [CrossRef]
- Bajocco, S.; Ferrara, C.; Alivernini, A.; Bascietto, M.; Ricotta, C. Remotely-sensed phenology of Italian forests: Going beyond the species. Int. J. Appl. Earth Obs. Geoinf. 2019, 74, 314–321. [Google Scholar] [CrossRef]
- Barrera, M.D.; Frangi, J.L.; Richter, L.; Perdomo, M.; Pinedo, L. Structural and functional changes in Nothofagus pumilio forests along an altitudinal gradient in Tierra del Fuego, Argentina. J. Veg. Sci. 2000, 11, 179–188. [Google Scholar] [CrossRef]
- Matskovsky, V.; Roig, F.A.; Fuentes, M.; Korneva, I.; Araneo, D.; Linderholm, H.; Aravena, J.C. Summer temperature changes in Tierra del Fuego since AD 1765: Atmospheric drivers and tree-ring reconstruction from the southernmost forests of the world. Clim. Dyn. 2023, 60, 1635–1649. [Google Scholar] [CrossRef]
- Mathiasen, P.; Premoli, A.C. Out in the cold: Genetic variation of Nothofagus pumilio (Nothofagaceae) provides evidence for latitudinally distinct evolutionary histories in austral South America. Mol. Ecol. 2010, 19, 371–385. [Google Scholar] [CrossRef] [PubMed]
- Soliani, C.; Marchelli, P.; Mondino, V.; Pastorino, M.; Mattera, G.; Gallo, L.; Aparicio, A.; Torres, A.D.; Tejera, L.; Schinelli Casares, T. Nothofagus pumilio and N. antarctica: The most widely distributed and cold-tolerant southern beeches in Patagonia. In Low Intensity Breeding of Native Forest Trees in Argentina; Pastorino, M.J., Marchelli, P., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Schiefer, F.; Kattenborn, T.; Frick, A.; Frey, J.; Schall, P.; Koch, B.; Schmidtlein, S. Mapping forest tree species in high resolution UAV-based RGB-imagery by means of convolutional neural networks. ISPRS J. Photogram. Rem. Sen. 2020, 170, 205–215. [Google Scholar] [CrossRef]
- Ruefenacht, B.; Finco, M.V.; Nelson, M.D.; Czaplewski, R.; Helmer, E.H.; Blackard, J.A.; Holden, G.R.; Lister, A.J.; Salajanu, D.; Weyermann, D.; et al. Conterminous U.S. and Alaska forest type mapping using forest inventory and analysis data. Phot. Eng. Rem. Sen. 2008, 11, 1379–1388. [Google Scholar] [CrossRef]
- Mahatara, D.; Acharya, A.K.; Dhakal, B.P.; Sharma, D.K.; Ulak, S.; Paudel, P. Maxent modelling for habitat suitability of vulnerable tree Dalbergia latifolia in Nepal. Silva Fenn. 2021, 55, e10441. [Google Scholar] [CrossRef]
- Liu, D.; Lei, X.; Gao, W.; Guo, H.; Xie, Y.; Fu, L.; Lei, Y.; Li, Y.; Zhang, Z.; Tang, S. Mapping the potential distribution suitability of 16 tree species under climate change in northeastern China using Maxent modelling. J. For. Res. 2022, 33, 1739–1750. [Google Scholar] [CrossRef]
- Trasobares, A.; Mola-Yudego, B.; Aquilué, N.; González-Olabarria, J.R.; Garcia-Gonzalo, J.; García-Valdés, R.; De Cáceres, M. Nationwide climate-sensitive models for stand dynamics and forest scenario simulation. For. Ecol. Manag. 2022, 505, e119909. [Google Scholar] [CrossRef]
- Yu, Z.; Zhao, H.; Liu, S.; Zhou, G.; Fang, J.; Yu, G.; Tang, X.; Wang, W.; Yan, J.; Wang, G.; et al. Mapping forest type and age in China’s plantations. Sci. Total Environ. 2020, 744, e140790. [Google Scholar] [CrossRef]
- Hościło, A.; Lewandowska, A. Mapping forest type and tree species on a regional scale using multi-temporal Sentinel-2 data. Rem. Sen. 2019, 11, 929. [Google Scholar] [CrossRef]
- Paredes, D.; Cellini, J.M.; Lencinas, M.V.; Martínez Pastur, G. Influencia del paisaje en las cortas de protección en bosques de Nothofagus pumilio en Tierra del Fuego, Argentina: Cambios en la estructura forestal y respuesta de la regeneración. Bosque 2020, 41, 55–64. [Google Scholar] [CrossRef]
- Zak, M.; Cabido, M. Spatial patterns of the Chaco vegetation of central Argentina: Integration of remote sensing and phytosociology. Appl. Veg. Sci. 2002, 5, 213–226. [Google Scholar] [CrossRef]
- Corona, P.; Chirici, G.; Marchetti, M. Forest ecosystem inventory and monitoring as a framework for terrestrial natural renewable resource survey programmes. Plant Biosyst. 2002, 136, 69–82. [Google Scholar] [CrossRef]
- Soriano, M.; Zuidema, P.; Barber, C.; Mohren, F.; Ascarrunz, N.; Licona, J.C.; Peña-Claros, M. Commercial logging of timber species enhances Amazon (Brazil) nut populations: Insights from Bolivian managed forests. Forests 2021, 12, 1059. [Google Scholar] [CrossRef]
- Matangaran, J.R.; Anissa, I.N.; Adlan, Q.; Mujahid, M. Changes in floristic diversity and stand damage of tropical forests caused by logging operations in North Kalimantan, Indonesia. Biodiv. J. Biol. Div. 2022, 23, 6358–6365. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| REGION | PHE | RICH | SOC | BA | CC | DH | TOBV | ELE | AMT | ISO | AP |
|---|---|---|---|---|---|---|---|---|---|---|---|
| TDF | 1 | 80.2 bc | 154.4 b | 41.9 ab | 66.4 a | 18.6 b | 299.7 ab | 421.2 d | 3.5 a | 51.6 c | 655.6 e |
| 2 | 61.4 a | 163.3 b | 37.8 a | 74.1 c | 20.9 c | 207.5 a | 217.0 bc | 4.3 bc | 51.2 c | 506.5 c | |
| 3 | 88.1 c | 163.6 b | 56.7 d | 69.3 ab | 20.7 c | 373.9 b | 203.9 b | 4.6 c | 49.9 b | 448.1 b | |
| 4 | 90.1 c | 140.5 a | 44.9 b | 67.7 a | 15.7 a | 238.1 a | 107.2 a | 5.1 d | 48.6 a | 385.6 a | |
| 5 | 70.1 b | 160.3 b | 52.9 cd | 66.4 a | 18.7 b | 331.5 b | 382.9 d | 3.7 a | 51.0 c | 533.6 d | |
| 6 | 57.5 a | 161.9 b | 42.6 ab | 73.3 bc | 21.6 c | 258.7 a | 289.7 d | 4.1 ab | 51.1 c | 521.7 cd | |
| F (p) | 77.54 (<0.001) | 51.45 (<0.001) | 22.66 (<0.001) | 9.64 (<0.001) | 34.56 (<0.001) | 26.02 (<0.001) | 36.20 (<0.001) | 41.66 (<0.001) | 132.00 (<0.001) | 345.81 (<0.001) | |
| PAT | 7 | 62.4 b | 137.9 c | 34.3 a | 70.0 c | 18.7 bc | 250.4 a | 1062.5 a | 7.0 c | 52.4 c | 1046.7 d |
| 8 | 66.6 b | 154.8 e | 36.8 a | 71.7 c | 19.1 c | 254.9 a | 1013.4 a | 6.8 c | 51.0 b | 1167.5 e | |
| 9 | 66.3 b | 150.1 d | 43.6 b | 72.4 c | 20.6 d | 333.2 b | 1123.2 a | 6.1 b | 51.4 b | 1273.9 f | |
| 11 | 48.3 a | 127.0 b | 34.2 a | 65.5 b | 17.0 b | 243.2 a | 1385.1 b | 5.8 ab | 52.4 c | 785.0 b | |
| 12 | 49.1 a | 126.7 b | 34.6 a | 65.8 b | 17.7 b | 235.7 a | 1355.2 b | 6.1 b | 52.5 c | 899.0 c | |
| 13 | 49.5 a | 119.1 a | 36.6 a | 62.3 a | 15.2 a | 240.0 a | 1078.9 a | 5.5 a | 49.3 a | 588.9 a | |
| F (p) | 31.41 (<0.001) | 183.20 (<0.001) | 11.08 (<0.001) | 67.19 (<0.001) | 64.57 (<0.001) | 17.23 (<0.001) | 35.62 (<0.001) | 24.53 (<0.001) | 61.28 (<0.001) | 932.19 (<0.001) | |
| ESP | 17 | 19.5 b | 39.0 b | 10.3 a | 46.4 b | 7.7 b | 45.9 b | 315.0 d | 15.9 b | 48.6 c | 576.3 b |
| 18 | 3.2 a | 33.9 a | 10.2 a | 37.6 a | 6.9 a | 35.8 a | 155.2 c | 15.1 a | 47.5 a | 445.3 a | |
| 19 | 39.0 cd | 81.6 d | 17.2 d | 66.4 e | 12.4 e | 118.1 e | 47.7 a | 18.7 c | 47.9 ab | 1226.0 d | |
| 20 | 36.2 c | 73.1 c | 13.5 c | 60.7 d | 9.8 d | 91.6 d | 80.4 b | 18.8 c | 48.0 b | 1068.8 c | |
| 21 | 39.2 d | 77.3 d | 12.4 b | 55.9 c | 8.4 c | 82.5 c | 58.5 a | 18.9 d | 47.5 a | 1064.1 c | |
| F (p) | 733.59 (<0.001) | 5368.24 (<0.001) | 182.12 (<0.001) | 1118.24 (<0.001) | 468.36 (<0.001) | 1323.15 (<0.001) | 824.95 (<0.001) | 4599.80 (<0.001) | 196.28 (<0.001) | 6567.81 (<0.001) | |
| MON | 22 | -- | 36.5 a | 5.9 a | 27.9 a | 6.2 ab | 28.0 ab | 185.1 a | 15.5 | 48.4 a | 292.9 b |
| 23 | -- | 39.4 b | 6.3 a | 28.5 a | 6.0 a | 25.6 a | 448.5 b | 15.4 | 48.3 a | 289.9 b | |
| 24 | -- | 36.6 a | 7.6 bc | 30.4 b | 6.3 b | 35.8 c | 1461.7 c | 15.6 | 50.4 b | 194.6 a | |
| 25 | -- | 36.8 a | 8.3 c | 32.7 b | 6.5 b | 31.9 bc | 202.1 a | 15.6 | 48.1 a | 361.4 c | |
| F (p) | -- | 7.99 (<0.001) | 18.49 (<0.001) | 6.70 (<0.001) | 8.58 (<0.001) | 38.13 (<0.001) | 196.66 (<0.001) | 0.98 (0.400) | 40.17 (<0.001) | 207.66 (<0.001) | |
| PCH | 38 | 30.6 f | 58.6 f | 13.8 i | 60.5 c | 10.7 f | 81.9 jk | 858.3 f | 17.5 b | 50.1 cd | 596.2 b |
| 39 | 30.1 f | 69.0 k | 11.6 ef | 66.6 e | 9.5 bc | 66.1 fgh | 71.4 a | 19.9 d | 49.8 c | 987.2 j | |
| 40 | 35.3 g | 58.0 f | 13.6 hi | 64.3 d | 11.2 gh | 85.5 k | 638.7 e | 19.4 c | 51.7 gh | 642.4 de | |
| 41 | 18.3 b | 41.5 b | 10.7 d | 45.4 a | 7.6 a | 55.5 d | 925.3 g | 16.8 a | 49.3 b | 527.7 a | |
| 42 | 56.2 j | 61.6 h | 12.6 g | 73.2 i | 10.1 d | 66.1 gh | 131.7 b | 22.4 k | 54.3 j | 847.7 i | |
| 43 | 29.9 f | 60.9 gh | 11.4 e | 66.9 ef | 10.1 d | 57.5 de | 155.0 b | 21.4 fg | 51.3 fg | 801.7 h | |
| 44 | 42.4 h | 59.8 fg | 12.0 f | 70.0 h | 10.7 ef | 64.8 fg | 215.1 c | 22.1 i | 52.5 i | 766.2 g | |
| 45 | 23.9 d | 48.7 d | 10.0 c | 60.7 c | 9.5 b | 47.1 c | 305.9 d | 20.6 e | 50.2 cd | 621.5 cd | |
| 46 | 34.9 fg | 69.6 k | 11.7 ef | 65.9 de | 10.0 cd | 59.5 de | 280.5 bcd | 22.3 jk | 51.5 fgh | 791.7 gh | |
| 47 | 20.8 c | 53.8 e | 9.8 c | 59.6 c | 9.4 b | 45.8 c | 232.9 cd | 20.5 e | 50.3 d | 662.9 e | |
| 48 | 26.5 e | 47.0 c | 9.2 b | 59.7 c | 9.5 b | 42.4 b | 253.4 d | 21.4 fg | 50.7 e | 609.7 bc | |
| 49 | 5.6 a | 36.7 a | 7.3 a | 48.1 b | 7.9 a | 26.9 a | 223.0 cd | 20.4 e | 48.0 a | 530.4 a | |
| 50 | 39.1 g | 62.5 hi | 12.6 g | 68.5 fg | 10.3 d | 68.1 h | 334.7 d | 21.8 hi | 51.9 h | 708.3 f | |
| 51 | 49.1 i | 64.3 ij | 13.6 i | 68.2 g | 11.3 h | 81.0 j | 78.4 a | 21.7 gh | 51.5 g | 1126.7 k | |
| 52 | 39.3 gh | 53.8 e | 14.7 j | 76.1 j | 11.7 h | 86.1 jk | 256.3 bcd | 22.3 hij | 53.1 i | 652.3 cde | |
| 53 | 38.1 g | 64.6 j | 13.3 h | 66.7 e | 10.9 fg | 76.9 i | 77.1 a | 21.5 fgh | 51.4 fg | 1160.3 l | |
| 54 | 29.8 ef | 66.4 jk | 11.8 ef | 63.0 d | 10.3 de | 61.0 ef | 112.9 ab | 20.4 e | 50.9 ef | 1017.8 j | |
| F (p) | 460.20 (<0.001) | 784.09 (<0.001) | 588.23 (<0.001) | 725.22 (<0.001) | 369.69 (<0.001) | 739.89 (<0.001) | 1044.68 (<0.001) | 963.78 (<0.001) | 331.25 (<0.001) | 2617.41 (<0.001) | |
| YUN | 32 | 65.9 e | 84.9 e | 17.9 c | 78.9 d | 18.1 c | 140.8 c | 1187.5 c | 17.6 c | 53.3 b | 729.3 c |
| 33 | 54.2 c | 69.9 ab | 15.7 a | 75.8 bc | 15.8 a | 119.3 a | 765.2 b | 19.5 d | 52.0 a | 742.5 c | |
| 34 | 73.5 e | 74.2 bc | 18.1 c | 78.1 d | 18.0 c | 137.8 c | 616.8 a | 20.8 e | 51.5 a | 861.7 d | |
| 35 | 60.4 d | 75.6 c | 17.6 bc | 77.8 d | 17.1 b | 136.1 c | 620.4 a | 21.1 e | 51.5 a | 987.9 e | |
| 36 | 6.6 a | 81.3 d | 19.4 d | 71.5 a | 15.3 a | 156.0 d | 2564.8 e | 12.5 a | 55.8 c | 285.9 a | |
| 37 | 28.1 b | 67.6 a | 17.3 b | 75.3 b | 15.9 a | 129.6 b | 1438.8 d | 16.8 b | 53.1 b | 528.9 b | |
| F (p) | 242.15 (<0.001) | 90.63 (<0.001) | 73.63 (<0.001) | 49.23 (<0.001) | 59.12 (<0.001) | 58.91 (<0.001) | 477.84 (<0.001) | 524.01 (<0.001) | 45.07 (<0.001) | 1919.23 (<0.001) | |
| AF | 26 | 77.9 c | 95.9 c | 19.5 c | 77.4 d | 21.1 c | 134.1 c | 511.9 e | 18.6 a | 57.7 d | 1856.4 e |
| 27 | 64.2 b | 98.1 d | 18.7 b | 75.1 bc | 20.3 b | 122.4 b | 199.5 b | 20.3 c | 54.9 b | 1596.2 a | |
| 29 | 77.1 c | 96.6 cd | 19.3 c | 76.0 c | 21.0 c | 135.4 c | 331.2 d | 19.7 b | 56.0 c | 1724.4 c | |
| 30 | 68.7 b | 93.0 b | 18.7 b | 74.7 b | 20.4 b | 122.9 b | 254.2 c | 20.1 c | 55.2 b | 1737.9 d | |
| 31 | 33.4 a | 82.7 a | 17.6 a | 73.1 a | 18.2 a | 112.2 a | 156.9 a | 20.7 d | 52.9 a | 1649.1 b | |
| F (p) | 142.20 (<0.001) | 78.96 (<0.001) | 87.54 (<0.001) | 32.12 (<0.001) | 63.44 (<0.001) | 117.01 (<0.001) | 275.91 (<0.001) | 268.19 (<0.001) | 206.00 (<0.001) | 893.30 (<0.001) |
| REGION | PHE | Plots | FT-1 | FT-2 | FT-3 | MONO | BI | MULTI |
|---|---|---|---|---|---|---|---|---|
| Country | 3741 | 50 | 115 | 1990 | 25.9% | 32.2% | 41.9% | |
| TDF | Total | 56 | 3 | 3 | 3 | 100.0% | 0.0% | 0.0% |
| 1 | 1 | 1 | 1 | 1 | 100.0% | 0.0% | 0.0% | |
| 2 | 7 | 2 | 2 | 2 | 100.0% | 0.0% | 0.0% | |
| 3 | 23 | 2 | 2 | 2 | 100.0% | 0.0% | 0.0% | |
| 4 | 12 | 3 | 3 | 3 | 100.0% | 0.0% | 0.0% | |
| 5 | 8 | 3 | 3 | 3 | 100.0% | 0.0% | 0.0% | |
| 6 | 5 | 2 | 2 | 2 | 100.0% | 0.0% | 0.0% | |
| PAT | Total | 172 | 5 | 5 | 25 | 86.0% | 13.4% | 0.6% |
| 7 | 20 | 4 | 4 | 12 | 45.0% | 50.0% | 5.0% | |
| 8 | 28 | 5 | 5 | 11 | 82.1% | 17.9% | 0.0% | |
| 9 | 21 | 5 | 5 | 6 | 90.5% | 9.5% | 0.0% | |
| 11 | 21 | 2 | 2 | 4 | 95.2% | 4.8% | 0.0% | |
| 12 | 38 | 4 | 4 | 8 | 86.8% | 13.2% | 0.0% | |
| 13 | 44 | 3 | 3 | 3 | 100.0% | 0.0% | 0.0% | |
| ESP | Total | 251 | 6 | 11 | 112 | 49.0% | 36.7% | 14.3% |
| 17 | 99 | 2 | 4 | 21 | 82.8% | 17.2% | 0.0% | |
| 18 | 11 | 1 | 3 | 6 | 72.7% | 27.3% | 0.0% | |
| 20 | 57 | 6 | 8 | 47 | 21.0% | 47.4% | 31.6% | |
| 21 | 84 | 6 | 7 | 52 | 25.0% | 53.6% | 21.4% | |
| MON | Total | 87 | 4 | 10 | 32 | 72.4% | 26.4% | 1.2% |
| 22 | 1 | 1 | 1 | 1 | 100.0% | 0.0% | 0.0% | |
| 23 | 58 | 4 | 9 | 24 | 69.0% | 31.0% | 0.0% | |
| 24 | 23 | 4 | 7 | 12 | 73.9% | 21.7% | 4.4% | |
| 25 | 5 | 1 | 2 | 2 | 100.0% | 0.0% | 0.0% | |
| PCH | Total | 2725 | 30 | 73 | 1462 | 18.7% | 35.3% | 46.0% |
| 38 | 85 | 14 | 26 | 66 | 27.1% | 43.5% | 29.4% | |
| 39 | 75 | 15 | 23 | 59 | 17.3% | 32.0% | 50.7% | |
| 40 | 37 | 11 | 16 | 36 | 18.9% | 24.3% | 56.8% | |
| 41 | 159 | 8 | 22 | 73 | 47.8% | 42.1% | 10.1% | |
| 42 | 149 | 14 | 21 | 129 | 14.1% | 23.5% | 62.4% | |
| 43 | 116 | 13 | 21 | 98 | 11.3% | 35.3% | 53.4% | |
| 44 | 373 | 23 | 34 | 281 | 9.9% | 30.0% | 60.1% | |
| 45 | 187 | 12 | 23 | 126 | 14.4% | 48.2% | 37.4% | |
| 46 | 42 | 10 | 13 | 37 | 7.1% | 31.0% | 61.9% | |
| 47 | 259 | 14 | 27 | 171 | 18.6% | 37.8% | 43.6% | |
| 48 | 455 | 16 | 27 | 248 | 16.0% | 42.6% | 41.4% | |
| 49 | 139 | 8 | 11 | 56 | 33.1% | 54.0% | 12.9% | |
| 50 | 109 | 14 | 24 | 101 | 11.0% | 32.1% | 56.9% | |
| 51 | 282 | 25 | 46 | 236 | 15.6% | 20.9% | 63.5% | |
| 52 | 40 | 12 | 16 | 36 | 25.0% | 35.0% | 40.0% | |
| 53 | 146 | 19 | 32 | 129 | 19.2% | 27.4% | 53.4% | |
| 54 | 72 | 13 | 21 | 47 | 40.3% | 25.0% | 34.7% | |
| YUN | Total | 289 | 25 | 41 | 242 | 20.4% | 29.8% | 49.8% |
| 32 | 80 | 15 | 18 | 74 | 15.0% | 26.2% | 58.8% | |
| 33 | 49 | 15 | 17 | 45 | 18.3% | 32.7% | 49.0% | |
| 34 | 19 | 12 | 14 | 19 | 5.3% | 26.3% | 68.4% | |
| 35 | 62 | 15 | 21 | 60 | 8.1% | 27.4% | 64.5% | |
| 36 | 14 | 4 | 4 | 5 | 85.7% | 0.0% | 14.3% | |
| 37 | 65 | 18 | 22 | 57 | 30.8% | 41.5% | 27.7% | |
| AF | Total | 161 | 19 | 28 | 160 | 4.4% | 13.0% | 82.6% |
| 26 | 31 | 12 | 14 | 31 | 0.0% | 12.9% | 87.1% | |
| 27 | 21 | 9 | 10 | 21 | 9.5% | 9.5% | 81.0% | |
| 29 | 43 | 13 | 17 | 43 | 0.0% | 16.3% | 83.7% | |
| 30 | 49 | 17 | 20 | 49 | 6.2% | 12.2% | 81.6% | |
| 31 | 17 | 11 | 11 | 17 | 11.8% | 11.8% | 76.4% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez Pastur, G.J.; Loto, D.; Rodríguez-Souilla, J.; Silveira, E.M.O.; Cellini, J.M.; Peri, P.L. Different Approaches of Forest Type Classifications for Argentina Based on Functional Forests and Canopy Cover Composition by Tree Species. Resources 2024, 13, 62. https://doi.org/10.3390/resources13050062
Martínez Pastur GJ, Loto D, Rodríguez-Souilla J, Silveira EMO, Cellini JM, Peri PL. Different Approaches of Forest Type Classifications for Argentina Based on Functional Forests and Canopy Cover Composition by Tree Species. Resources. 2024; 13(5):62. https://doi.org/10.3390/resources13050062
Chicago/Turabian StyleMartínez Pastur, Guillermo J., Dante Loto, Julián Rodríguez-Souilla, Eduarda M. O. Silveira, Juan M. Cellini, and Pablo L. Peri. 2024. "Different Approaches of Forest Type Classifications for Argentina Based on Functional Forests and Canopy Cover Composition by Tree Species" Resources 13, no. 5: 62. https://doi.org/10.3390/resources13050062