Mechanism of Fibrosis Induced by Echinococcus spp.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methodology
3. Mechanisms of Hepatic Fibrosis Induced by Echinococcus
3.1. MiRNAs Play an Important Role in the Hepatic Fibrosis Induced by Echinococcus
3.2. TGF-β Plays an Important Role in the Hepatic Fibrosis Induced by Echinococcus
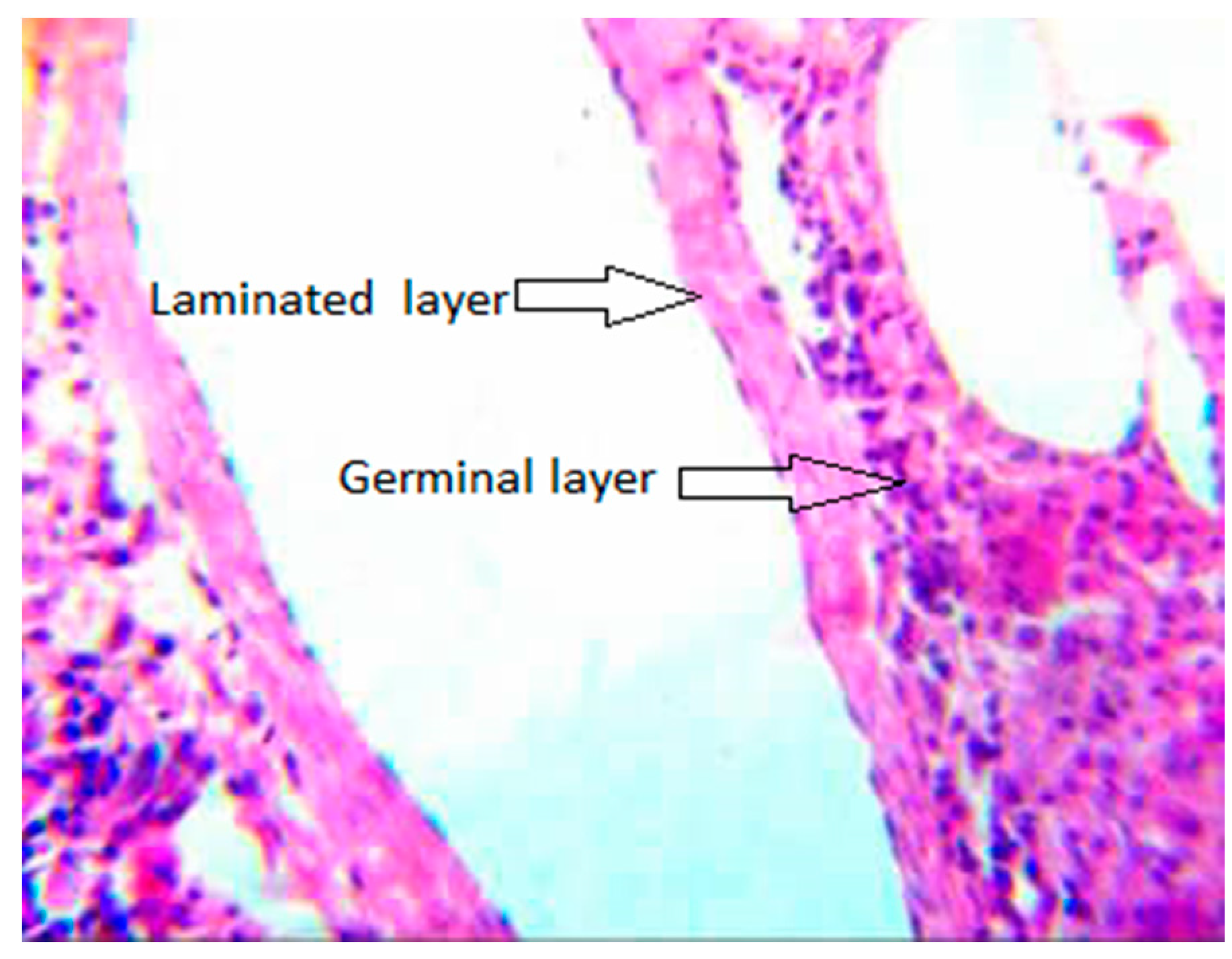
3.3. Collagen Fiber around the Metacestodes of E. granulosus
3.4. Other Hepatic Fibrosis Factors Induced by Echinococcus
4. Fibrosis in Other Organs Induced by Echinococcus
5. Therapeutics
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gao, H. The Study of Relation between Calcium/Angiogeesis Signal Pathway and Growth Status of Hydatid. Master’s Thesis, Lanzhou University, Lanzhou, China, June 2018. [Google Scholar]
- Eckert, J.; Gemmell, M.A.; Meslin, F.X.; Pawlowski, Z.S. WHO/OIE manual on echinococcosis in humans and animals: A public health problem of global concern. Int. J. Parasitol. 2001, 31, 1717–1718. [Google Scholar]
- Nakao, M.; Lavikainen, A.; Yanagida, T.; Ito, A. Phylogenetic systematics of the genus Echinococcus (Cestoda: Taeniidae). Int. J. Parasitol. 2013, 43, 1017–1029. [Google Scholar] [CrossRef] [PubMed]
- Xiao, N.; Qiu, J.M.; NaKao, M.; Li, T.Y.; Chen, X.W.; Schantz, P.M.; Craig, P.S.; Ito, A. Biological Features of a New Echinococcus Species (Echinococcus shiquicus) in the East of Qinghai-Tibet Plateau. Chin. J. Parasitol. Parasit. Dis. 2008, 26, 307–312. [Google Scholar]
- Moro, P.; Schantz, P. Echinococcosis: A review. Int. J. Infect. Dis. 2009, 13, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, E.; Garcia, H.H.; Junghanss, T. Cystic echinococcosis: Chronic, complex, and still neglected. PLoS Negl. Trop. Dis. 2011, 5, e1146. [Google Scholar] [CrossRef] [PubMed]
- Wahlstrom, H.; Isomursu, M.; Hallgren, G.; Christensson, D.; Cedersmyg, M.; Wallensten, A.; Hjertqvist, M.; Davidson, R.K.; Uhlhorn, H.; Hopp, P. Combining information from surveys of several species to estimate the probability of freedom from Echinococcus multilocularis in Sweden, Finland and mainland Norway. Acta Vet. Scand. 2011, 53, 9. [Google Scholar] [CrossRef] [PubMed]
- Brehm, K.; Koziol, U. Echinococcus-Host Interactions at Cellular and Molecular Levels. Adv. Parasitol. 2017, 95, 147–212. [Google Scholar] [CrossRef] [PubMed]
- Craig, P.S.; Mcmanus, D.P.; Lightowlers, M.W.; Chabalgoity, J.A.; Garcia, H.H.; Gavidia, C.M.; Gilman, R.H.; Gonzalez, A.E.; Lorca, M.; Naquira, C. Prevention and control of cystic echinococcosis. Lancet Infect. Dis. 2007, 7, 385–394. [Google Scholar] [CrossRef]
- Brunetti, E.; Kern, P.; Vuitton, D.A. Expert consensus for the diagnosis and treatment of cystic and alveolar echinococcosis in humans. Acta Trop. 2010, 114, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Tong, S.; Jing, T.; Chong, S.; Cai, X.; Jing, Z.; Han, J. Investigation on Echinococcosis in Animals in Gannan Tibetan Autonomous Prefecture. Chin. J. Parasitol. Parasit. Dis. 2009, 27, 27–30. [Google Scholar]
- Lu, Y.; Huang, Q.; Zhou, Z.; Lu, Y. An overview of parasitic diseases by analyzing publications at National Institue of Parasitic Diseases Control and Prevention in the last five years. Chin. J. Dis. Control Prev. 2015, 19, 86–90. [Google Scholar]
- Ito, A.; Urbani, C.; Qiu, J.; Vuitton, D.A.; Qiu, D.; Heath, D.D.; Craig, P.S.; Zheng, F.; Schantz, P.M. Control of echinococcosis and cysticercosis: A public health challenge to international cooperation in China. Acta Trop. 2003, 86, 3–17. [Google Scholar] [CrossRef]
- Budke, C.M.; Deplazes, P.; Torgerson, P.R. Global Socioeconomic Impact of Cystic Echinococcosis. Emerg. Infect. Dis. 2006, 12, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Lee, U.E.; Friedman, S.L. Mechanisms of Hepatic Fibrogenesis. Best Pract. Res. Clin. Gastroenterol. 2011, 25, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Vuitton, D.A.; Guerret-Stocker, S.; Carbillet, J.P.; Mantion, G.; Miguet, J.P.; Grimaud, J.A. Collagen immunotyping of the hepatic fibrosis in human alveolar echinococcosis. Z. Parasitenkd. 1986, 72, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Boldin, M.P.; Baltimore, D. MicroRNAs, new effectors and regulators of NF-κB. Immunol. Rev. 2012, 246, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Kwiecinski, M.; Noetel, A.; Elfimova, N.; Trebicka, J.; Schievenbusch, S.; Strack, I.; Molnar, L.; Von, B.M.; Töx, U.; Nischt, R. Hepatocyte growth factor (HGF) inhibits collagen I and IV synthesis in hepatic stellate cells by miRNA-29 induction. PLoS ONE 2011, 6, e24568. [Google Scholar] [CrossRef] [PubMed]
- Lakner, A.M.; Steuerwald, N.M.; Walling, T.L.; Ghosh, S.; Li, T.; Mckillop, I.H.; Russo, M.W.; Bonkovsky, H.L.; Schrum, L.W. Inhibitory effects of microRNA 19b in hepatic stellate cell-mediated fibrogenesis. Hepatology 2012, 56, 300–310. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Wu, C.Q.; Zhang, Z.Q.; Yao, D.K.; Zhu, L. Loss of expression of miR-335 is implicated in hepatic stellate cell migration and activation. Exp. Cell Res. 2011, 317, 1714–1725. [Google Scholar] [CrossRef]
- Guo, C.J.; Pan, Q.; Jiang, B.; Chen, G.Y.; Li, D.G. Effects of upregulated expression of microRNA-16 on biological properties of culture-activated hepatic stellate cells. Apoptosis 2009, 14, 1331. [Google Scholar] [CrossRef] [PubMed]
- Sekiya, Y.; Ogawa, T.; Iizuka, M.; Yoshizato, K.; Ikeda, K.; Kawada, N. Down-regulation of cyclin E1 expression by microRNA-195 accounts for interferon-β-induced inhibition of hepatic stellate cell proliferation. J. Cell. Physiol. 2011, 226, 2535–2542. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Li, W.; Guo, K.; Xiao, Y.; Wang, Y.; Fan, J. miR-181b promotes hepatic stellate cells proliferation by targeting p27 and is elevated in the serum of cirrhosis patients. Biochem. Biophys. Res. Commun. 2012, 421, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Sekiya, Y.; Ogawa, T.; Yoshizato, K.; Ikeda, K.; Kawada, N. Suppression of hepatic stellate cell activation by microRNA-29b. Biochem. Biophys. Res. Commun. 2011, 412, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Lin, Z.; Dong, P.; Lu, Z.; Gao, S.; Chen, X.; Wu, C.; Yu, F. Activation of hepatic stellate cells is suppressed by microRNA-150. Int. J. Mol. Med. 2013, 32, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, X. miR-133a Effect Study on Echinococcus multilocularis Induced Hepatic Stellate Cell Activation In Vitro and Liver Fibrosis in Infected Mice In Vivo. Master’s Thesis, Xinjiang Medical University, Urumchi, China, March 2016. [Google Scholar]
- Wang, L. Crosstalk between miR-19b and TGF-β1 Signaling of Hepatic Stellate Cell Proliferation. Master’s Thesis, Xinjiang University, Urumchi, China, June 2014. [Google Scholar]
- Zhang, C.; Wang, L.; Ali, T.; Li, L.; Bi, X.; Wang, J.; Lu, G.; Shao, Y.; Vuitton, D.A.; Wen, H.; et al. Hydatid cyst fluid promotes peri-cystic fibrosis in cystic echinococcosis by suppressing miR-19 expression. Parasit. Vectors 2016, 9, 278. [Google Scholar] [CrossRef] [PubMed]
- Brenner, D.A. Molecular pathogenesis of liver fibrosis. Trans. Am. Clin. Climatol. Assoc. 2009, 120, 361–368. [Google Scholar]
- Hernandezgea, V.; Friedman, S.L. Pathogenesis of Liver Fibrosis. Annu. Rev. Pathol. 2011, 6, 425–456. [Google Scholar] [CrossRef]
- Biernacka, A.; Dobaczewski, M.; Frangogiannis, N.G. TGF-β signaling in fibrosis. Growth Factors 2011, 29, 196–202. [Google Scholar] [CrossRef]
- Dooley, S.; Ten, P.D. TGF-β in progression of liver disease. Cell Tissue Res. 2012, 347, 245–256. [Google Scholar] [CrossRef]
- Gressner, A.M.; Weiskirchen, R.; Breitkopf, K.; Dooley, S. Roles of TGF-beta in hepatic fibrosis. Front. Biosci. 2002, 7, d793. [Google Scholar] [CrossRef]
- Ueno, H.; Sakamoto, T.; Nakamura, T.; Qi, Z.; Astuchi, N.; Takeshita, A.; Shimizu, K.; Ohashi, H. A soluble transforming growth factor beta receptor expressed in muscle prevents liver fibrogenesis and dysfunction in rats. Hum. Gene Ther. 2000, 11, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, C.; Wei, X.; Blagosklonov, O.; Lv, G.; Lu, X.; Mantion, G.; Vuitton, D.A.; Wen, H.; Lin, R. TGF-beta and TGF-beta/Smad signaling in the interactions between Echinococcus multilocularis and its hosts. PLoS ONE 2013, 8, e55379. [Google Scholar] [CrossRef]
- Liu, Y.; Abudounnasier, G.; Zhang, T.; Liu, X.; Wang, Q.; Yan, Y.; Ding, J.; Wen, H.; Yimiti, D.; Ma, X. Increased Expression of TGF-beta1 in Correlation with Liver Fibrosis during Echinococcus granulosus Infection in Mice. Korean J. Parasitol. 2016, 54, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Chen, X.; Peng, X.; Zhang, S.; Niu, J.; Sun, H. The specific expressions of collagenI, III and IV in the pericystic layer of hepatic hydatid cyst. Chin. J. Zoon. 2004, 876–879. [Google Scholar] [CrossRef]
- Wu, X.; Chen, X.; Peng, X.; Zhang, S.; Niu, J.; Sun, H. The special of TGF-β1 and TNF-αin the pericystic layer of hepatic hydatid cysy. Chin. J. End. 2004, 25–27. [Google Scholar] [CrossRef]
- Peng, X.; Wu, X.; Zhang, S.; Niu, J. Re-recognition of pathological structure of fibrocyst wall around hepatic hydatid cyst. World Chin. J. Digestol. 2005, 10–13. [Google Scholar] [CrossRef]
- Vuitton, D.A. The ambiguous role of immunity in echinococcosis: Protection of the host or of the parasite? Acta Trop. 2002, 85, 119–132. [Google Scholar] [CrossRef]
- Amiot, F.; Vuong, P.; Defontaines, M.; Pater, C.; Dautry, F.; Liance, M. Secondary alveolar echinococcosis in lymphotoxin-alpha and tumour necrosis factor-alpha deficient mice: Exacerbation of Echinococcus multilocularis larval growth is associated with cellular changes in the periparasitic granuloma. Parasite Immunol. 1999, 21, 475–483. [Google Scholar] [CrossRef]
- Yan, Y.; Liu, Y.; Yang, N.; Li, B.; Feng, N.; Jiang, T.; Sun, Q.; Wen, H.; Ding, J.; Ma, X. The expression of matrix metalloproteinase 2(MMP2) in mice infected with Echinococcus. Chin. J. Pathog. Biol. 2018, 13, 168–172. [Google Scholar]
- Forster, S.; Koziol, U.; Schafer, T.; Duvoisin, R.; Cailliau, K.; Vanderstraete, M.; Dissous, C.; Brehm, K. The role of fibroblast growth factor signalling in Echinococcus multilocularis development and host-parasite interaction. PLoS Negl. Trop. Dis. 2019, 13, e0006959. [Google Scholar] [CrossRef]
- Abbassioun, K.; Amirjamshidi, A. Diagnosis and Management of Hydatid Cyst of the Central Nervous System: Part 2: Hydatid Cysts of the Skull, Orbit, and Spine. Neurosurg. Q. 2001, 11, 10–16. [Google Scholar] [CrossRef]
- Fiennes, A.G.; Thomas, D.G. Combined medical and surgical treatment of spinal hydatid disease: A case report. J. Neurol. Neurosurg. Psychiatry 1982, 45, 927–930. [Google Scholar] [CrossRef]
- Sapkas, G.S.; Machinis, T.G.; Chloros, G.D.; Fountas, K.N.; Themistocleous, G.S.; Vrettakos, G. Spinal hydatid disease, a rare but existent pathological entity: Case report and review of the literature. South. Med. J. 2006, 99, 178–183. [Google Scholar] [CrossRef]
- Abdelhakim, K.; Khalil, A.; Haroune, B.; Oubaid, M.; Mondher, M. A case of sacral hydatid cyst. Int. J. Surg. Case Rep. 2014, 5, 434–436. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, A.A.; Allen, J.T.; Rogan, M.T. Echinococcus granulosus cyst fluid enhances epithelial-mesenchymal transition. Parasite Immunol. 2018, 40, e12533. [Google Scholar] [CrossRef]
- Nunes, M.C.; Guimarães Júnior, M.H.; Diamantino, A.C.; Gelape, C.L.; Ferrari, T.C. Cardiac manifestations of parasitic diseases. Heart 2017, 103, 651–658. [Google Scholar] [CrossRef]
- Labsi, M.; Soufli, I.; Khelifi, L.; Amir, Z.C.; Touil-Boukoffa, C. In vivo treatment with IL-17A attenuates hydatid cyst growth and liver fibrogenesis in an experimental model of echinococcosis. Acta Trop. 2018, 181, 6–10. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, S.; Niu, J.; Wu, X.; Abudu; Pan, H.; Mo, L.; Yang, H. A new principle of wurgical procedures of hepatic hydatid disease—An analysis of 648 cases. Fu Bu Wai Ke 2003, 16, 825–827. [Google Scholar]
- Peng, X.; Zhang, S.; Niu, J.; Wu, X.; Pan, H.; Mo, L.; Chen, Y.; Yang, H. Total subadventitial cystectomy for the treatment of 30 patients with hepatic hydatid cyst. Chin. J. Gen. Surg. 2002, 17, 529–530. [Google Scholar]
- Labsi, M.; Soufli, I.; Khelifi, L.; Amir, Z.C.; Touil-Boukoffa, C. A preventive effect of the combination of albendazole and pomegranate peel aqueous extract treatment in cystic echinococcosis mice model: An alternative approach. Acta Trop. 2019, 197, 105050. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, F.; Chong, S.; Qin, M.; Li, S.; Wei, R.; Zhao, Y. Mechanism of Fibrosis Induced by Echinococcus spp. Diseases 2019, 7, 51. https://doi.org/10.3390/diseases7030051
Niu F, Chong S, Qin M, Li S, Wei R, Zhao Y. Mechanism of Fibrosis Induced by Echinococcus spp. Diseases. 2019; 7(3):51. https://doi.org/10.3390/diseases7030051
Chicago/Turabian StyleNiu, Fuqiu, Shigui Chong, Mingqun Qin, Shenmei Li, Riming Wei, and Yumin Zhao. 2019. "Mechanism of Fibrosis Induced by Echinococcus spp." Diseases 7, no. 3: 51. https://doi.org/10.3390/diseases7030051
APA StyleNiu, F., Chong, S., Qin, M., Li, S., Wei, R., & Zhao, Y. (2019). Mechanism of Fibrosis Induced by Echinococcus spp. Diseases, 7(3), 51. https://doi.org/10.3390/diseases7030051