7,8-dimethoxycoumarin Attenuates the Expression of IL-6, IL-8, and CCL2/MCP-1 in TNF-α-Treated HaCaT Cells by Potentially Targeting the NF-κB and MAPK Pathways

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Culture
2.3. Cell Viability Assay
2.4. Measurement of Cytokine Concentration
2.5. Western Blot Assay
2.6. Statistical Analysis
3. Results
3.1. Effect of DMC on Viability of HaCaT Cells
3.2. Effects of DMC on the Expression of Pro-Inflammatory Cytokines in TNF-α-treated HaCaT Cells
3.3. Effect of TNF-α on the MAPKs and NF-κB Pathway in HaCaT Cells
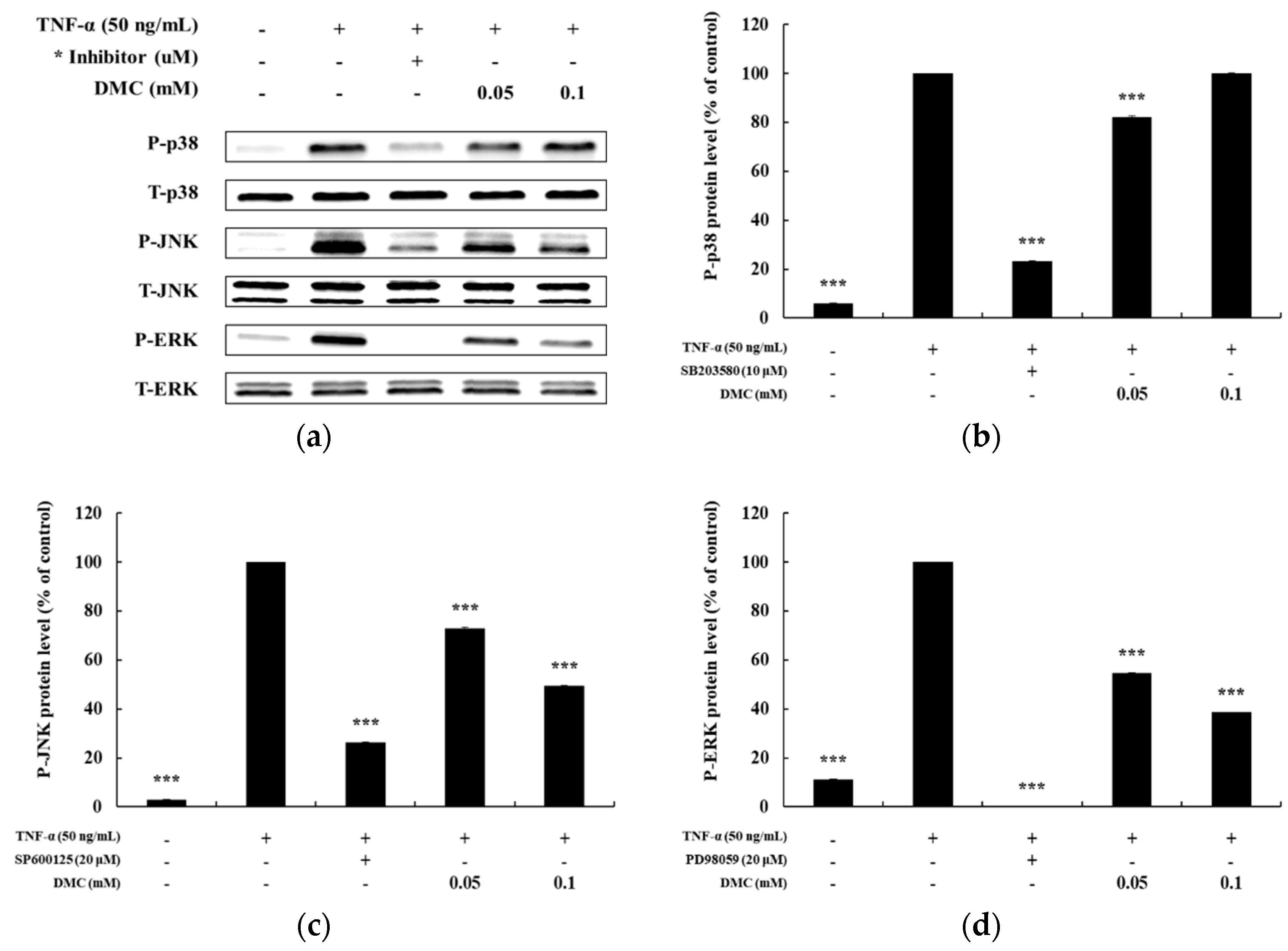
3.4. Effect of DMC on the MAPK Pathway in TNF-α-Treated HaCaT Cells
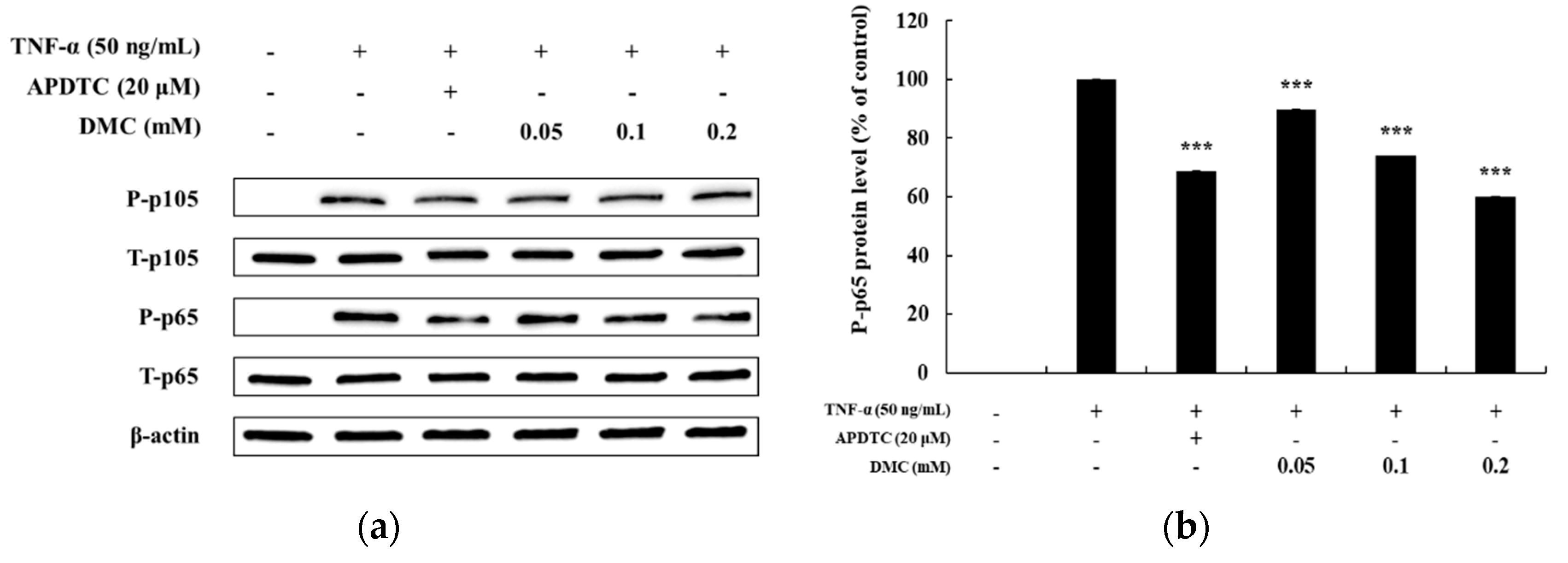
3.5. Effect of DMC on the NF-κB Pathway in TNF-α Treated HaCaT Cells
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Garcia, M.; Morello, E.; Garnier, J.; Barrault, C.; Garnier, M.; Burucoa, C.; Lecron, J.; Si-Tahar, M.; Bernard, F.X.; Bodet, C. Pseudomonas aeruginosa flagellum is critical for invasion, cutaneous persistence and induction of inflammatory response of skin epidermis. Virulence 2018, 9, 1163–1175. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Jiang, H.; Li, W.; Qiang, M.; Dong, T.; Li, H. Role of Vitamin C in Skin Diseases. Front. Physiol. 2018, 9, 819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.H.; Yoo, J.M.; Cho, W.K.; Ma, J.Y. Anti-inflammatory effects of Sanguisorbae Radix water extract on the suppression of mast cell degranulation and STAT-1/Jak-2 activation in BMMCs and HaCaT keratinocytes. BMC Complement. Altern. Med. 2016, 16, 347. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Yoo, J.M.; Lee, E.; Lee, B.; Cho, W.K.; Park, K.I.; Yeul, M.J. Anti-inflammatory effects of PerillaeHerbaethanolic extract against TNF-α/IFN-γ-stimulated human keratinocyte HaCaT cells. J. Ethnopharmacol. 2018, 211, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Liu, J.; Wang, J.; Luo, Q.; Zhang, H.; Liu, B.; Xu, F.; Pang, Q.; Liu, Y.; Dong, J. Icariin inhibits TNF-α/IFN-γ induced inflammatory response via inhibition of the substance P and p38-MAPK signaling pathway in human keratinocytes. Int. Immunopharmacol. 2015, 29, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Hwang, Y.H.; Gu, M.J.; Cho, W.K.; Ma, J.Y. Ethanol extracts of Sanguisorba officinalis L. suppress TNF-α/IFN-γ-induced pro-inflammatory chemokine production in HaCaT cells. Phytomedicine 2015, 22, 1262–1268. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, X.; Wang, H.; Zhao, B.; Wu, X.; Su, L.; Xie, S.; Wang, Y.; Li, J.; Liu, J.; et al. PKCζ as a promising therapeutic target for TNFα-induced inflammatory disorders in chronic cutaneous wounds. Int. J. Mol. Med. 2017, 40, 1335–1346. [Google Scholar] [CrossRef]
- Cho, J.W.; Lee, K.S.; Kim, C.W. Curcumin attenuates the expression of IL-1beta, IL-6, and TNF-alpha as well as cyclin E in TNF-alpha-treated HaCaT cells; NF-kappaB and MAPKs as potential upstream targets. Int. J. Mol. Med. 2007, 19, 469–474. [Google Scholar]
- Choi, H.J.; Lee, J.H.; Jung, Y.S. (+)-Nootkatone inhibits tumor necrosis factor α/interferon γ-induced production of chemokines in HaCaT cells. BiochemBiophys. Res. Commun. 2014, 447, 278–284. [Google Scholar] [CrossRef]
- Jung, T.D.; Choi, S.I.; Choi, S.H.; Cho, B.Y.; Sim, W.S.; Han-Xionggao; Lee, S.J.; Park, S.J.; Kim, D.B.; Kim, Y.C.; et al. Changes in the Anti-Allergic Activities of Sesame by Bioconversion. Nutrients 2018, 10, 210. [Google Scholar] [CrossRef]
- Jeong, S.J.; Lim, H.S.; Seo, C.S.; Kim, J.H.; Jin, S.E.; Yoo, S.R.; Shin, H.K. Traditional herbal formula Jakyakgamcho-tang (Paeonia lactiflora and Glycyrrhiza uralensis) impairs inflammatory chemokine production by inhibiting activation of STAT1 and NF-κB in HaCaT cells. Phytomedicine 2015, 22, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Dai, Y.W.; Peng, H.L.; Kang, C.W.; Kuo, C.Y.; Liou, C.J. Phloretin ameliorates chemokines and ICAM-1 expression via blocking of the NF-κB pathway in the TNF-α-treatedHaCaT human keratinocytes. Int. Immunopharmacol. 2015, 27, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.Y.; Zhang, A.L.; Chen, D.; Xue, C.C.; Lenon, G.B. Chinese herbal medicine for atopic dermatitis: A systematic review. J. Am. Acad. Dermatol. 2013, 69, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Kudo, K.; Hasegawa, S.; Suzuki, Y.; Hirano, R.; Wakiguchi, W.; Kittaka, S. 1α, 25-Dihydroxyvitamin D(3) inhibits vascular cellular adhesion molecule-1 expressionand interleukin-8 production in human coronary arterial endothelial cells. J. Steroid Biochem. Mol. Biol. 2012, 132, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Schooling, C.M.; Au Yeung, S.L.; Freeman, G.; Cowling, B.J. The effect of statins on testosterone in men and women, a systematic review and meta-analysis of randomized controlled trials. BMC Med. 2013, 11, 1714–7015. [Google Scholar] [CrossRef]
- Wu, C.; Feng, D.; Ma, H.; Xie, H.; Wang, H.; Wang, J. Effect of Pinusmassoniana bark extract on IFN-gamma-induced ICAM-1 expression in HaCaT human keratinocytes. J. Ethnopharmacol. 2009, 122, 48–53. [Google Scholar] [CrossRef]
- Lee, J.H.; Jeon, Y.D.; Lee, Y.M.; Kim, D.K. The suppressive effect of puerarin on atopic dermatitis-like skin lesions through regulation of inflammatory mediatorsin vitro and in vivo. Biochem. Biophys. Res. Commun. 2018, 498, 707–714. [Google Scholar] [CrossRef]
- Choi, J.K.; Jang, Y.H.; Lee, S.; Lee, S.R.; Choi, Y.A.; Jin, M.; Choi, J.H.; Park, J.H.; Park, P.H.; Choi, H.; et al. Chrysin attenuates atopic dermatitis by suppressing inflammation of keratinocytes. Food Chem. Toxicol. 2017, 110, 142–150. [Google Scholar] [CrossRef]
- Li, Z.; Hu, J.; Sun, M.; Song, X.; Li, G.; Liu, Y.; Li, G.; Ji, H.; Liu, G.; Chen, N. In vitro and in vivo anti-inflammatory effects of IMMLG5521, a coumarin derivative. Int. Immunopharmacol. 2013, 17, 400–403. [Google Scholar] [CrossRef]
- Pérez Gutiérrez, R.M.; Vargas Solis, R.; Garcia Baez, E.; Gallardo Navarro, Y. Hypoglycemic activity of constituents from Astianthusviminalis in normal and streptozotocin-induced diabetic mice. J. Nat. Med. 2009, 63, 393–401. [Google Scholar] [CrossRef]
- Sood, S.; Muthuraman, A.; Gill, N.S.; Bali, M.; Sharma, P.D. Role of 7,8-dimethoxycoumarin in anti-secretary and anti-inflammatory action on pyloric ligation-induced gastritis in rats. J. Asian Nat. Prod. Res. 2010, 12, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Muthuraman, A.; Sood, S.; Ramesh, M.; Puri, K.D.; Peters, A.; Chauhan, A.; Arora, P.K.; Rana, A. Therapeutic potential of 7,8-dimethoxycoumarin on cisplatin- and ischemia/reperfusion injury-induced acute renal failure in rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2012, 385, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Misra, L.N.; Wouatsa, N.A.; Kumar, S.; Kumar, R.V.; Tchoumbougnang, F. Antibacterial, cytotoxic activities and chemical composition of fruits of two Cameroonian Zanthoxylum species. J. Ethnopharmacol. 2013, 148, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.; Xu, M.; Yang, J.; Xi, H.; Xu, D. Chemical studies on the constituents of Daphne koreane Nakai. ZhongguoZhong Yao ZaZhi 1990, 15, 609–610. [Google Scholar] [CrossRef]
- Sung, Y.Y.; Kim, Y.S.; Kim, H.K. Illiciumverum extract inhibits TNF-α- and IFN-γ-induced expression of chemokines and cytokines in human keratinocytes. J. Ethnopharmacol. 2012, 144, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.H.; Jeong, G.S. Fisetin inhibits TNF-α-treated inflammatory action and hydrogen peroxide-induced oxidative damage in human keratinocyte HaCaT cells through PI3K/AKT/Nrf-2-mediated heme oxygenase-1 expression. Int. Immunopharmacol. 2015, 29, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wu, L. Coumarins from the roots of Angelica dahurica cause anti-allergic inflammation. Exp. Ther. Med. 2017, 14, 874–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, T.M.; Franco, D.P.; Vitorio, F.; Kummerle, A.E. Coumarin Compounds in Medicinal Chemistry: Some Important Examples from the Last Years. Curr. Top. Med. Chem. 2018, 18, 124–148. [Google Scholar] [CrossRef] [PubMed]
- Stefanachi, A.; Leonetti, F.; Pisani, L.; Catto, M.; Carotti, A. Coumarin: A Natural, Privileged and Versatile Scaffold for Bioactive Compounds. Molecules 2018, 23, 250. [Google Scholar] [CrossRef]
- Fiorito, S.; Epifano, F.; Preziuso, F.; Cacciatore, I.; di Stefano, A.; Taddeo, V.A.; de Medina, P.; Genovese, S. Natural oxyprenylatedcoumarins are modulators of melanogenesis. Eur. J. Med. Chem. 2018, 152, 274–282. [Google Scholar] [CrossRef]
- Yang, J.Y.; Koo, J.H.; Song, Y.G.; Kwon, K.B.; Lee, J.H.; Sohn, H.S.; Park, B.H.; Jhee, E.C.; Park, J.W. Stimulation of melanogenesis by scoparone in B16 melanoma cells. Acta Pharmacol. Sin. 2006, 27, 1467–1473. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, X.; Li, Z.; Zhang, L.; Liu, Y.; Ding, H.; Yin, S. Isofraxidin, a coumarin component improves high-fat diet induced hepatic lipid homeostasis disorder and macrophage inflammation in mice. Food Funct. 2017, 8, 2886–2896. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Zhou, T.; Wang, J.; Sang, X.; Lan, L.; Luo, L.; Yin, Z. Daphnetin reduces endotoxin lethality in mice and decreases LPS-induced inflammation in Raw264.7 cells via suppressing JAK/STATs activation and ROS production. Inflamm. Res. 2017, 66, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Jang, S.I.; Kim, Y.J.; Chung, H.T.; Yun, Y.G.; Kang, T.H.; Jeong, O.S.; Kim, Y.C. Scopoletin suppresses pro-inflammatory cytokines and PGE2 from LPS-stimulated cell line, RAW 264.7 cells. Fitoterapia 2004, 75, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Liu, Z.; Zhang, T.; Sun, S.; Ye, J.; Li, Z.; Mao, L.; Ren, J. Enhancement of Anti-Inflammatory Properties of Nobiletin in Macrophages by a Nano-Emulsion Preparation. J. Agric. Food Chem. 2018, 66, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Kim, S.; Kim, J.H.; Lee, G.S.; Lee, J.N.; Lee, N.H.; Hyun, C.G. Pratol, an O-Methylated Flavone, Induces Melanogenesis in B16F10 Melanoma Cells via p-p38 and p-JNK Upregulation. Molecules 2017, 22, 1704. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.S.; Ko, H.C.; Kim, S.S.; Park, K.J.; An, H.J.; Choi, Y.H.; Kim, S.J.; Lee, N.H.; Hyun, C.G. Tangeretin triggers melanogenesis through the activation of melanogenic signaling proteins and sustained extracellular signal- regulated kinase in B16/F10 murine melanoma cells. Nat. Prod. Commun. 2015, 10, 389–392. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.J.; Koo, D.H.; Kang, G.J.; Han, S.C.; Lee, B.W.; Koh, Y.S.; Hyun, J.W.; Lee, N.H.; Ko, M.H.; Kang, H.K.; et al. Dieckol, a Component of Ecklonia cava, Suppresses the Production of MDC/CCL22 via Down-Regulating STAT1 Pathway in Interferon-γ Stimulated HaCaT Human Keratinocytes. Biomol. Ther. 2015, 23, 238–244. [Google Scholar] [CrossRef]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, N.; Chung, Y.C.; Kang, C.I.; Park, S.-M.; Hyun, C.-G. 7,8-dimethoxycoumarin Attenuates the Expression of IL-6, IL-8, and CCL2/MCP-1 in TNF-α-Treated HaCaT Cells by Potentially Targeting the NF-κB and MAPK Pathways. Cosmetics 2019, 6, 41. https://doi.org/10.3390/cosmetics6030041
Lee N, Chung YC, Kang CI, Park S-M, Hyun C-G. 7,8-dimethoxycoumarin Attenuates the Expression of IL-6, IL-8, and CCL2/MCP-1 in TNF-α-Treated HaCaT Cells by Potentially Targeting the NF-κB and MAPK Pathways. Cosmetics. 2019; 6(3):41. https://doi.org/10.3390/cosmetics6030041
Chicago/Turabian StyleLee, Nari, You Chul Chung, Choon Il Kang, Sung-Min Park, and Chang-Gu Hyun. 2019. "7,8-dimethoxycoumarin Attenuates the Expression of IL-6, IL-8, and CCL2/MCP-1 in TNF-α-Treated HaCaT Cells by Potentially Targeting the NF-κB and MAPK Pathways" Cosmetics 6, no. 3: 41. https://doi.org/10.3390/cosmetics6030041