Keratoacanthoma Pathobiology in Mouse Models

Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
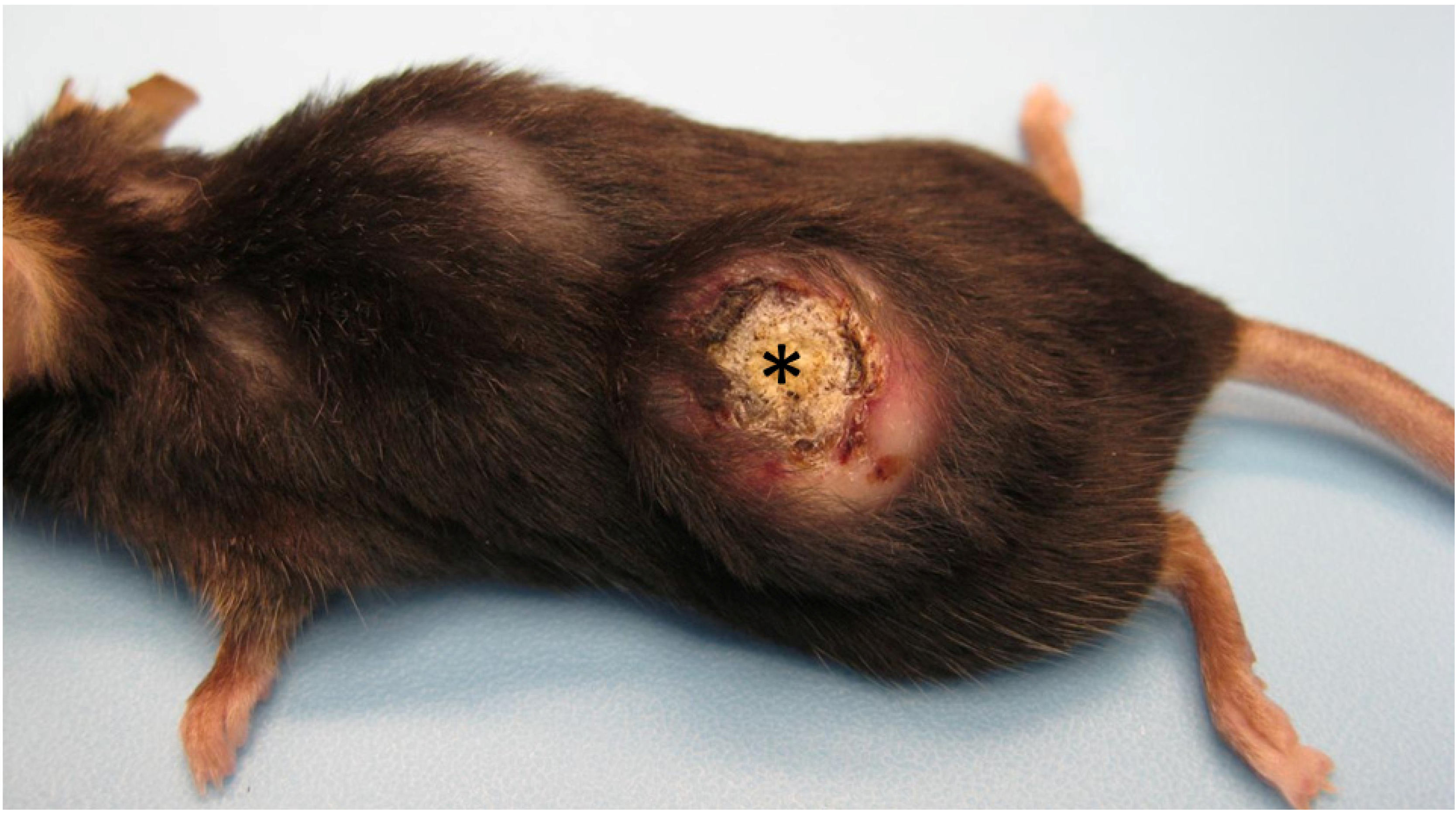
2.2. Clinical Tumor Distribution

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mouse | Neck | Head | Ear | Chest | Back | Abdomen | Side | Limbs/paws | Tail | Perineal | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Zimz1 | 1% (1) | 20% (18) | 11% (10) | 8% (7) | 28% (25) | 2% (2) | 2% (2) | 21% (19) | 5% (5) | 2% (2) | 90 |
| RAG-SB | 8% (4) | 8% (4) | 2% (1) | 30% (14) | 23% (11) | 8% (4) | 10% (5) | 8% (4) | 0% (0) | 0% (0) | 47 |
2.3. Histopathology
2.4. Histologic grading
| KA Phase | Morphologic features | Transition to next phase |
|---|---|---|
| Growth |
|
|
| Maturation |
|
|
| Regression |
|
2.5. Immunohistochemistry
| Marker | Antibody | Dilution | Source | Conditions |
|---|---|---|---|---|
| CD3 | Cat# RM-9107-5 | 1:200 | Neomarkers | HIER, citrate buffer (pH 6.0) |
| B220 | Cat# MCA1258G | 1:6000 | Serotec | HIER, citrate buffer (pH 6.0) |
| F4/80 | Cat# MCAP497 | 1:6400 | Serotec | HIER, citrate buffer (pH 6.0) [11] |
| Arginase-1 | Cat# sc20150 | 1:300 | Santa Cruz | HIER, citrate buffer (pH 6.0) [12] |
| Nos2 | Cat# sc649 | 1:200 | Santa Cruz | HIER, citrate buffer (pH 6.0) |
| Granzyme B | Cat# ab4059 | 1:200 | Abcam | HIER, citrate buffer (pH 6.0) |
3. Results
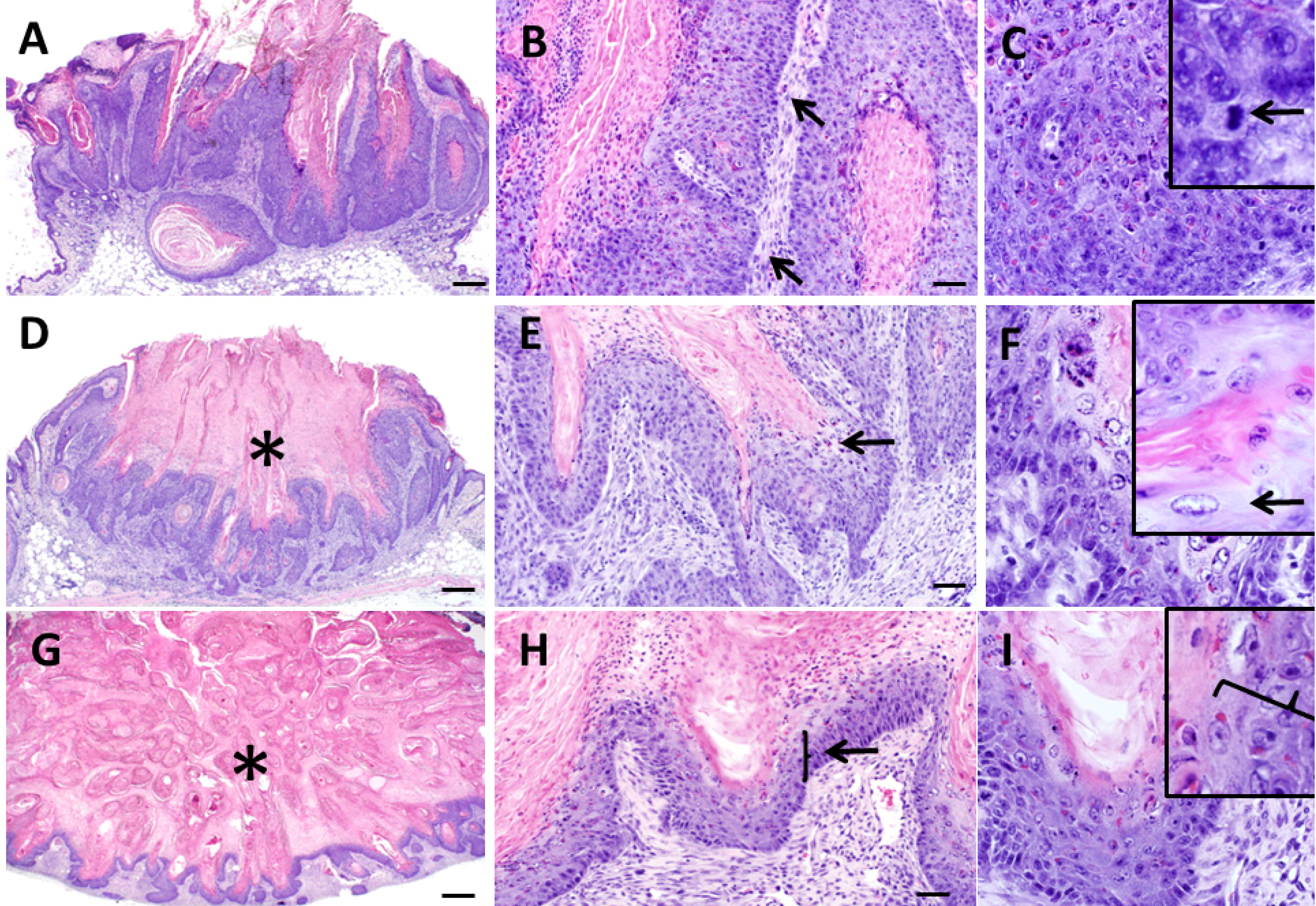
3.1. Pathological Appearance of Murine Keratoacanthomas
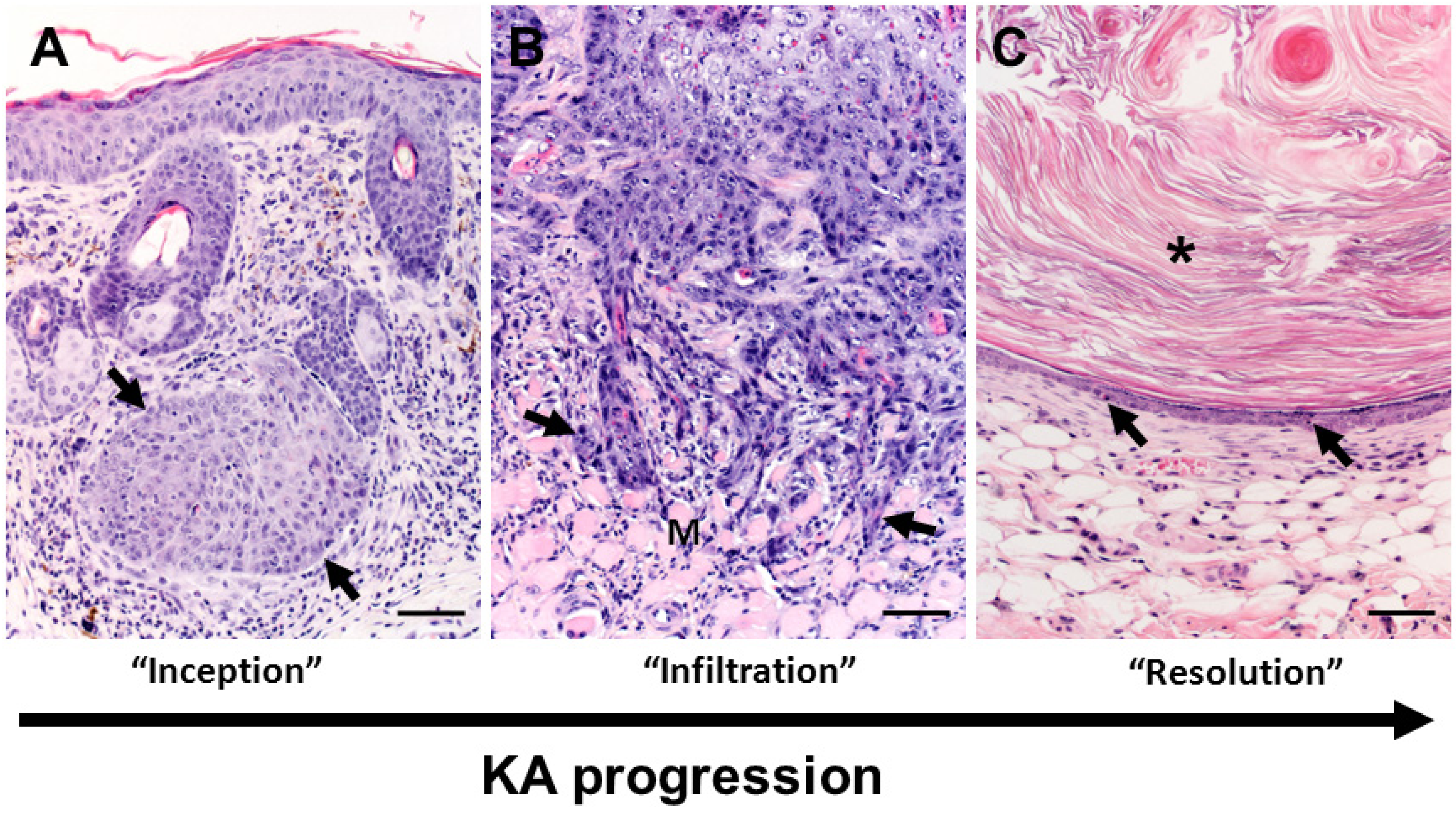
3.2. Growth Phase



3.3. Maturation Phase

3.4. Regression Phase
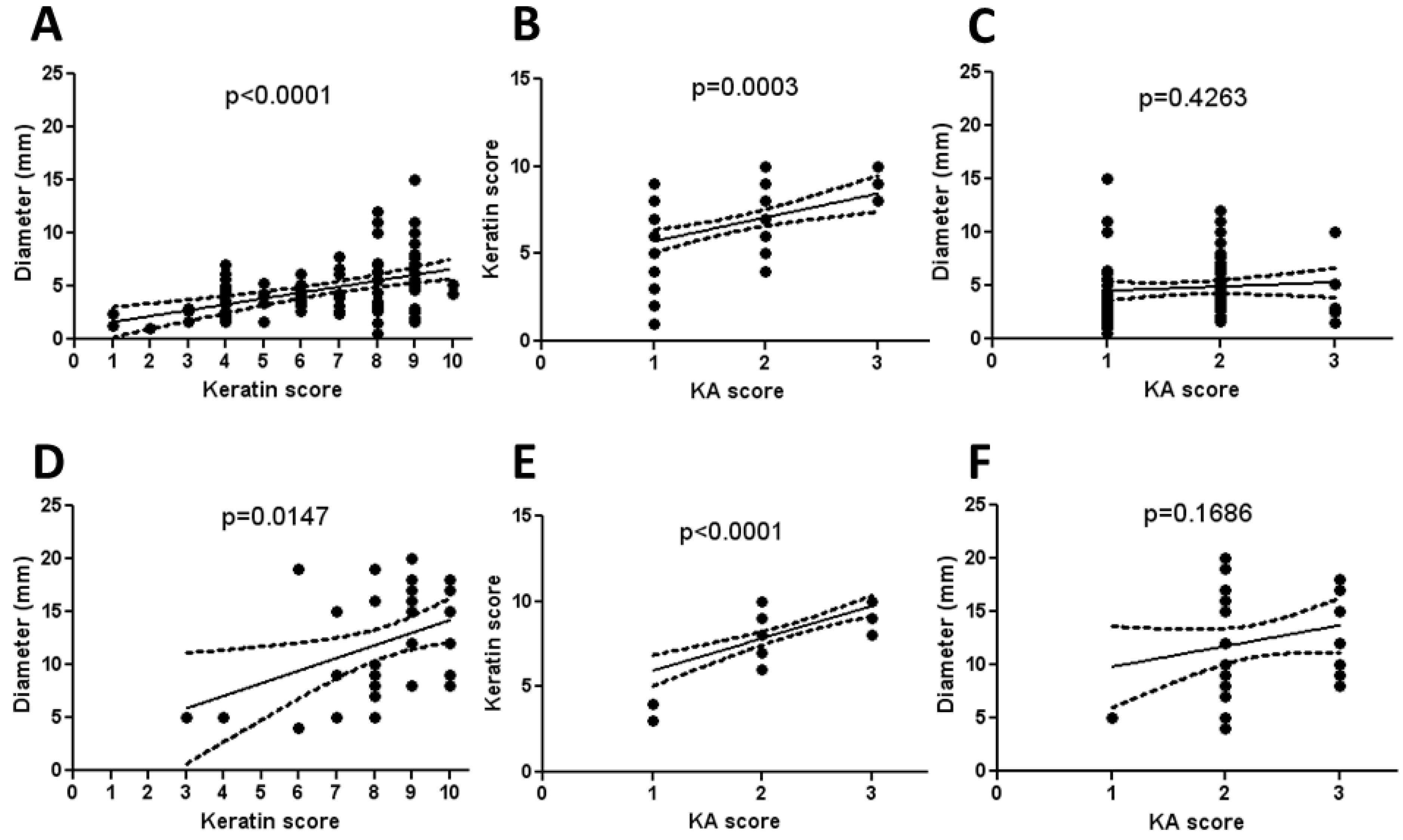
3.5. Histologic Classification of Murine Keratoacanthomas

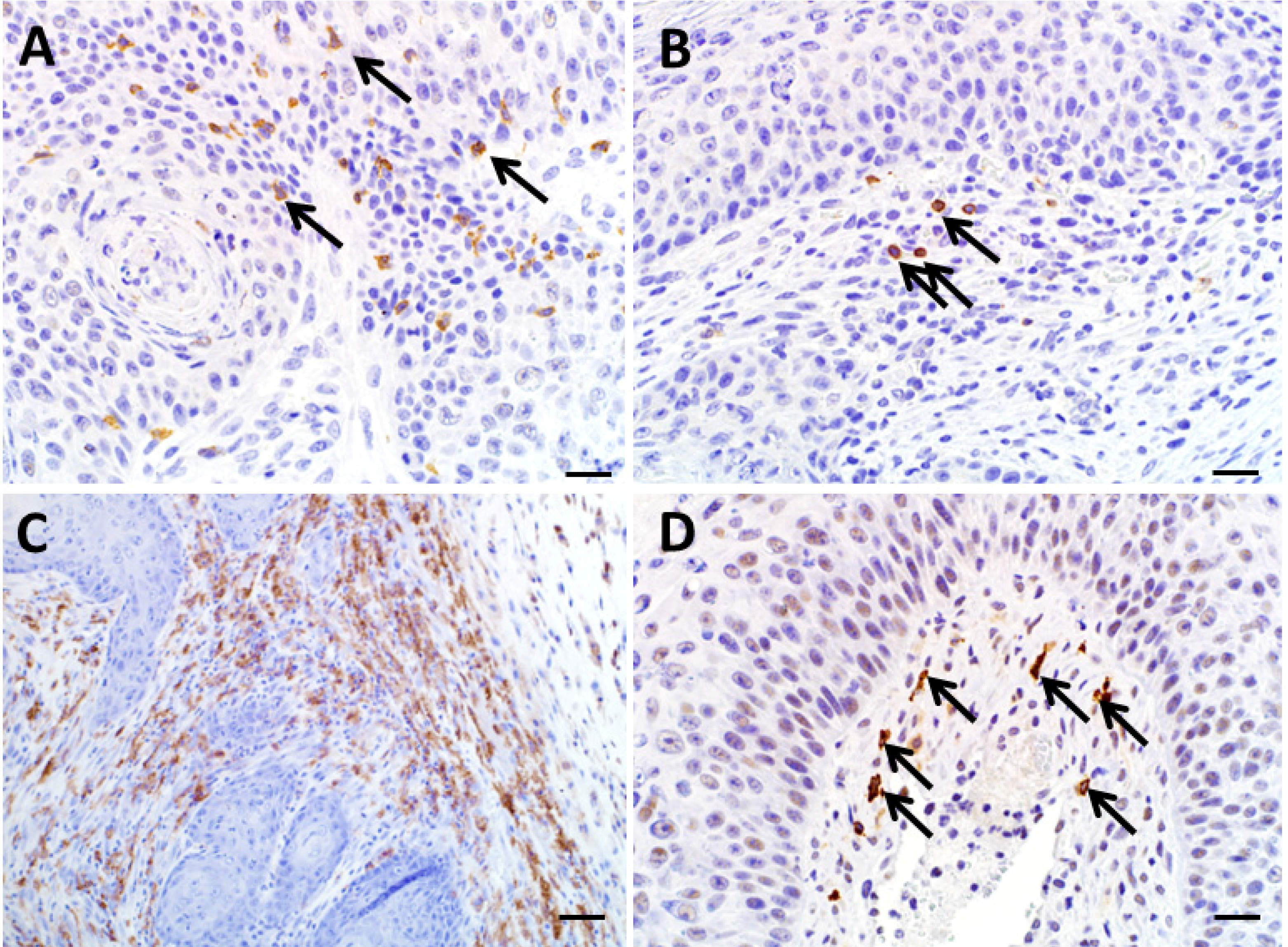
3.6. Utility of Immunohistochemistry to Assess Immune Cell Infiltration of Regressing Murine Keratoacanthomas

4. Discussion
5. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Ko, C.J. Keratoacanthoma: Facts and controversies. Clin. Dermatol. 2010, 28, 254–261. [Google Scholar] [CrossRef]
- Cardiff, R.D. The Tumor Pathology of Genetically Engineered Mice: Genomic Pathology. Genet. Eng. Mice Cancer Res. 2012, 133–180. [Google Scholar] [CrossRef]
- Kossard, S.; Tan, K.B.; Choy, C. Keratoacanthoma and infundibulocystic squamous cell carcinoma. Am. J. Dermatopathol. 2008, 30, 127–134. [Google Scholar] [CrossRef]
- Mandrell, J.C.; Santa Cruz, D. Keratoacanthoma: Hyperplasia, benign neoplasm, or a type of squamous cell carcinoma? Semi. Diagn. Pathol. 2009, 26, 150–163. [Google Scholar] [CrossRef]
- Schwartz, R.A.; Tarlow, M.M.; Lambert, W.C. Keratoacanthoma-like squamous cell carcinoma within the fibroepithelial polyp. Dermatol. Surg. 2004, 30, 349–350. [Google Scholar] [CrossRef]
- Rinker, M.H.; Fenske, N.A.; Scalf, L.A.; Glass, L.F. Histologic variants of squamous cell carcinoma of the skin. Cancer Control 2001, 8, 354–363. [Google Scholar]
- Karaa, A.; Khachemoune, A. Keratoacanthoma: A tumor in search of a classification. Int. J. Dermatol. 2007, 46, 671–678. [Google Scholar] [CrossRef]
- Rogers, L.M.; Riordan, J.D.; Swick, B.L.; Meyerholz, D.K.; Dupuy, A.J. Ectopic expression of zmiz1 induces cutaneous squamous cell malignancies in a mouse model of cancer. J. Invest. Dermatol. 2013, 133, 1863–1869. [Google Scholar] [CrossRef]
- Rogers, L.M.; Olivier, A.K.; Meyerholz, D.K.; Dupuy, A.J. Adaptive immunity does not strongly suppress spontaneous tumors in a sleeping beauty model of cancer. J. Immunol. 2013, 190, 4393–4399. [Google Scholar] [CrossRef]
- Gibson-Corley, K.N.; Olivier, A.K.; Meyerholz, D.K. Principles for valid histopathologic scoring in research. Vet. Pathol. 2013, 50, 1007–1015. [Google Scholar] [CrossRef]
- Meyerholz, D.K.; Williard, D.E.; Grittmann, A.M.; Samuel, I. Murine pancreatic duct ligation induces stress kinase activation, acute pancreatitis, and acute lung injury. Am. J. Surg. 2008, 196, 675–682. [Google Scholar] [CrossRef]
- Hochstedler, C.M.; Leidinger, M.R.; Maher-Sturm, M.T.; Gibson-Corley, K.N.; Meyerholz, D.K. Immunohistochemical detection of arginase-i expression in formalin-fixed lung and other tissues. J. Histotechnol. 2013, 36, 128–134. [Google Scholar] [CrossRef]
- Shinkai, Y.; Rathbun, G.; Lam, K.P.; Oltz, E.M.; Stewart, V.; Mendelsohn, M.; Charron, J.; Datta, M.; Young, F.; Stall, A.M.; et al. Rag-2-deficient mice lack mature lymphocytes owing to inability to initiate v(d)j rearrangement. Cell 1992, 68, 855–867. [Google Scholar] [CrossRef]
- Schwartz, R.A. Keratoacanthoma: A clinico-pathologic enigma. Dermatol. Surg. 2004, 30, 326–333, discussion 333. [Google Scholar] [CrossRef]
- Brennan, P.A.; Umar, T.; Smith, G.I.; Lo, C.H.; Tant, S. Expression of nitric oxide synthase-2 in cutaneous squamous cell carcinoma of the head and neck. Br. J. Oral Maxillofac. Surg. 2002, 40, 191–194. [Google Scholar] [CrossRef]
- Ghadially, F.N. A comparative morphological study of the kerato-acanthoma of man and similar experimentally produced lesions in the rabbit. J. Pathol. Bacteriol. 1958, 75, 441–453. [Google Scholar] [CrossRef]
- Misago N, T.T.; Toda S, Narisawa Y. The changes in the expression levels of follicular markers in keratoacanthoma depend on the stage: Keratoacanthoma is a follicular neoplasm exhibiting infundibular/isthmic differentiation without expression of ck15. J. Cutan. Pathol. 2014. [Google Scholar] [CrossRef]
- Batinac, T.; Zamolo, G.; Hadzisejdic, I.; Zauhar, G. A comparative study of granzyme b expression in keratoacanthoma and squamous cell carcinoma. J. Dermatol. Sci. 2006, 44, 109–112. [Google Scholar] [CrossRef]
- Peng, X.; Griffith, J.W.; Han, R.; Lang, C.M.; Kreider, J.W. Development of keratoacanthomas and squamous cell carcinomas in transgenic rabbits with targeted expression of ejras oncogene in epidermis. Am. J. Pathol. 1999, 155, 315–324. [Google Scholar] [CrossRef]
- Ramselaar, C.G.; Ruitenberg, E.J.; Kruizinga, W. Regression of induced keratoacanthomas in anagen (hair-growth phase) skin-grafts in mice. Cancer Res. 1980, 40, 1668–1673. [Google Scholar]
- Li, J.; Wang, K.; Gao, F.; Jensen, T.D.; Li, S.T.; DeAngelis, P.M.; Kolvraa, S.; Proby, C.; Forslund, O.; Bolund, L.; et al. Array comparative genomic hybridization of keratoacanthomas and squamous cell carcinomas: Different patterns of genetic aberrations suggest two distinct entities. J. Invest. Dermatol. 2012, 132, 2060–2066. [Google Scholar] [CrossRef]
- Sundberg, J.P.; Sundberg, B.A.; Beamer, W.G. Comparison of chemical carcinogen skin tumor induction efficacy in inbred, mutant, and hybrid strains of mice: Morphologic variations of induced tumors and absence of a papillomavirus cocarcinogen. Mol. Carcinog. 1997, 20, 19–32. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gibson-Corley, K.N.; Rogers, L.M.; Goeken, A.; Dupuy, A.J.; Meyerholz, D.K. Keratoacanthoma Pathobiology in Mouse Models. Diseases 2014, 2, 106-119. https://doi.org/10.3390/diseases2020106
Gibson-Corley KN, Rogers LM, Goeken A, Dupuy AJ, Meyerholz DK. Keratoacanthoma Pathobiology in Mouse Models. Diseases. 2014; 2(2):106-119. https://doi.org/10.3390/diseases2020106
Chicago/Turabian StyleGibson-Corley, Katherine N., Laura M. Rogers, Adam Goeken, Adam J. Dupuy, and David K. Meyerholz. 2014. "Keratoacanthoma Pathobiology in Mouse Models" Diseases 2, no. 2: 106-119. https://doi.org/10.3390/diseases2020106