Adiponectin Isoforms Differentially Affect Gene Expression and the Lipidome of Primary Human Hepatocytes

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
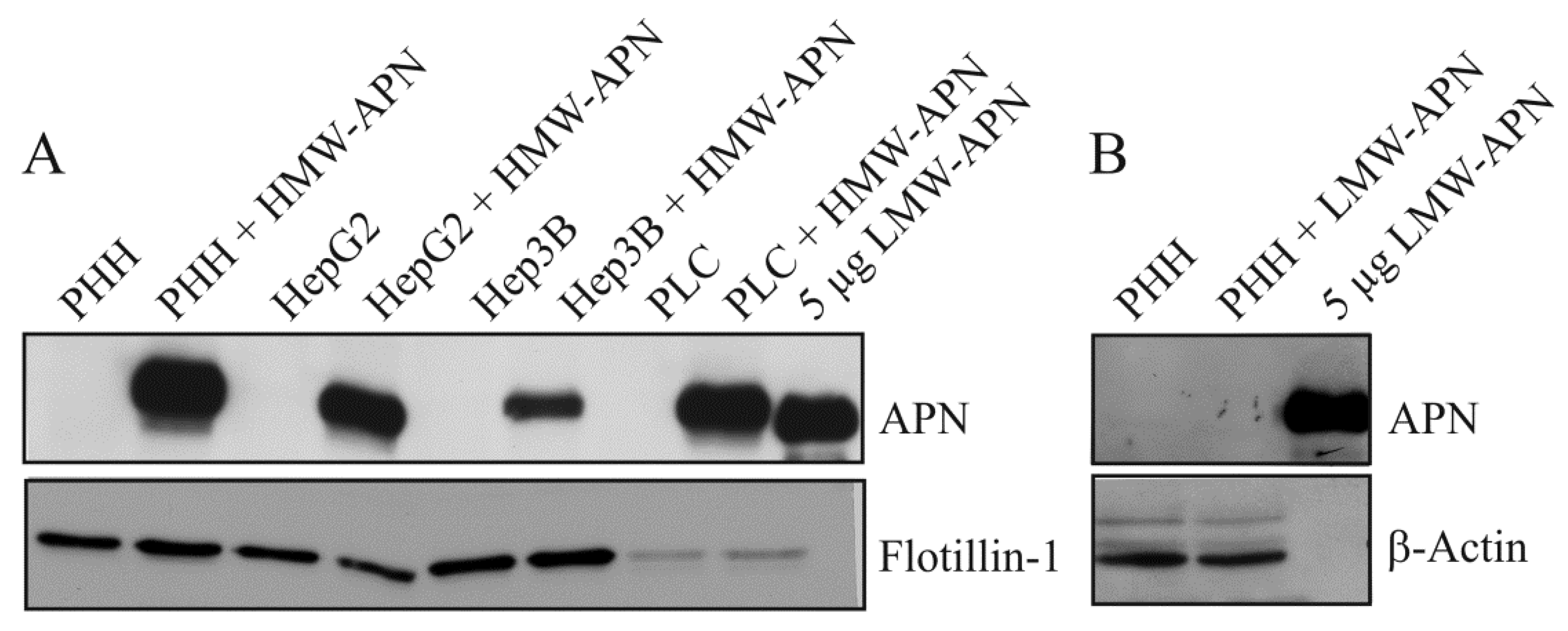
2.1. APN Isoforms in Hepatocyte Lysates

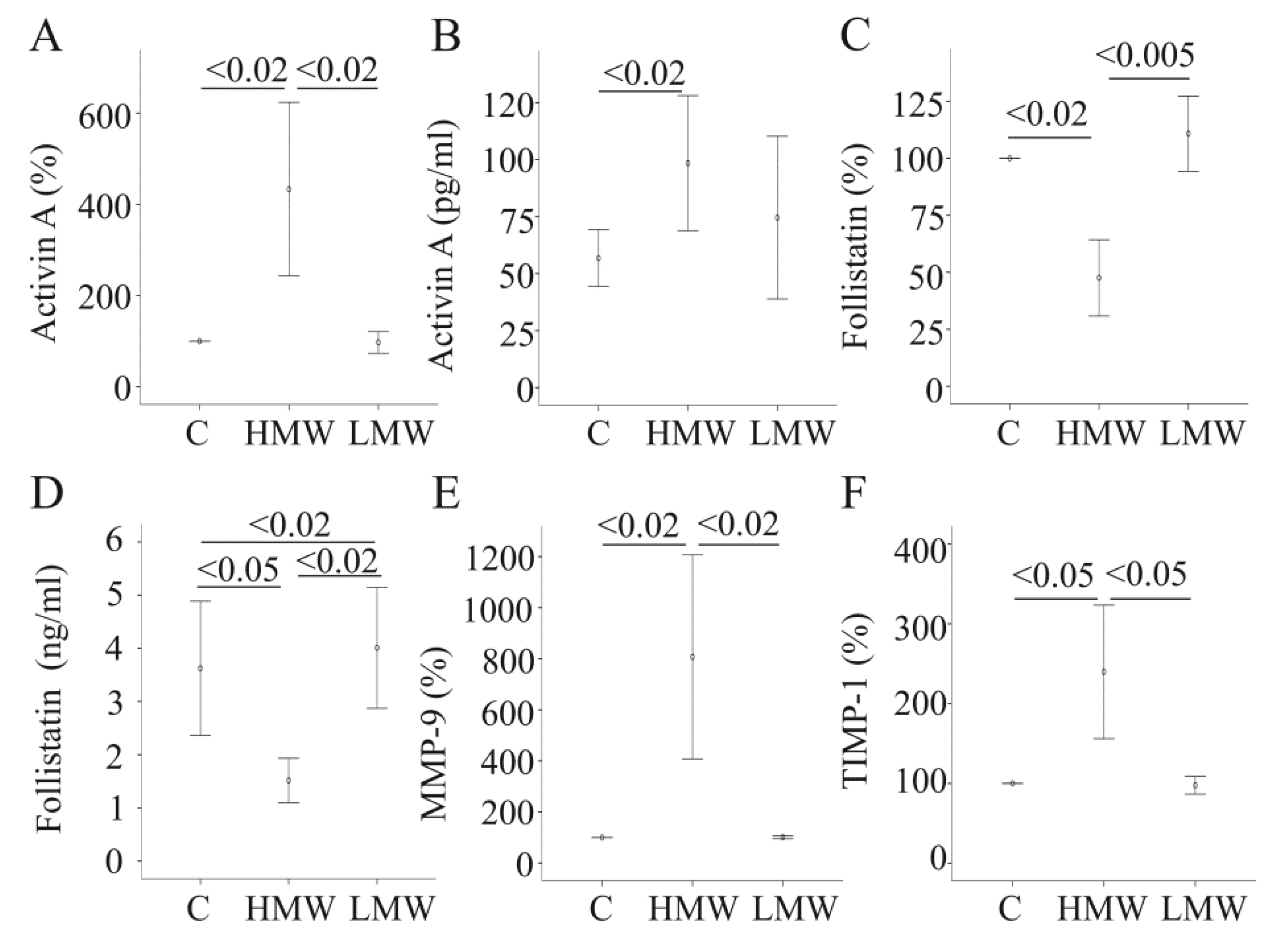
2.2. Effect of APN Isoforms on Genes/Proteins Involved in Fibrosis

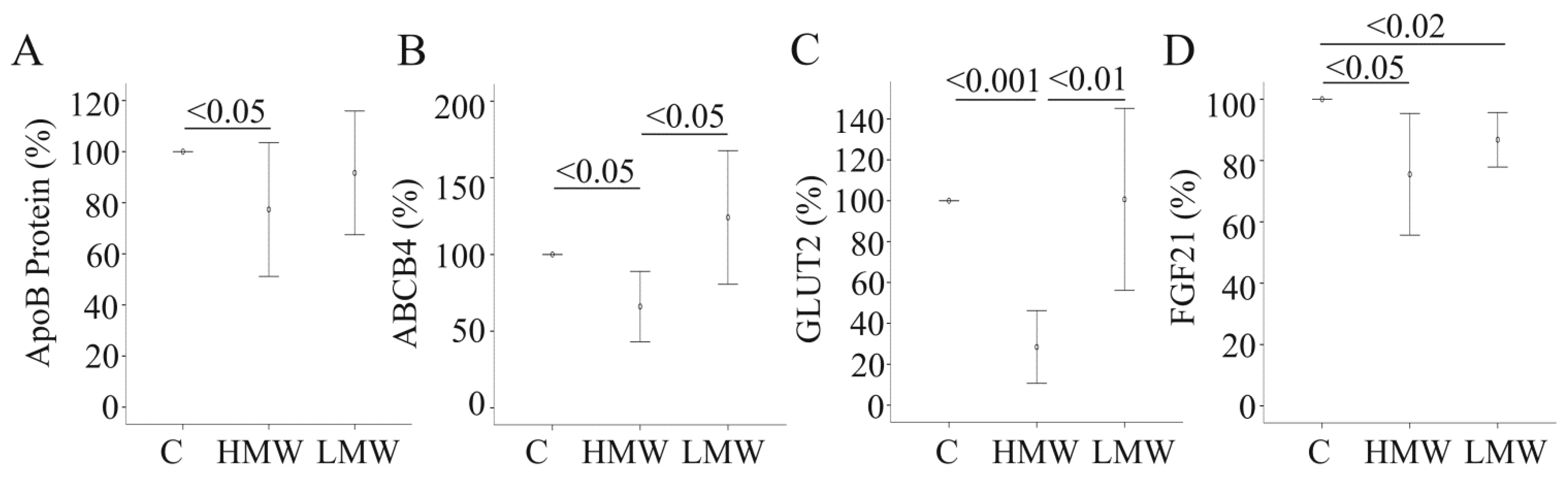
2.3. Effect of APN Isoforms on Genes Involved in Glucose and Lipid Metabolism

2.4. Effect of APN Isoforms on Fibroblast Growth Factor 21
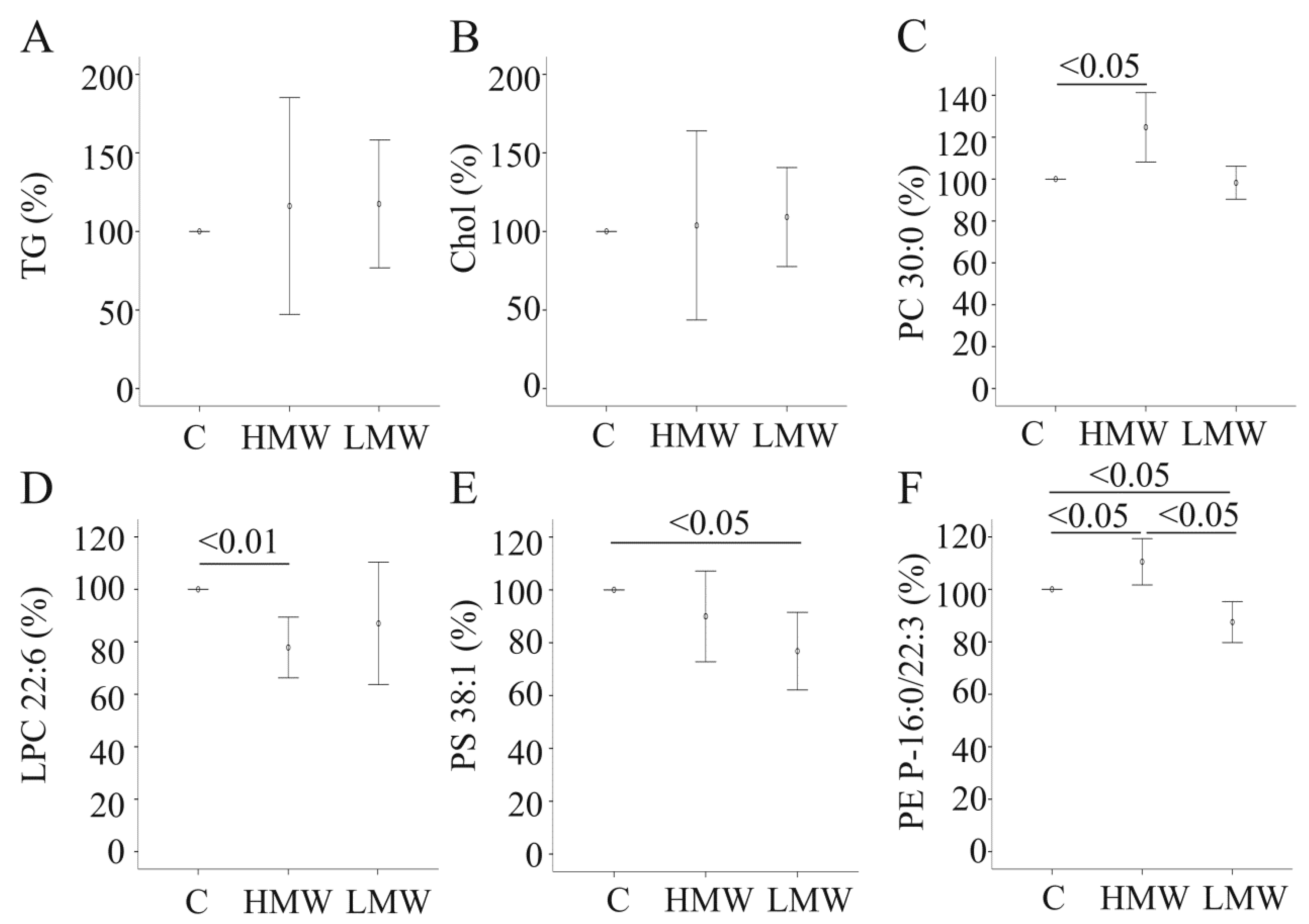
2.5. Effect of APN Isoforms on Triglycerides and Cholesterol

2.6. Effect of APN Isoforms on Phosphatidylcholine and Lysophosphatidylcholine
2.7. Effect of APN Isoforms on Phosphatidylethanolamine and Phosphatidylserine
2.8. Effect of APN Isoforms on Plasmalogens
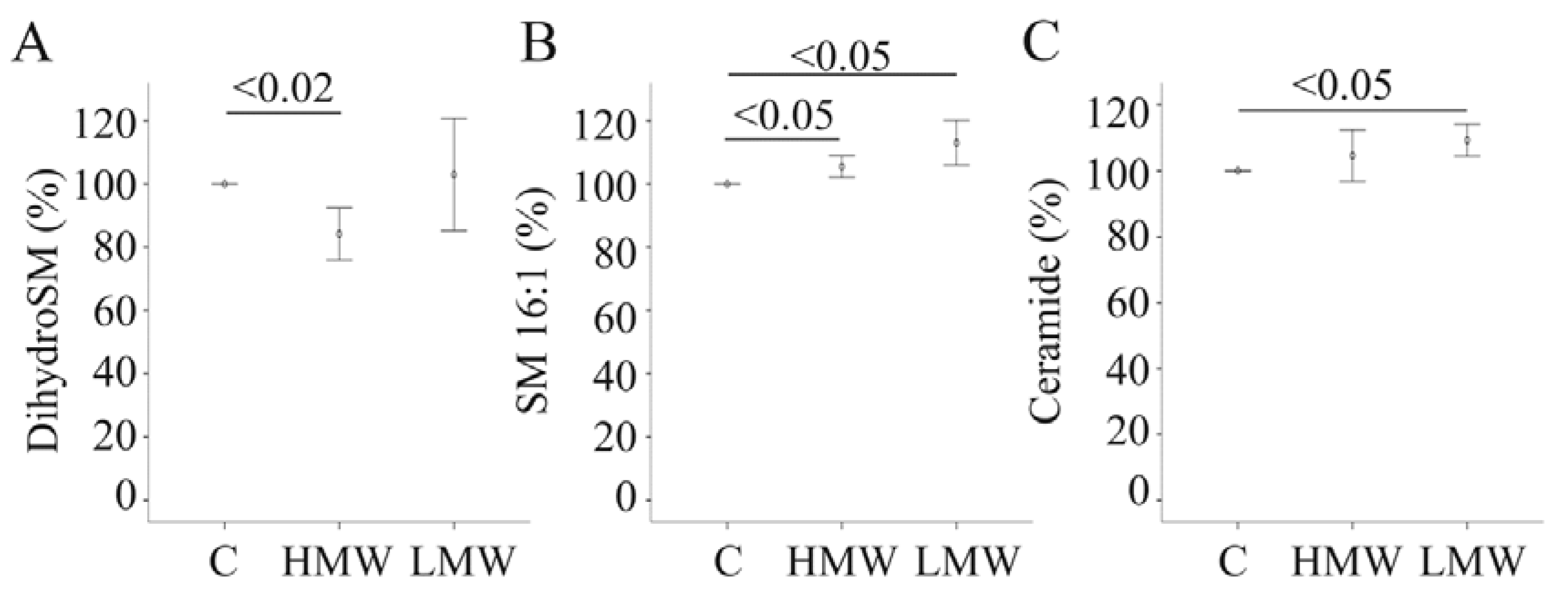
2.9. Effect of APN Isoforms on Sphingomyelin and Ceramides

3. Experimental Section
3.1. Materials
3.2. Primary Human Cells
3.3. Quantification of Lipids
3.4. Monitoring of Gene Expression by Real-Time RT-PCR
3.5. SDS-PAGE and Immunoblotting
3.6. ELISA
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schaffler, A.; Scholmerich, J.; Buchler, C. Mechanisms of disease: Adipocytokines and visceral adipose tissue–Emerging role in nonalcoholic fatty liver disease. Nat. Clin. Pract. Gastroenterol. Hepatol. 2005, 2, 273–280. [Google Scholar] [CrossRef]
- Buechler, C.; Wanninger, J.; Neumeier, M. Adiponectin, a key adipokine in obesity related liver diseases. World J. Gastroenterol. 2011, 17, 2801–2811. [Google Scholar]
- Handa, P.; Maliken, B.D.; Nelson, J.E.; Morgan-Stevenson, V.; Messner, D.J.; Dhillon, B.K.; Klintworth, H.M.; Beauchamp, M.; Yeh, M.M.; Elfers, C.T.; et al. Reduced adiponectin signaling due to weight gain results in nonalcoholic steatohepatitis through impaired mitochondrial biogenesis. Hepatology 2013. [Google Scholar] [CrossRef]
- Miller, R.A.; Chu, Q.; le Lay, J.; Scherer, P.E.; Ahima, R.S.; Kaestner, K.H.; Foretz, M.; Viollet, B.; Birnbaum, M.J. Adiponectin suppresses gluconeogenic gene expression in mouse hepatocytes independent of LKB1-AMPK signaling. J. Clin Invest. 2011, 121, 2518–2528. [Google Scholar] [CrossRef]
- Neumeier, M.; Sigruener, A.; Eggenhofer, E.; Weigert, J.; Weiss, T.S.; Schaeffler, A.; Schlitt, H.J.; Aslanidis, C.; Piso, P.; Langmann, T.; et al. High molecular weight adiponectin reduces apolipoprotein B and E release in human hepatocytes. Biochem. Biophys. Res. Commun. 2007, 352, 543–548. [Google Scholar] [CrossRef]
- Oku, H.; Matsuura, F.; Koseki, M.; Sandoval, J.C.; Yuasa-Kawase, M.; Tsubakio-Yamamoto, K.; Masuda, D.; Maeda, N.; Ohama, T.; Ishigami, M.; et al. Adiponectin deficiency suppresses ABCA1 expression and ApoA-I synthesis in the liver. FEBS Lett. 2007, 581, 5029–5033. [Google Scholar] [CrossRef]
- Wanninger, J.; Neumeier, M.; Hellerbrand, C.; Schacherer, D.; Bauer, S.; Weiss, T.S.; Huber, H.; Schaffler, A.; Aslanidis, C.; Scholmerich, J.; et al. Lipid accumulation impairs adiponectin-mediated induction of activin A by increasing TGFbeta in primary human hepatocytes. Biochim. Biophys. Acta 2011, 1811, 626–633. [Google Scholar] [CrossRef]
- Wanninger, J.; Walter, R.; Bauer, S.; Eisinger, K.; Schaffler, A.; Dorn, C.; Weiss, T.S.; Hellerbrand, C.; Buechler, C. MMP-9 activity is increased by adiponectin in primary human hepatocytes but even negatively correlates with serum adiponectin in a rodent model of non-alcoholic steatohepatitis. Exp. Mol. Pathol. 2011, 91, 603–607. [Google Scholar] [CrossRef]
- Yamauchi, T.; Nio, Y.; Maki, T.; Kobayashi, M.; Takazawa, T.; Iwabu, M.; Okada-Iwabu, M.; Kawamoto, S.; Kubota, N.; Kubota, T.; et al. Targeted disruption of AdipoR1 and AdipoR2 causes abrogation of adiponectin binding and metabolic actions. Nat. Med. 2007, 13, 332–339. [Google Scholar]
- Wedemeyer, I.; Bechmann, L.P.; Odenthal, M.; Jochum, C.; Marquitan, G.; Drebber, U.; Gerken, G.; Gieseler, R.K.; Dienes, H.P.; Canbay, A. Adiponectin inhibits steatotic CD95/Fas up-regulation by hepatocytes: Therapeutic implications for hepatitis C. J. Hepatol. 2009, 50, 140–149. [Google Scholar] [CrossRef]
- Bechmann, L.P.; Kocabayoglu, P.; Sowa, J.P.; Sydor, S.; Best, J.; Schlattjan, M.; Beilfuss, A.; Schmitt, J.; Hannivoort, R.A.; Kilicarslan, A.; et al. Free fatty acids repress small heterodimer partner (SHP) activation and adiponectin counteracts bile acid-induced liver injury in superobese patients with nonalcoholic steatohepatitis. Hepatology 2013, 57, 1394–1406. [Google Scholar] [CrossRef]
- Holland, W.L.; Miller, R.A.; Wang, Z.V.; Sun, K.; Barth, B.M.; Bui, H.H.; Davis, K.E.; Bikman, B.T.; Halberg, N.; Rutkowski, J.M.; et al. Receptor-mediated activation of ceramidase activity initiates the pleiotropic actions of adiponectin. Nat. Med. 2011, 17, 55–63. [Google Scholar] [CrossRef]
- Wanninger, J.; Liebisch, G.; Schmitz, G.; Bauer, S.; Eisinger, K.; Neumeier, M.; Ouchi, N.; Walsh, K.; Buechler, C. Lipidomic analysis of the liver identifies changes of major and minor lipid species in adiponectin deficient mice. Exp. Mol. Pathol. 2013, 94, 412–417. [Google Scholar]
- Rutkowski, J.M.; Scherer, P.E. Isolation and quantitation of adiponectin higher order complexes. Methods Enzymol. 2014, 537, 243–259. [Google Scholar] [CrossRef]
- Tsao, T.S.; Murrey, H.E.; Hug, C.; Lee, D.H.; Lodish, H.F. Oligomerization state-dependent activation of NF-kappa B signaling pathway by adipocyte complement-related protein of 30 kDa (Acrp30). J. Biol. Chem. 2002, 277, 29359–29362. [Google Scholar]
- Neumeier, M.; Weigert, J.; Schaffler, A.; Wehrwein, G.; Muller-Ladner, U.; Scholmerich, J.; Wrede, C.; Buechler, C. Different effects of adiponectin isoforms in human monocytic cells. J. Leukoc. Biol. 2006, 79, 803–808. [Google Scholar] [CrossRef]
- Wedellova, Z.; Kovacova, Z.; Tencerova, M.; Vedral, T.; Rossmeislova, L.; Siklova-Vitkova, M.; Stich, V.; Polak, J. The impact of full-length, trimeric and globular adiponectin on lipolysis in subcutaneous and visceral adipocytes of obese and non-obese women. PLoS One 2013, 8, e66783. [Google Scholar] [CrossRef]
- Pajvani, U.B.; Du, X.; Combs, T.P.; Berg, A.H.; Rajala, M.W.; Schulthess, T.; Engel, J.; Brownlee, M.; Scherer, P.E. Structure-function studies of the adipocyte-secreted hormone Acrp30/adiponectin. Implications fpr metabolic regulation and bioactivity. J. Biol. Chem. 2003, 278, 9073–9085. [Google Scholar] [CrossRef]
- Baessler, A.; Schlossbauer, S.; Stark, K.; Strack, C.; Riegger, G.; Schunkert, H.; Hengstenberg, C.; Fischer, M. Adiponectin multimeric forms but not total adiponectin levels are associated with myocardial infarction in non-diabetic men. J. Atheroscler. Thromb. 2011, 18, 616–627. [Google Scholar] [CrossRef]
- Schober, F.; Neumeier, M.; Weigert, J.; Wurm, S.; Wanninger, J.; Schaffler, A.; Dada, A.; Liebisch, G.; Schmitz, G.; Aslanidis, C.; et al. Low molecular weight adiponectin negatively correlates with the waist circumference and monocytic IL-6 release. Biochem. Biophys. Res. Commun. 2007, 361, 968–973. [Google Scholar] [CrossRef]
- Engl, J.; Sturm, W.; Sandhofer, A.; Kaser, S.; Tschoner, A.; Tatarczyk, T.; Weiss, H.; Tilg, H.; Patsch, J.R.; Ebenbichler, C.F. Effect of pronounced weight loss on visceral fat, liver steatosis and adiponectin isoforms. Eur. J. Clin. Invest. 2008, 38, 238–244. [Google Scholar] [CrossRef]
- Kaser, S.; Tatarczyk, T.; Stadlmayr, A.; Ciardi, C.; Ress, C.; Tschoner, A.; Sandhofer, A.; Paulweber, B.; Ebenbichler, C.F.; Patsch, J.R. Effect of obesity and insulin sensitivity on adiponectin isoform distribution. Eur. J. Clin. Invest. 2008, 38, 827–834. [Google Scholar] [CrossRef]
- Bredella, M.A.; Torriani, M.; Ghomi, R.H.; Thomas, B.J.; Brick, D.J.; Gerweck, A.V.; Harrington, L.M.; Miller, K.K. Adiponectin is inversely associated with intramyocellular and intrahepatic lipids in obese premenopausal women. Obesity (Silver Spring) 2011, 19, 911–916. [Google Scholar] [CrossRef]
- Goetz, R. Metabolism: Adiponectin—A mediator of specific metabolic actions of FGF21. Nat. Rev. Endocrinol. 2013, 9, 506–508. [Google Scholar] [CrossRef]
- Neumeier, M.; Hellerbrand, C.; Gabele, E.; Buettner, R.; Bollheimer, C.; Weigert, J.; Schaffler, A.; Weiss, T.S.; Lichtenauer, M.; Scholmerich, J.; et al. Adiponectin and its receptors in rodent models of fatty liver disease and liver cirrhosis. World J. Gastroenterol. 2006, 12, 5490–5494. [Google Scholar]
- Halberg, N.; Schraw, T.D.; Wang, Z.V.; Kim, J.Y.; Yi, J.; Hamilton, M.P.; Luby-Phelps, K.; Scherer, P.E. Systemic fate of the adipocyte-derived factor adiponectin. Diabetes 2009, 58, 1961–1970. [Google Scholar] [CrossRef]
- Yndestad, A.; Haukeland, J.W.; Dahl, T.B.; Bjoro, K.; Gladhaug, I.P.; Berge, C.; Damas, J.K.; Haaland, T.; Loberg, E.M.; Linnestad, P.; et al. A complex role of activin A in non-alcoholic fatty liver disease. Am. J. Gastroenterol. 2009, 104, 2196–2205. [Google Scholar] [CrossRef]
- D’Amico, F.; Consolo, M.; Amoroso, A.; Skarmoutsou, E.; Mauceri, B.; Stivala, F.; Malaponte, G.; Bertino, G.; Neri, S.; Mazzarino, M.C. Liver immunolocalization and plasma levels of MMP-9 in non-alcoholic steatohepatitis (NASH) and hepatitis C infection. Acta Histochem. 2010, 112, 474–481. [Google Scholar] [CrossRef]
- Nakasone, H.; Terasako-Saito, K.; Yamazaki, R.; Sato, M.; Tanaka, Y.; Sakamoto, K.; Kurita, M.; Yamasaki, R.; Wada, H.; Ishihara, Y.; et al. Impact of high-/middle-molecular-weight adiponectin on the synthesis and regulation of extracellular matrix in dermal fibroblasts. Exp. Hematol. 2014, 42, 261–273. [Google Scholar] [CrossRef]
- Saja, K.; Sudhakaran, P.R. cAMP-mediated upregulation of gelatinases in primary cultures of isolated rat hepatocytes. Indian J. Biochem. Biophys. 2004, 41, 294–298. [Google Scholar]
- Holland, W.L.; Adams, A.C.; Brozinick, J.T.; Bui, H.H.; Miyauchi, Y.; Kusminski, C.M.; Bauer, S.M.; Wade, M.; Singhal, E.; Cheng, C.C.; et al. An FGF21-adiponectin-ceramide axis controls energy expenditure and insulin action in mice. Cell. Metab. 2013, 17, 790–797. [Google Scholar] [CrossRef]
- Braverman, N.E.; Moser, A.B. Functions of plasmalogen lipids in health and disease. Biochim. Biophys. Acta 2012, 1822, 1442–1452. [Google Scholar] [CrossRef]
- Kolak, M.; Westerbacka, J.; Velagapudi, V.R.; Wagsater, D.; Yetukuri, L.; Makkonen, J.; Rissanen, A.; Hakkinen, A.M.; Lindell, M.; Bergholm, R.; et al. Adipose tissue inflammation and increased ceramide content characterize subjects with high liver fat content independent of obesity. Diabetes 2007, 56, 1960–1968. [Google Scholar] [CrossRef]
- Thasler, W.E.; Weiss, T.S.; Schillhorn, K.; Stsoll, P.T.; Irrgang, B.; Jauch, K.W. Charitable state-controlled foundation human tissue and cell research: Ethic and legal aspects in the supply of surgically removed human tissue for research in the academic and commercial sector in Germany. Cell Tissue Bank 2003, 4, 49–56. [Google Scholar] [CrossRef]
- Damm, G.P.E.; Burkhardt, B.; Vermehren, J.; Nüssler, A.K.; Weiss, T.S. Human parenchymal and non-parenchymal liver cell isolation, culture and characterization. Hepatol. Int. 2013, 7, 951–958. [Google Scholar] [CrossRef]
- Godoy, P.; Hewitt, N.J.; Albrecht, U.; Andersen, M.E.; Ansari, N.; Bhattacharya, S.; Bode, J.G.; Bolleyn, J.; Borner, C.; Bottger, J.; et al. Recent advances in 2D and 3D in vitro systems using primary hepatocytes, alternative hepatocyte sources and non-parenchymal liver cells and their use in investigating mechanisms of hepatotoxicity, cell signaling and ADME. Arch. Toxicol. 2013, 87, 1315–1530. [Google Scholar] [CrossRef] [Green Version]
- Liebisch, G.; Lieser, B.; Rathenberg, J.; Drobnik, W.; Schmitz, G. High-throughput quantification of phosphatidylcholine and sphingomyelin by electrospray ionization tandem mass spectrometry coupled with isotope correction algorithm. Biochim. Biophys. Acta 2004, 1686, 108–117. [Google Scholar] [CrossRef]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef]
- Liebisch, G.; Drobnik, W.; Reil, M.; Trumbach, B.; Arnecke, R.; Olgemoller, B.; Roscher, A.; Schmitz, G. Quantitative measurement of different ceramide species from crude cellular extracts by electrospray ionization tandem mass spectrometry (ESI-MS/MS). J. Lipid Res. 1999, 40, 1539–1546. [Google Scholar]
- Liebisch, G.; Vizcaino, J.A.; Kofeler, H.; Trotzmuller, M.; Griffiths, W.J.; Schmitz, G.; Spener, F.; Wakelam, M.J. Shorthand notation for lipid structures derived from mass spectrometry. J. Lipid Res. 2013, 54, 1523–1530. [Google Scholar] [CrossRef]
- Frommer, K.W.; Schaffler, A.; Buchler, C.; Steinmeyer, J.; Rickert, M.; Rehart, S.; Brentano, F.; Gay, S.; Muller-Ladner, U.; Neumann, E. Adiponectin isoforms: A potential therapeutic target in rheumatoid arthritis? Ann. Rheum. Dis. 2012, 71, 1724–1732. [Google Scholar] [CrossRef] [Green Version]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wanninger, J.; Liebisch, G.; Eisinger, K.; Neumeier, M.; Aslanidis, C.; Voggenreiter, L.; Pohl, R.; Weiss, T.S.; Krautbauer, S.; Buechler, C. Adiponectin Isoforms Differentially Affect Gene Expression and the Lipidome of Primary Human Hepatocytes. Metabolites 2014, 4, 394-407. https://doi.org/10.3390/metabo4020394
Wanninger J, Liebisch G, Eisinger K, Neumeier M, Aslanidis C, Voggenreiter L, Pohl R, Weiss TS, Krautbauer S, Buechler C. Adiponectin Isoforms Differentially Affect Gene Expression and the Lipidome of Primary Human Hepatocytes. Metabolites. 2014; 4(2):394-407. https://doi.org/10.3390/metabo4020394
Chicago/Turabian StyleWanninger, Josef, Gerhard Liebisch, Kristina Eisinger, Markus Neumeier, Charalampos Aslanidis, Lisa Voggenreiter, Rebekka Pohl, Thomas S. Weiss, Sabrina Krautbauer, and Christa Buechler. 2014. "Adiponectin Isoforms Differentially Affect Gene Expression and the Lipidome of Primary Human Hepatocytes" Metabolites 4, no. 2: 394-407. https://doi.org/10.3390/metabo4020394