
Aggressiveness in Italian Children with ADHD: MAOA Gene Polymorphism Involvement

 ,
, 
Abstract
1. Introduction
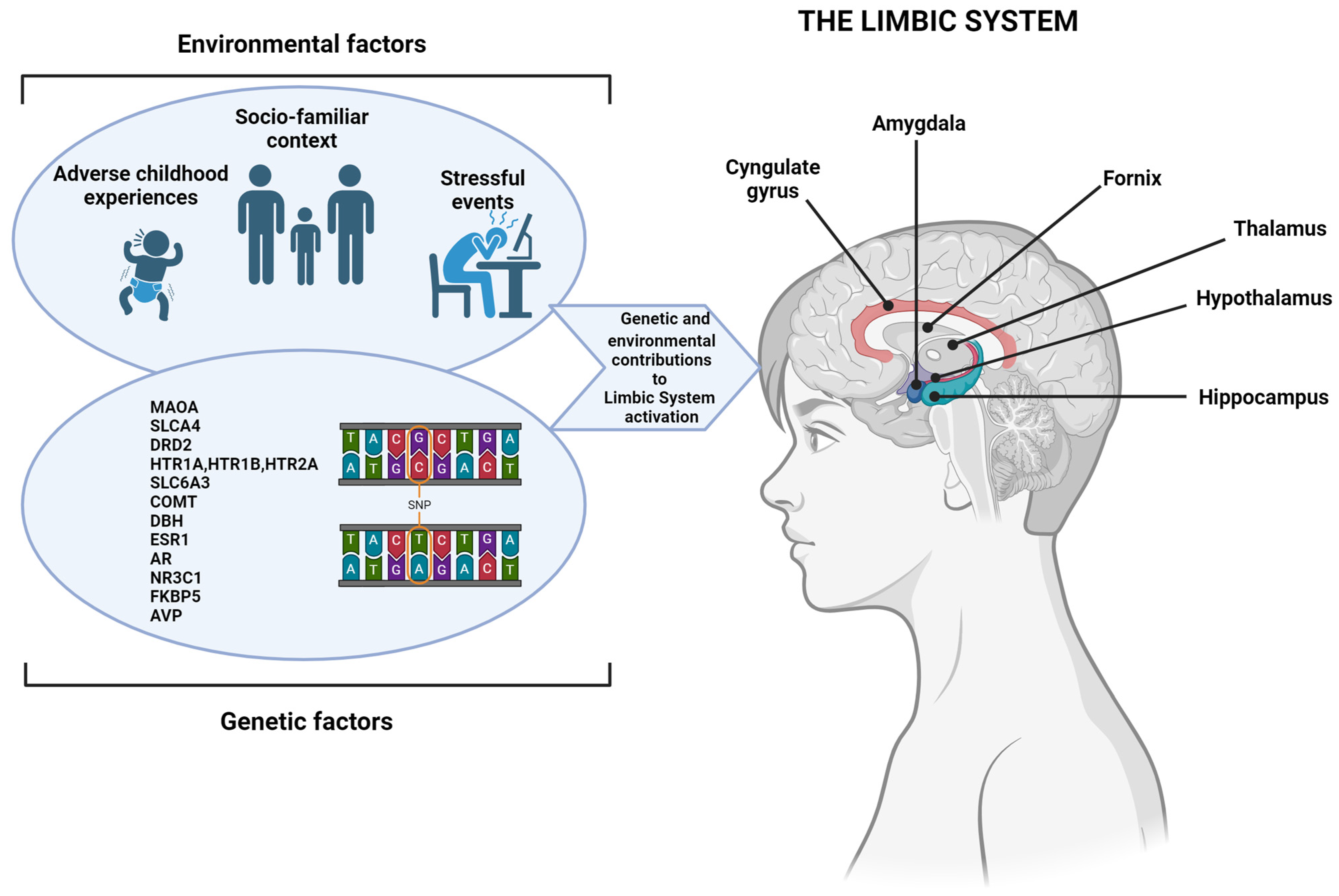
1.1. Characteristics and Prevalence of ADHD
1.2. Factors Associated with ADHD
1.3. Aim and Scope
2. Materials and Methods
2.1. Population Sample and Diagnostic Assessment
2.2. Genotyping
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gnanavel, S.; Sharma, P.; Kaushal, P.; Hussain, S. Attention deficit hyperactivity disorder and comorbidity: A review of literature. World J. Clin. Cases 2019, 7, 2420–2426. [Google Scholar] [CrossRef] [PubMed]
- Curatolo, P.; Paloscia, C.; D’Agati, E.; Moavero, R.; Pasini, A. The neurobiology of attention deficit/hyperactivity disorder. Eur. J. Paediatr. Neurol. 2009, 13, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Salari, N.; Ghasemi, H.; Abdoli, N.; Rahmani, A.; Shiri, M.H.; Hashemian, A.H.; Akbari, H.; Mohammadi, M. The global prevalence of ADHD in children and adolescents: A systematic review and meta-analysis. Ital. J. Pediatr. 2023, 49, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Reale, L.; Bonati, M. ADHD prevalence estimates in Italian children and adolescents: A methodological issue. Ital. J. Pediatr. 2018, 44, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Arnold, L.E. Sex differences in ADHD: Conference summary. J. Abnorm. Child. Psychol. 1996, 24, 555–569. [Google Scholar] [CrossRef]
- Gaub, M.; Carlson, C.L. Gender differences in ADHD: A meta-analysis and critical review. J. Am. Acad. Child. Adolesc. Psychiatry 1997, 36, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Rucklidge, J.J. Gender differences in attention-deficit/hyperactivity disorder. Psychiatr. Clin. N. Am. 2010, 33, 357–373. [Google Scholar] [CrossRef]
- Russell, G.; Rodgers, L.R.; Ukoumunne, O.C.; Ford, T. Prevalence of parent-reported ASD and ADHD in the UK: Findings from the Millennium Cohort Study. J. Autism Dev. Disord. 2014, 44, 31–40. [Google Scholar] [CrossRef]
- Ronald, A.; Simonoff, E.; Kuntsi, J.; Asherson, P.; Plomin, R. Evidence for overlapping genetic influences on autistic and ADHD behaviours in a community twin sample. J. Child. Psychol. Psychiatry Allied Discip. 2008, 49, 535–542. [Google Scholar] [CrossRef]
- Gidziela, A.; Ahmadzadeh, Y.I.; Michelini, G.; Allegrini, A.G.; Agnew-Blais, J.; Lau, L.Y.; Duret, M.; Procopio, F.; Daly, E.; Ronald, A.; et al. A meta-analysis of genetic effects associated with neurodevelopmental disorders and co-occurring conditions. Nat. Hum. Behav. 2023, 7, 642–656. [Google Scholar] [CrossRef]
- Hu, Y.; Ye, B.; Ma, T.; Feng, Z.; Chen, X.; Huang, D.; Liu, L. Exposure to COVID-19 and aggression: The mediating role of anxiety and the moderating role of rumination. Curr. Psychol. 2023, 22, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kirley, A.; Hawi, Z.; Daly, G.; McCarron, M.; Mullins, C.; Millar, N.; Waldman, I.; Fitzgerald, M.; Gill, M. Dopaminergic System Genes. in ADHD: Toward a Biological Hypothesis. Neuropsychopharmacology 2002, 27, 607–619. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Barr, C.R.; Misener, V.L. Dopamine system genes and ADHD: A review of the evidence. Future Neurol. 2008, 3, 705–728. [Google Scholar] [CrossRef]
- Faraone, S.V.; Larsson, H. Genetics of attention deficit hyperactivity disorder. Mol. Psychiatry 2019, 24, 562–575. [Google Scholar] [CrossRef] [PubMed]
- Balogh, L.; Pulay, A.J.; Réthelyim, J.M. Genetics in the ADHD Clinic: How Can Genetic Testing Support the Current Clinical Practice? Front. Psychol. 2022, 13, 751041–751058. [Google Scholar] [CrossRef] [PubMed]
- Saylor, K.E.; Amann, B.H. Impulsive aggression as a comorbidity of attention-deficit/hyperactivity disorder in children and adolescents. J. Child. Adolesc. Psychopharmacol. 2016, 26, 19–25. [Google Scholar] [CrossRef]
- Drechsler, R.; Brem, S.; Brandeis, D.; Grünblatt, E.; Berger, G.; Walitza, S. ADHD: Current Concepts and Treatments in Children and Adolescents. Neuropediatrics 2020, 51, 315–335. [Google Scholar] [CrossRef] [PubMed]
- Brevik, E.J.; van Donkelaar, M.M.; Weber, H.; Sánchez-Mora, C.; Jacob, C.; Rivero, O.; Kittel-Schneider, S.; Garcia-Martínez, I.; Aebi, M.; van Hulzen, K.; et al. Genome-wide analyses of aggressiveness in attention-deficit hyperactivity disorder. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2016, 171, 733–747. [Google Scholar] [CrossRef]
- Kolla, N.J.; Bortolato, M. The role of monoamine oxidase A in the neurobiology of aggressive, antisocial, and violent behavior: A tale of mice and men. Prog. Neurobiol. 2020, 194, 101875–101898. [Google Scholar] [CrossRef]
- Xu, M.K.; Gaysina, D.; Tsonaka, R.; Morin, A.J.S.; Croudace, T.J.; Barnett, J.H.; Houwing-Duistermaat, J.; Richards, M.; Jones, P.B.; LHA Genetics Group. Monoamine Oxidase A (MAOA) Gene and Personality Traits from Late Adolescence through Early Adulthood: A Latent Variable Investigation. Front. Psychol. 2017, 8, 1736–1748. [Google Scholar] [CrossRef]
- Manuck, S.B.; Flory, J.D.; Ferrell, R.E.; Mann, J.J.; Muldoon, M.F. A regulatory polymorphism of the monoamine oxidase-A gene may be associated with variability in aggression, impulsivity, and central nervous system serotonergic responsivity. Psychiatry Res. 2000, 95, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Volavka, J.; Bilder, R.; Nolan, K. Catecholamines and aggression: The role of COMT and MAO polymorphisms. Ann. N. Y Acad. Sci. 2004, 1036, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Huizinga, D.; Haberstick, B.C.; Smolen, A.; Menard, S.; Young, S.E.; Corley, R.P.; Stalling, M.C.; Grotpeter, J.; Hewitt, J.K. Childhood maltreatment, subsequent antisocial behavior, and the role of monoamine oxidase A genotype. Biol. Psychiatry 2006, 60, 677–683. [Google Scholar] [CrossRef]
- Sjöberg, R.L.; Ducci, F.; Barr, C.S.; Newman, T.K.; Dell’osso, L.; Virkkunen, M.; Goldman, D.A. Non-additive interaction of a functional MAO-A VNTR and testosterone predicts antisocial behavior. Neuropsychopharmacology 2008, 33, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.S.; Mohyuddin, A.; Colonna, V.; Mehdi, S.Q.; Ayub, Q. Monoamine Oxidase A gene polymorphisms and self reported aggressive behaviour in a Pakistani ethnic group. J. Pak. Med. Assoc. 2015, 65, 818–824. [Google Scholar] [PubMed]
- Eun, T.K.; Jeong, S.H.; Lee, K.Y.; Kim, S.H.; Ahn, Y.M.; Bang, Y.W.; Joo, E.J. Association between the 5-HTTLPR Genotype and Childhood Characteristics in Mood Disorders. Clin. Psychopharmacol. Neurosci. 2016, 14, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Twitchell, G.R.; Hanna, G.L.; Cook, E.H.; Stoltenberg, S.F.; Fitzgerald, H.E.; Zucker, R.A. Serotonin transporter promoter polymorphism genotype is associated with behavioral disinhibition and negative affect in children of alcoholics. Alcohol. Clin. Exp. Res. 2001, 25, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Zalsman, G.; Frisch, A.; Bromberg, M.; Gelernter, J.; Michaelovsky, E.; Campino, A.; Erlich, Z.; Tyano, S.; Apter, A.; Weizman, A. Family-based association study of serotonin transporter promoter in suicidal adolescents: No association with suicidality but possible role in violence traits. Am. J. Med. Genet. 2001, 105, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Liao, D.L.; Hong, C.J.; Shih, H.L.; Tsai, S.J. Possible association between serotonin transporter promoter region polymorphism and extremely violent crime in Chinese males. Biol. Psychiatry 2004, 50, 284–287. [Google Scholar] [CrossRef]
- Gerra, G.; Garofano, L.; Santoro, G.; Bosari, S.; Pellegrini, C.; Zaimovic, A.; Moi, G.; Bussandri, M.; Moi, A.; Brambilla, F.; et al. Association between low-activity serotonin transporter genotype and heroin dependence: Behavioral and personality correlates. Am. J. Med. Genet. (Neuropsychiatr. Genet.) Part B 2004, 126B, 37–42. [Google Scholar] [CrossRef]
- Rossi, R.; Socci, V.; Gregori, E.; Talevi, D.; Collazzoni, A.; Pacitti, F.; Stratta, P.; Rossi, A.; Di Lorenzo, G. ResilienCity: Resilience and Psychotic-Like Experiences 10 Years After L’Aquila Earthquake. Front. Psychiatry 2020, 11, 77–83. [Google Scholar] [CrossRef] [PubMed]
- da Cunha-Bang, S.; Knudsen, G.M. The Modulatory Role of Serotonin on Human Impulsive Aggression. Biol. Psychiatry 2021, 90, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Kuzhiyengal Mambra, A.J.; Kotian, S. Understanding Aggressive Behavior: A Comprehensive Review of Research. IJNRD 2023, 8, 144–149. [Google Scholar]
- Zhen, B.; Yao, B.; Zhou, X. Pandemic Exposure, Post-traumatic Stress Disorder, Conflict Behaviors, and Online Aggressive Behaviors among College Students during the COVID-19 Pandemic: Examining the Moderating Role of Gender. Front. Psychiatry 2022, 13, 809173–809180. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Leon, S.; Wegman-Ostrosky, T.; Perelman, C.; Sepulveda, R.; Rebolledo, P.A.; Cuapio, A.; Villapol, S. More than 50 long-term effects of COVID-19: A systematic review and meta-analysis. Sci. Rep. 2021, 11, 16144–16156. [Google Scholar] [CrossRef] [PubMed]
- Boccia, S.; Ricciardi, W.; Ioannidis, J.P.A. What Other Countries Can Learn From Italy During the COVID-19 Pandemic. JAMA Intern. Med. 2020, 180, 927–928. [Google Scholar] [CrossRef] [PubMed]
- Giallonardo, V.; Sampogna, G.; Del Vecchio, V.; Luciano, M.; Albert, U.; Carmassi, C.; Carrà, G.; Cirulli, F.; Dell’Osso, B.; Nanni, M.G.; et al. The Impact of Quarantine and Physical Distancing Following COVID-19 on Mental Health: Study Protocol of a Multicentric Italian Population Trial. Front. Psychiatry 2020, 11, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Demontis, D.; Walters, R.K.; Rajagopal, V.M.; Waldman, I.D.; Grove, J.; Als, T.D.; Dalsgaard, S.; Ribases, M.; Bybjerg Grauholm, J.; Baekvad Hansen, M.; et al. Author Correction: Risk variants and polygenic architecture of disruptive behavior disorders in the context of attention-deficit/hyperactivity disorder. Nat. Commun. 2021, 12, 1166–1177. [Google Scholar] [CrossRef]
- Seo, D.; Patrick, C.J.; Kennealy, P.J. Role of Serotonin and Dopamine System Interactions in the Neurobiology of Impulsive Aggression and its Comorbidity with other Clinical Disorders. Aggress. Violent Behav. 2008, 13, 383–395. [Google Scholar] [CrossRef]
- Narvaes, R.; Martins de Almeida, R.M. Aggressive behavior and three neurotransmitters: Dopamine, GABA, and serotonin-A review of the last 10 years. Psychol. Neurosci. 2014, 7, 601–607. [Google Scholar] [CrossRef]
- Palumbo, S.; Mariotti, V.; Iofrida, C.; Pellegrini, S. Genes and Aggressive Behavior: Epigenetic Mechanisms Underlying Individual Susceptibility to Aversive Environments. Front. Behav. Neurosci. 2018, 12, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Godar, S.C.; Fite, P.J.; McFarlin, K.M.; Bortolato, M. The role of monoamine oxidase A in aggression: Current translational developments and future challenges. Prog. Neuropsychopharmacol. Biol. Psychiatry. 2016, 69, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Popova, N.K. From genes to aggressive behavior: The role of serotonergic system. Bioessays 2006, 28, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Hornung, J.P. The human raphe nuclei and the serotonergic system. J. Chem. Neuroanat. 2003, 26, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, L.; Goldman, D. Genetics of impulsive behaviour. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 25, 1615–1627. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Quadros, I.M.; de Almeida, R.M.M.; Miczek, K.A. Brain serotonin receptors and transporters: Initiation vs. termination of escalated aggression. Psychopharmacology 2011, 213, 183–212. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, M.; Ricci, L.A.; Coppersmith, G.A.; Melloni, R.H. The effect of increased serotonergic neurotransmission on aggression: A critical meta-analytical review of preclinical studies. Psychopharmacology 2009, 205, 349–368. [Google Scholar] [CrossRef]
- Mentis, A.F.A.; Dardiotis, E.; Katsouni, E.; Chrousos, G.P. Fromwarrior genesto translational solutions: Novel insights into monoamine oxidases (MAOs) and aggression. Transl. Psychiatry 2021, 11, 130–140. [Google Scholar] [CrossRef] [PubMed]
- van Rhijn, J.R.; Shi, Y.; Bormann, M.; Mossink, B.; Frega, M.; Recaioglu, H.; Hakobjan, M.; Gunnewiek, T.K.; Schoenmaker, C.; Palmer, E.; et al. Brunner syndrome associated MAOA mutations result in NMDAR hyperfunction and increased network activity in human dopaminergic neurons. Neurobiol. Dis. 2022, 163, 105587–105600. [Google Scholar] [CrossRef]
- Hotamisligil, G.S.; Breakefield, X.O. Human monoamine oxidase A gene determines levels of enzyme activity. Am. J. Hum. Genet. 1991, 49, 383–392. [Google Scholar]
- Wang, M.; Li, H.; Deater-Deckard, K.; Zhang, W. Interacting Effect of Catechol-O-Methyltransferase (COMT) and Monoamine Oxidase A (MAOA) Gene Polymorphisms, and Stressful Life Events on Aggressive Behavior in Chinese Male Adolescents. Front. Psychol. 2018, 9, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Antypa, N.; Giegling, I.; Calati, R.; Schneider, B.; Hartmann, A.M.; Friedl, M.; Konte, B.; Lia, L.; De Ronchi, D.; Serretti, A.; et al. MAOA and MAOB polymorphisms and anger-related traits in suicidal participants and controls. Eur. Arch. Psychiatry Clin. Neurosci. 2013, 263, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Lundwall, R.A.; Rasmussen, C.G. MAOA Influences the Trajectory of Attentional Development. Front. Hum. Neurosci. 2016, 10, 424–439. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Cao, Y.; Zhang, L.; Zhang, W. Parenting Practices and Adolescent Effortful Control:MAOAT941G Gene Polymorphism as a Moderator. Front. Psychol. 2020, 11, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W. Genetic and environmental influences on behavior: Capturing all the interplay. Psycholog. Rev. 2007, 114, 423–440. [Google Scholar] [CrossRef] [PubMed]
- Passamonti, L.; Cerasa, A.; Gioia, M.C.; Magariello, A.; Muglia, M.; Quattrone, A.; Fera, F. Genetically dependent modulation of serotonergic inactivation in the human prefrontal cortex. Neuroimage 2008, 40, 1264–1273. [Google Scholar] [CrossRef] [PubMed]
- Slaby, I.; Hain, H.S.; Abrams, D.; Mentch, F.D.; Glessner, J.T.; Sleiman, P.M.A.; Hakonarson, H. An electronic health record (EHR) phenotype algorithm to identify patients with attention deficit hyperactivity disorders (ADHD) and psychiatric comorbidities. J. Neurodev. Disord. 2022, 14, 37–46. [Google Scholar] [CrossRef]
- Hernández-Andrade, L.; Hermosillo-Abundis, A.C.; Betancourt-Navarrete, B.L.; Ruge, D.; Trenado, C.; Lemuz-López, R.; Pelayo-González, H.J.; López-Cortés, V.A.; Bonilla-Sánchez, M.D.R.; García-Flores, M.A.; et al. EEG Global Coherence in Scholar ADHD Children during Visual Object Processing. Int. J. Environ. Res. Public Health 2022, 19, 5953. [Google Scholar] [CrossRef]


{kind=link}
| Age (Mean ± SD) | 10.8 ± 0.84 |
| Sex (n, %) | |
| Female | 6 (7.5%) |
| Male | 74 (92.5%) |
| MAOA rs6323F | 5′-TAATTAATGCGATCCCTCCG-3′ |
| MAOArs6323 R | 5′-TGAGGAAATTGACAGACCAAGA-3′ |
| MAOA rs1137070F | 5′-GGCAACGTTTTTGGCATCTGGTC-3′ |
| MAOArs1137070 R | 5′-ACTCATGCTGACAAGGAGGAACA-3′ |
| ADHD | Controls | p | OR | |||
|---|---|---|---|---|---|---|
| MAOA SNP rs6323 (G891T) | ||||||
| Genotypes | n = 80 | % | n = 80 | % | ||
| TT | 56 | 0.70 | 51 | 0.64 | ns § | - |
| GT | 4 | 0.05 | 21 | 0.26 | 0.0008 | 0.1479 |
| GG | 20 | 0.25 | 8 | 0.10 | 0.02 | 3.0 |
| Alleles | 2n = 160 | % | 2n = 160 | % | ||
| T | 116 | 0.73 | 123 | 0.77 | ns § | - |
| G | 44 | 0.27 | 37 | 0.23 | ns § | - |
| MAOA SNP rs1137070 (T1410C) | ||||||
| Genotypes | n = 80 | % | n = 81 * | % | ||
| CC | 56 | 0.70 | 51 | 0.63 | ns § | - |
| CT | 5 | 0.06 | 21 | 0.26 | 0.0006 | 0.19 |
| TT | 19 | 0.24 | 9 | 0.11 | 0.0344 | 2.49 |
| Alleles | 2n = 160 | % | 2n = 162 | % | ||
| C | 117 | 0.73 | 123 | 0.76 | ns § | - |
| T | 43 | 0.27 | 39 | 0.24 | ns § | - |
| Aggress. ADHD | Non-Aggress. ADHD | Controls | ||||
|---|---|---|---|---|---|---|
| MAOA SNP rs6323 (G891T) | ||||||
| Genotypes | n = 32 | % | n = 48 | % | n = 80 | % |
| TT | 22 | 0.70 | 34 | 0.71 | 51 | 0.64 |
| GT | 3 | 0.10 | 1 | 0.02 | 21 | 0.26 |
| GG | 7 | 0.20 | 13 | 0.27 | 8 | 0.10 |
| Alleles | 2n = 64 | % | 2n = 96 | % | 2n = 160 | % |
| T | 47 | 0.74 | 69 | 0.77 | 123 | 0.77 |
| G | 17 | 0.26 | 27 | 0.23 | 37 | 0.23 |
| MAOA SNP rs1137070 (T1410C) | ||||||
| Genotypes | n = 32 | % | n = 48 | % | n = 81 * | % |
| CC | 22 | 0.70 | 34 | 0.71 | 51 | 0.63 |
| CT | 3 | 0.10 | 3 | 0.06 | 21 | 0.26 |
| TT | 7 | 0.20 | 11 | 0.23 | 9 | 0.11 |
| Alleles | 2n = 64 | % | 2n = 96 | % | 2n = 162 | % |
| C | 47 | 0.73 | 71 | 0.74 | 123 | 0.76 |
| T | 17 | 0.27 | 25 | 0.26 | 39 | 0.24 |
| Aggressive ADHD | Non-Aggressive ADHD | |||
|---|---|---|---|---|
| 5HTT SNP | ||||
| rs4795541(S)/(L) Polymorphism | ||||
| Genotypes | n = 32 | % | n = 48 | % |
| LL | 11 | 0.70 | 11 | 0.23 |
| LS | 16 | 0.10 | 28 | 0.58 |
| SS | 5 | 0.20 | 9 | 0.18 |
| Alleles | 2n = 64 | % | 2n = 96 | % |
| L | 38 | 0.60 | 50 | 0.52 |
| S | 26 | 0.40 | 46 | 0.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neri, L.; Marziani, B.; Sebastiani, P.; Del Beato, T.; Colanardi, A.; Legge, M.P.; Aureli, A. Aggressiveness in Italian Children with ADHD: MAOA Gene Polymorphism Involvement. Diseases 2024, 12, 70. https://doi.org/10.3390/diseases12040070
Neri L, Marziani B, Sebastiani P, Del Beato T, Colanardi A, Legge MP, Aureli A. Aggressiveness in Italian Children with ADHD: MAOA Gene Polymorphism Involvement. Diseases. 2024; 12(4):70. https://doi.org/10.3390/diseases12040070
Chicago/Turabian StyleNeri, Ludovico, Beatrice Marziani, Pierluigi Sebastiani, Tiziana Del Beato, Alessia Colanardi, Maria Pia Legge, and Anna Aureli. 2024. "Aggressiveness in Italian Children with ADHD: MAOA Gene Polymorphism Involvement" Diseases 12, no. 4: 70. https://doi.org/10.3390/diseases12040070
APA StyleNeri, L., Marziani, B., Sebastiani, P., Del Beato, T., Colanardi, A., Legge, M. P., & Aureli, A. (2024). Aggressiveness in Italian Children with ADHD: MAOA Gene Polymorphism Involvement. Diseases, 12(4), 70. https://doi.org/10.3390/diseases12040070