Delta-Tocotrienol Modulates Glutamine Dependence by Inhibiting ASCT2 and LAT1 Transporters in Non-Small Cell Lung Cancer (NSCLC) Cells: A Metabolomic Approach

Abstract
:1. Introduction
2. Results
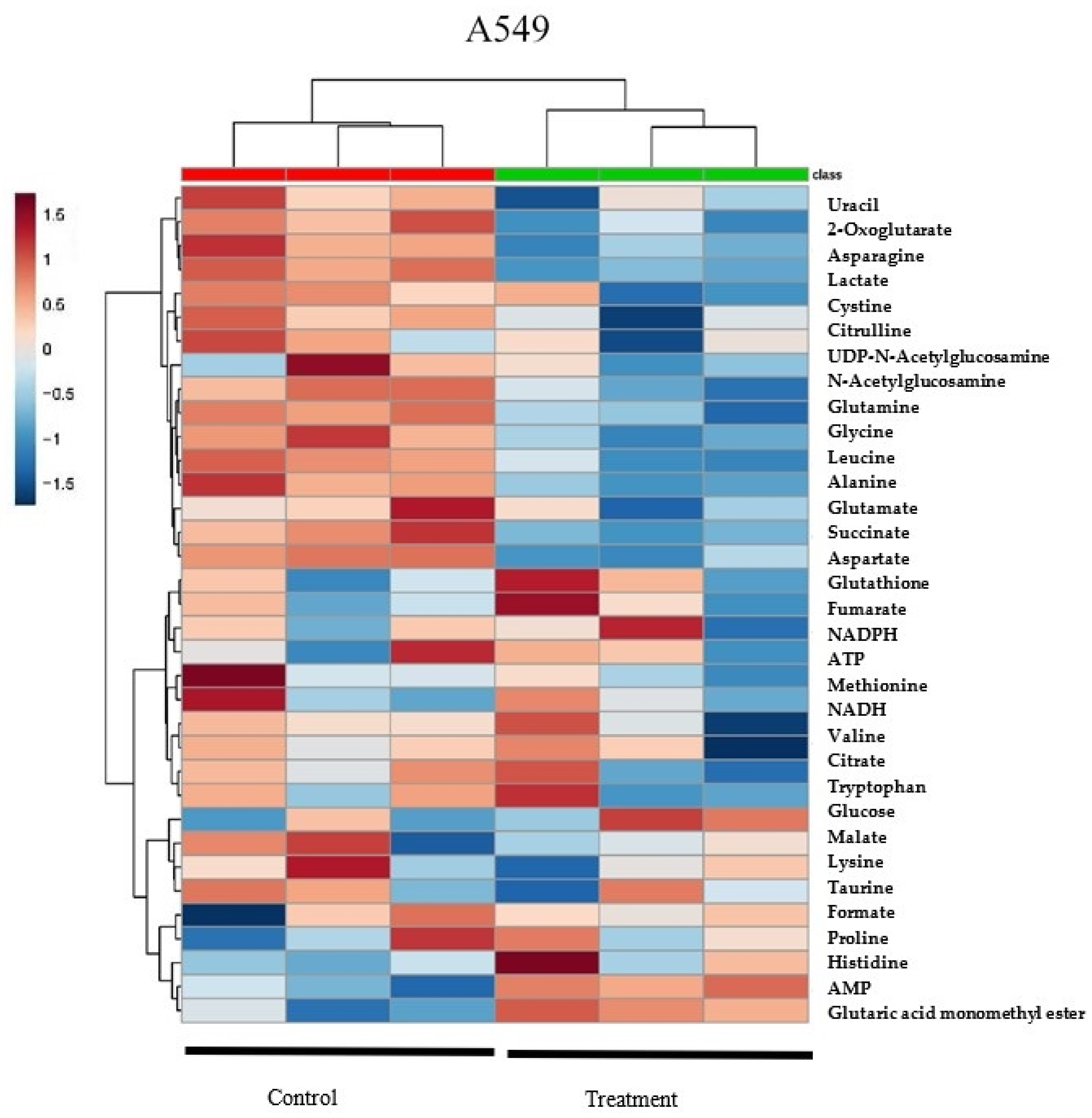
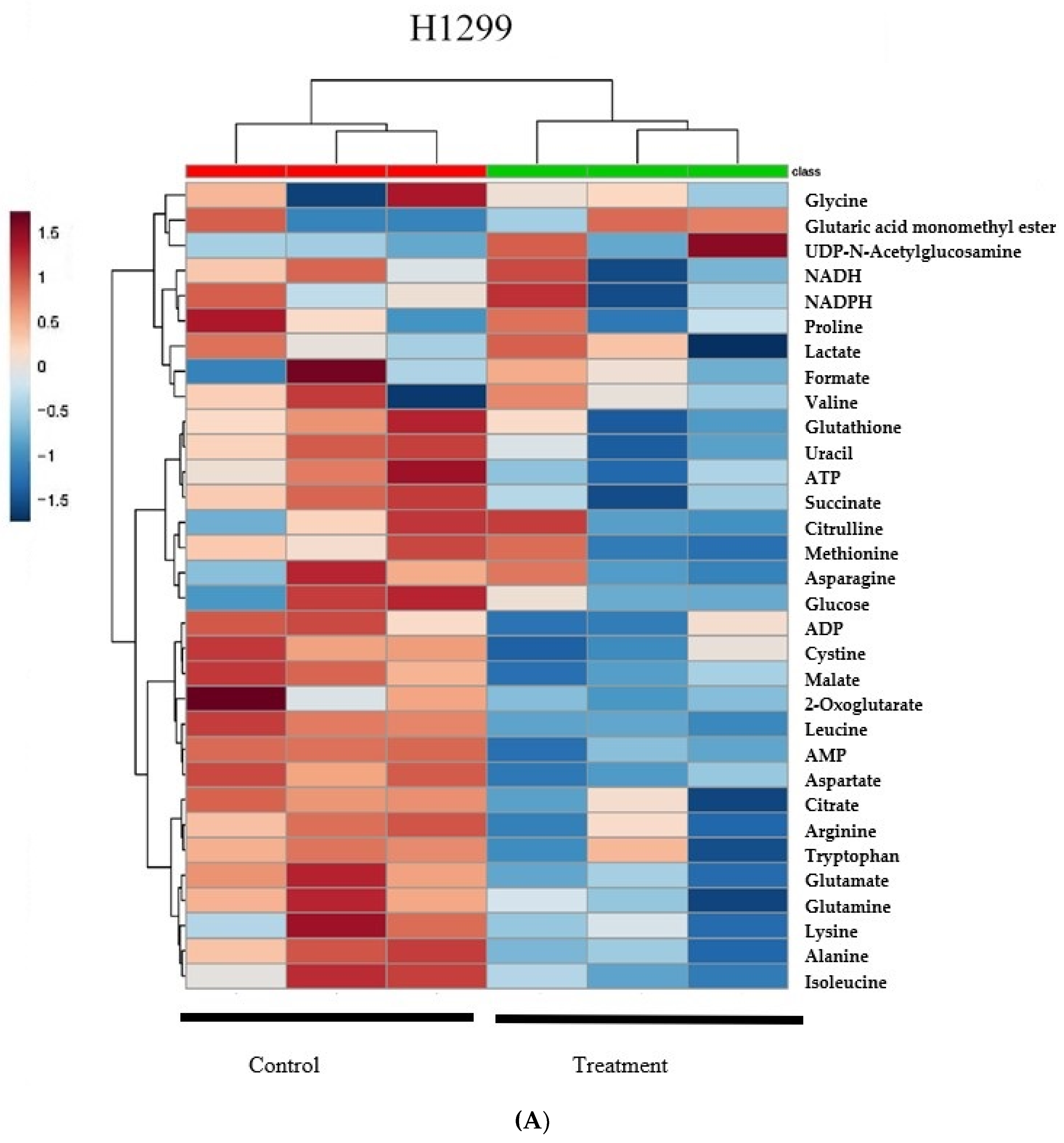
2.1. δT Changes Metabolite Profiles in A549 and H1299 Cells
2.2. Quantification of Metabolites Reveals That δT Alters Glutamine Metabolism
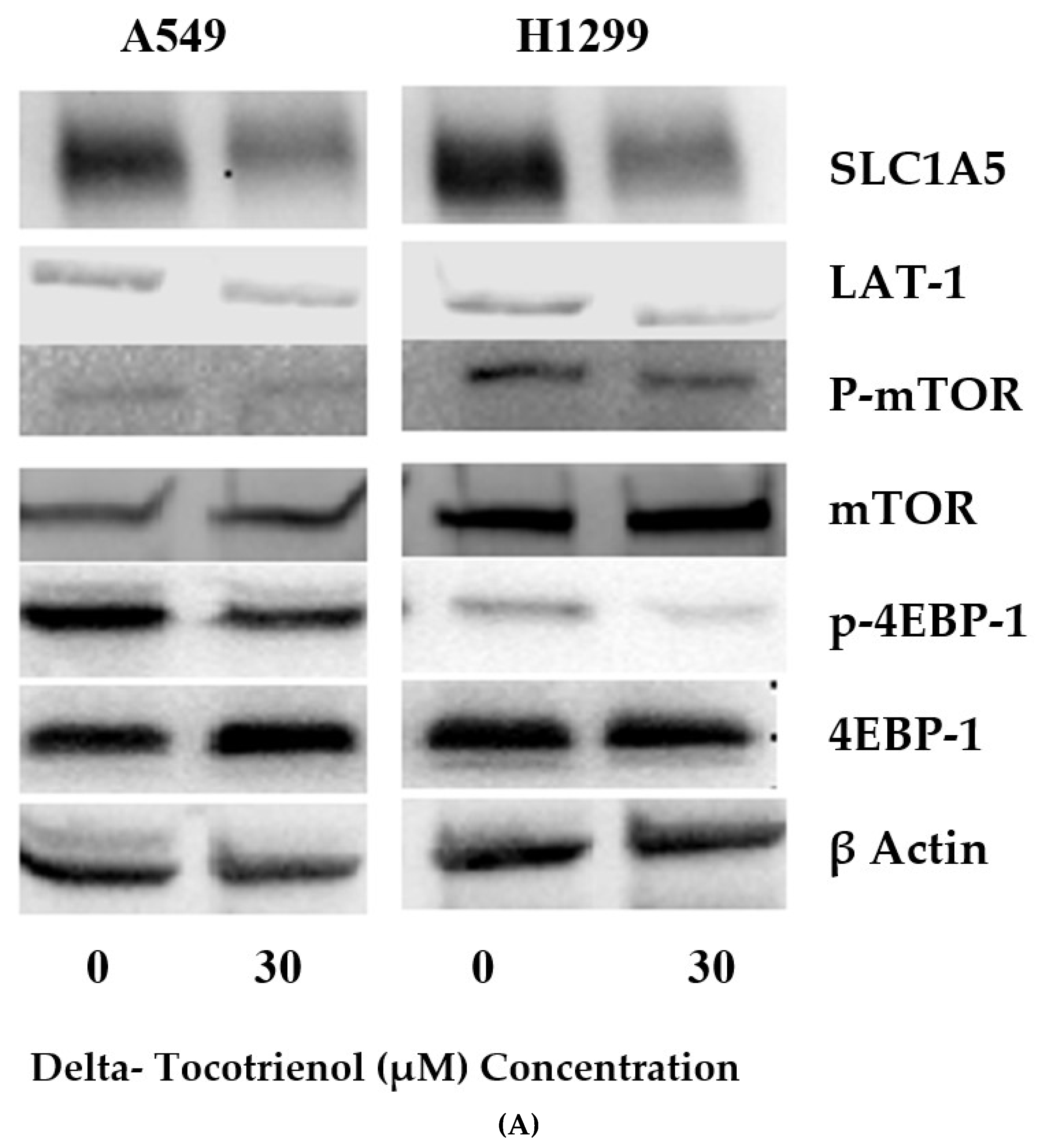
2.3. δT Inhibits Glutamine Transporters (LAT-1 and ASCT2) and the mTOR Pathway in A549 and H1299 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatment with δ-T
4.2. Intracellular Metabolite Extraction and Determination
4.3. 1H-NMR Spectroscopy
4.4. 1H-NMR Spectroscopy Processing
4.5. Quality Control
4.6. Multivariate Data Analysis: OPLS-DA
4.7. Metabolite Identification and Quantification from Chenomx NMR Suite
4.8. Additional Multivariate Data Analysis and Metabolic Pathway Identification Using MetaboAnalyst 3.0 Software
4.9. Western Blot for Protein Expression Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Society, A.C. Lung Cancer (Non-Small Cell). Available online: http://www.cancer.org/acs/groups/cid/documents/webcontent/003115-pdf.pdf (accessed on 09 March 2019).
- American Cancer Society. Cancer Facts & Figures 2016; American Cancer Society: Atlanta, GA, USA, 2016. [Google Scholar]
- Kelsey, C.R.; Clough, R.W.; Marks, L.B. Local Recurrence Following Initial Resection of NSCLC: Salvage Is Possible with Radiation Therapy. Cancer J. 2006, 12, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Gold, K.A.; Lippman, S.M. Natural-agent mechanisms and early-phase clinical development. Top. Curr. Chem. 2013, 329, 241–252. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Shishodia, S. Molecular targets of dietary agents for prevention and therapy of cancer. Biochem. Pharmacol. 2006, 71, 1397–1421. [Google Scholar] [CrossRef]
- Surh, Y.J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer 2003, 3, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Theriault, A.; Chao, J.-T.; Wang, Q.; Gapor, A.; Adeli, K. Tocotrienol: A review of its therapeutic potential. Clin. Biochem. 1999, 32, 309–319. [Google Scholar] [CrossRef]
- De Silva, L.; Chuah, L.H.; Meganathan, P.; Fu, J.-Y. Tocotrienol and cancer metastasis. BioFactors 2016, 42, 149–162. [Google Scholar] [CrossRef]
- Constantinou, C.; Papas, A.; Constantinou, A.I. Vitamin E and cancer: An insight into the anticancer activities of vitamin E isomers and analogs. Int. J. Cancer 2008, 123, 739–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajasinghe, L.D. Anti-Cancer Effects of Tocotrienols in NSCLC. Ph.D. Thesis, Wayne State University, Detroit, MI, USA, 2017. [Google Scholar]
- Ji, X.; Wang, Z.; Geamanu, A.; Sarkar, F.H.; Gupta, S.V. Inhibition of cell growth and induction of apoptosis in non-small cell lung cancer cells by delta-tocotrienol is associated with notch-1 down-regulation. J. Cell. Biochem. 2011, 112, 2773–2783. [Google Scholar] [CrossRef]
- Rajasinghe, L.D.; Pindiprolu, R.H.; Gupta, S.V. Delta-tocotrienol inhibits non-small-cell lung cancer cell invasion via the inhibition of NF-κB, uPA activator, and MMP-9. OncoTargets Ther. 2018, 11, 4301–4314. [Google Scholar] [CrossRef]
- Kwon, H.; Oh, S.; Jin, X.; An, Y.J.; Park, S. Cancer metabolomics in basic science perspective. Arch. Pharm. Res. 2015, 38, 372–380. [Google Scholar] [CrossRef]
- Puchades-Carrasco, L.; Pineda-Lucena, A. Metabolomics Applications in Precision Medicine: An Oncological Perspective. Curr. Top. Med. Chem. 2017, 17, 2740–2751. [Google Scholar] [CrossRef]
- Tran, Q.; Lee, H.; Park, J.; Kim, S.H.; Park, J. Targeting Cancer Metabolism—Revisiting the Warburg Effects. Toxicol. Res. 2016, 32, 177–193. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.; Deng, X.; Khuri, F.R.; Owonikoko, T.K. Altered glutamine metabolism and therapeutic opportunities for lung cancer. Clin. Lung Cancer 2014, 15, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, I.B.; Lim, S.-W.; Loh, H.-S. Synergistic Apoptotic Effects of Tocotrienol Isomers and Acalypha wilkesiana on A549 and U87MG Cancer Cells. Trop. Life Sci. Res. 2018, 29, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Zhdanov, A.V.; Waters, A.H.C.; Golubeva, A.V.; Dmitriev, R.I.; Papkovsky, D.B. Availability of the key metabolic substrates dictates the respiratory response of cancer cells to the mitochondrial uncoupling. Biochim. Biophys. Acta BBA-Bioenerg. 2014, 1837, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez Herrera, K.N.; Lee, J.; Haigis, M.C. Intersections between mitochondrial sirtuin signaling and tumor cell metabolism. Crit. Rev. Biochem. Mol. Biol. 2015, 50, 242–255. [Google Scholar] [CrossRef] [Green Version]
- Robert, S.M.; Sontheimer, H. Glutamate Transporters in the Biology of Malignant Gliomas. Cell. Mol. Life Sci. CMLS 2014, 71, 1839–1854. [Google Scholar] [CrossRef]
- Fuchs, B.C.; Finger, R.E.; Onan, M.C.; Bode, B.P. ASCT2 silencing regulates mammalian target-of-rapamycin growth and survival signaling in human hepatoma cells. Am. J. Physiol. Cell Physiol. 2007, 293, C55–C63. [Google Scholar] [CrossRef]
- Shimizu, K.; Kaira, K.; Tomizawa, Y.; Sunaga, N.; Kawashima, O.; Oriuchi, N.; Tominaga, H.; Nagamori, S.; Kanai, Y.; Yamada, M.; et al. ASC amino-acid transporter 2 (ASCT2) as a novel prognostic marker in non-small cell lung cancer. Br. J. Cancer 2014, 110, 2030–2039. [Google Scholar] [CrossRef] [Green Version]
- Conciatori, F.; Ciuffreda, L.; Bazzichetto, C.; Falcone, I.; Pilotto, S.; Bria, E.; Cognetti, F.; Milella, M. mTOR Cross-Talk in Cancer and Potential for Combination Therapy. Cancers 2018, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.J.; Khelifa, S.; Feng, Y.; Lau, E.; Cardiff, R.; Kim, H.; Rimm, D.L.; Kluger, Y.; Ronai, Z.e. Abstract 2440: RNF5 mediates ER stress-induced degradation of SLC1A5 in breast cancer. Cancer Res. 2014, 74, 2440. [Google Scholar] [CrossRef]
- Shimizu, K.; Kaira, K.; Tomizawa, Y.; Sunaga, N.; Kawashima, O.; Oriuchi, N.; Kana, Y.; Yamada, M.; Oyama, T.; Takeyoshi, I. P0143 ASC amino acid transporter 2 (ASCT2) as a novel prognostic marker in non-small-cell lung cancer. Eur. J. Cancer 2014, 50, e49. [Google Scholar] [CrossRef]
- Fuchs, B.C.; Bode, B.P. Amino acid transporters ASCT2 and LAT1 in cancer: Partners in crime? Semin. Cancer Biol. 2005, 15, 254–266. [Google Scholar] [CrossRef]
- van den Heuvel, A.P.J.; Jing, J.; Wooster, R.F.; Bachman, K.E. Analysis of glutamine dependency in non-small cell lung cancer: GLS1 splice variant GAC is essential for cancer cell growth. Cancer Biol. Ther. 2012, 13, 1185–1194. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Hardie, R.A.; Hoy, A.J.; van Geldermalsen, M.; Gao, D.; Fazli, L.; Sadowski, M.C.; Balaban, S.; Schreuder, M.; Nagarajah, R.; et al. Targeting ASCT2-mediated glutamine uptake blocks prostate cancer growth and tumour development. J. Pathol. 2015, 236, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Imai, H.; Kaira, K.; Oriuchi, N.; Shimizu, K.; Tominaga, H.; Yanagitani, N.; Sunaga, N.; Ishizuka, T.; Nagamori, S.; Promchan, K. Inhibition of L-type amino acid transporter 1 has antitumor activity in non-small cell lung cancer. Anticancer Res. 2010, 30, 4819–4828. [Google Scholar]
- Fuchs, B.C.; Bode, B.P. Stressing out over survival: Glutamine as an apoptotic modulator. J. Surg. Res. 2006, 131, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Matés, J.M.; Segura, J.A.; Alonso, F.J.; Márquez, J. Pathways from glutamine to apoptosis. Front. Biosci. 2006, 11, 3164–3180. [Google Scholar] [CrossRef]
- Rajasinghe, L.; Gupta, S. Tocotrienols suppress non-small lung cancer cells via downregulation of the Notch-1 signaling pathway (644.1). FASEB J. 2014, 28. [Google Scholar] [CrossRef]
- Rajasinghe, L.; Pindiprolu, R.; Razalli, N.; Wu, Y.; Gupta, S. Delta Tocotrienol Inhibits MMP-9 Dependent Invasion and Metastasis of Non-Small Cell Lung Cancer (NSCLC) Cell by Suppressing Notch-1 Mediated NF-κb and uPA Pathways. FASEB J. 2015, 29. [Google Scholar] [CrossRef]
- Rajasinghe, L.D.; Gupta, S.V. Tocotrienol-rich mixture inhibits cell proliferation and induces apoptosis via down-regulation of the Notch-1/NF-κB pathways in NSCLC cells. Nutr. Diet. Suppl. 2017, 9, 103–114. [Google Scholar] [CrossRef]
- Rajasinghe, L.D.; Gupta, S.V. Delta Tocotrienal Inhibit mTOR Pathway by Modulating Glutamine Uptake and Transporters in Non-Small Cell Lung Cancer. FASEB J. 2016, 30. [Google Scholar] [CrossRef]
- Jewell, J.L.; Kim, Y.C.; Russell, R.C.; Yu, F.-X.; Park, H.W.; Plouffe, S.W.; Tagliabracci, V.S.; Guan, K.-L. Differential regulation of mTORC1 by leucine and glutamine. Science 2015, 347, 194–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petronini, P.G.; Urbani, S.; Alfieri, R.; Borghetti, A.F.; Guidotti, G.G. Cell susceptibility to apoptosis by glutamine deprivation and rescue: Survival and apoptotic death in cultured lymphoma-leukemia cell lines. J. Cell. Physiol. 1996, 169, 175–185. [Google Scholar] [CrossRef]
- Folkers, K.; Satyamitra, M.; Srinivasan, V. Delta-tocotrienol Mediates the Cellular Response to Radiation-Induced DNA Damage through Upregulation of Anti-Apoptotic Effectors in Human Lung Fibroblast Cell Lines. FASEB J. 2015, 29. [Google Scholar] [CrossRef]
- Husain, K.; Centeno, B.A.; Coppola, D.; Trevino, J.; Sebti, S.M.; Malafa, M.P. δ-Tocotrienol, a natural form of vitamin E, inhibits pancreatic cancer stem-like cells and prevents pancreatic cancer metastasis. Oncotarget 2017, 8, 31554–31567. [Google Scholar] [CrossRef] [Green Version]
- Montagnani Marelli, M.; Marzagalli, M.; Moretti, R.M.; Beretta, G.; Casati, L.; Comitato, R.; Gravina, G.L.; Festuccia, C.; Limonta, P. Vitamin E δ-tocotrienol triggers endoplasmic reticulum stress-mediated apoptosis in human melanoma cells. Sci. Rep. 2016, 6, 30502. [Google Scholar] [CrossRef] [PubMed]
- Swift, S.N.; Pessu, R.L.; Chakraborty, K.; Villa, V.; Lombardini, E.; Ghosh, S.P. Acute toxicity of subcutaneously administered vitamin E isomers delta-and gamma-tocotrienol in mice. Int. J. Toxicol. 2014, 33, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Mahipal, A.; Klapman, J.; Vignesh, S.; Yang, C.S.; Neuger, A.; Chen, D.-T.; Malafa, M.P. Pharmacokinetics and safety of vitamin E δ-tocotrienol after single and multiple doses in healthy subjects with measurement of vitamin E metabolites. Cancer Chemother. Pharmacol. 2016, 78, 157–165. [Google Scholar] [CrossRef]
- Shen, C.-L.; Wang, S.; Yang, S.; Tomison, M.D.; Abbasi, M.; Hao, L.; Scott, S.; Khan, M.S.; Romero, A.W.; Felton, C.K. A 12-week evaluation of annatto tocotrienol supplementation for postmenopausal women: Safety, quality of life, body composition, physical activity, and nutrient intake. BMC Complement. Altern. Med. 2018, 18, 198. [Google Scholar] [CrossRef] [PubMed]
- Scalise, M.; Galluccio, M.; Console, L.; Pochini, L.; Indiveri, C. The Human SLC7A5 (LAT1): The Intriguing Histidine/Large Neutral Amino Acid Transporter and Its Relevance to Human Health. Front. Chem. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Shin, G.; Kang, T.-W.; Yang, S.; Baek, S.-J.; Jeong, Y.-S.; Kim, S.-Y. GENT: Gene Expression Database of Normal and Tumor Tissues. Cancer Inform. 2011, 10, CIN–S7226. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, K.; Matsuo, H.; Kanai, Y.; Endou, H.; Hiroi, S.; Tominaga, S.; Mukai, M.; Ikeda, E.; Ozeki, Y.; Aida, S.; et al. LAT1 expression in normal lung and in atypical adenomatous hyperplasia and adenocarcinoma of the lung. Virchows Archiv 2006, 448, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Hassanein, M.; Hoeksema, M.D.; Shiota, M.; Qian, J.; Harris, B.K.; Chen, H.; Clark, J.E.; Alborn, W.E.; Eisenberg, R.; Massion, P.P. SLC1A5 mediates glutamine transport required for lung cancer cell growth and survival. Clin. Cancer Res. 2013, 19, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Masle-Farquhar, E.; Bröer, A.; Yabas, M.; Enders, A.; Bröer, S. ASCT2 (SLC1A5)-Deficient Mice Have Normal B-Cell Development, Proliferation, and Antibody Production. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Cui, H. Targeting Glutamine Induces Apoptosis: A Cancer Therapy Approach. Int. J. Mol. Sci. 2015, 16, 22830–22855. [Google Scholar] [CrossRef] [Green Version]
- Kanai, Y.; Hediger, M.A. The glutamate/neutral amino acid transporter family SLC1: Molecular, physiological and pharmacological aspects. Pflugers Archiv 2004, 447, 469–479. [Google Scholar] [CrossRef]
- Petroulakis, E.; Mamane, Y.; Le Bacquer, O.; Shahbazian, D.; Sonenberg, N. mTOR signaling: Implications for cancer and anticancer therapy. Br. J. Cancer 2006, 94, 195–199. [Google Scholar] [CrossRef]
- Shaw, R.J.; Cantley, L.C. Ras, PI (3) K and mTOR signalling controls tumour cell growth. Nature 2006, 441, 424–430. [Google Scholar] [CrossRef]
- Yamauchi, K.; Sakurai, H.; Kimura, T.; Wiriyasermkul, P.; Nagamori, S.; Kanai, Y.; Kohno, N. System L amino acid transporter inhibitor enhances anti-tumor activity of cisplatin in a head and neck squamous cell carcinoma cell line. Cancer Lett. 2009, 276, 95–101. [Google Scholar] [CrossRef]
- Laplante, M.; Sabatini, D.M. mTOR signaling at a glance. J. Cell Sci. 2009, 122, 3589–3594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Huang, S. Role of mTOR Signaling in Tumor Cell Motility, Invasion and Metastasis. Curr. Protein Pept. Sci. 2011, 12, 30–42. [Google Scholar] [PubMed]
- Circu, M.L.; Aw, T.Y. Glutathione and modulation of cell apoptosis. Biochim. Biophys. Acta 2012, 1823, 1767–1777. [Google Scholar] [CrossRef] [Green Version]
- Dalton, T.P.; Chen, Y.; Schneider, S.N.; Nebert, D.W.; Shertzer, H.G. Genetically altered mice to evaluate glutathione homeostasis in health and disease. Free Radic. Biol. Med. 2004, 37, 1511–1526. [Google Scholar] [CrossRef] [PubMed]
- Friesen, C.; Kiess, Y.; Debatin, K.M. A critical role of glutathione in determining apoptosis sensitivity and resistance in leukemia cells. Cell Death Differ. 2004, 11, S73–S85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cazanave, S.; Berson, A.; Haouzi, D.; Vadrot, N.; Fau, D.; Grodet, A.; Letteron, P.; Feldmann, G.; El-Benna, J.; Fromenty, B.; et al. High hepatic glutathione stores alleviate Fas-induced apoptosis in mice. J. Hepatol. 2007, 46, 858–868. [Google Scholar] [CrossRef]
- Armstrong, J.S.; Steinauer, K.K.; Hornung, B.; Irish, J.M.; Lecane, P.; Birrell, G.W.; Peehl, D.M.; Knox, S.J. Role of glutathione depletion and reactive oxygen species generation in apoptotic signaling in a human B lymphoma cell line. Cell Death Differ. 2002, 9, 252–263. [Google Scholar] [CrossRef] [Green Version]
- Saadat, N.; Liu, F.; Haynes, B.; Nangia-Makker, P.; Bao, X.; Li, J.; Polin, L.; Gupta, S.; Mao, G.; Shekhar, M.P. Nano-targeted Delivery of Rad6/Translesion Synthesis Inhibitor for Triple Negative Breast Cancer Therapy. Mol. Cancer Ther. 2018. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. Web-based inference of biological patterns, functions and pathways from metabolomic data using MetaboAnalyst. Nat. Protoc. 2011, 6, 743–760. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite Name | Mean ± SD (Control) | Mean ± SD (Treatment) | p-Value | Fold Changes Control/Trt |
| Amino Acids | ||||
| Aspartate | 102.3 ± 11.9 | 55.9 ± 4.7 | 0.0016 | 1.8 |
| Glutamate | 80.8 ± 7.9 | 48.7 ± 4.7 | 0.0019 | 1.7 |
| Leucine | 33.7 ± 4.1 | 17 ± 3.7 | 0.0030 | 2.0 |
| Glycine | 33.1 ± 1.2 | 20.4 ± 4.2 | 0.0035 | 1.6 |
| Alanine | 31 ± 1.4 | 19.8 ± 3.9 | 0.0045 | 1.6 |
| Glutamine | 99.9 ± 6.7 | 64.7 ± 13.3 | 0.0073 | 1.5 |
| Histidine | 54 ± 8.4 | 85.9 ± 31.3 | 0.0815 | 0.6 |
| Asparagine | 116.9 ± 16.2 | 54.5 ± 13.1 | 0.0033 | 2.1 |
| Taurine | 90.3 ± 19.9 | 78.2 ± 26.8 | 0.2822 | 1.2 |
| Valine | 23.8 ± 1.4 | 21.6 ± 6.3 | 0.2878 | 1.1 |
| Tryptophan | 81.3 ± 15 | 72.7 ± 28.7 | 0.3340 | 1.1 |
| Proline | 51.9 ± 49.3 | 63.7 ± 25.7 | 0.3659 | 0.8 |
| Lysine | 41.6 ± 22.8 | 37.2 ± 6.1 | 0.4075 | 1.1 |
| Isoleucine | 31.5 ± 9.9 | 30.6 ± 7 | 0.4499 | 1.0 |
| Methionine | 5.8 ± 5.3 | 5.5 ± 3.4 | 0.4653 | 1.1 |
| Arginine | nd | nd | ||
| Intermediate of TCA Cycle and Energy Metabolism | ||||
| Lactate | 138.5 ± 5.6 | 99.9 ± 3.6 | 0.0003 | 1.4 |
| 2-Oxoglutarate | 43.6 ± 3.3 | 29.3 ± 4.7 | 0.0061 | 1.5 |
| AMP | 32.1 ± 5 | 45 ± 1.7 | 0.0063 | 0.7 |
| Glutaric acid monomethyl ester | 17.8 ± 6.4 | 34 ± 2.8 | 0.0077 | 0.5 |
| Malate | 90.2 ± 10.7 | 48.7 ± 10.3 | 0.0111 | 1.9 |
| Succinate | 9.3 ± 2.6 | 5.2 ± 2.8 | 0.0645 | 1.8 |
| Glucose | 119.1 ± 53.4 | 187.3 ± 63.7 | 0.1139 | 0.6 |
| ADP | 47.8 ± 8.3 | 40.8 ± 4.8 | 0.1370 | 1.2 |
| Citrate | 42.4 ± 3.8 | 35.6 ± 11.6 | 0.1959 | 1.2 |
| NADH | 38.4 ± 3.5 | 43.4 ± 16 | 0.3040 | 0.9 |
| NADPH | 47 ± 6.3 | 51.3 ± 12.5 | 0.3118 | 0.9 |
| ATP | 42.2 ± 5.4 | 42.9 ± 11.3 | 0.4653 | 1.0 |
| Nucleic acid Associataed Metabolites | ||||
| Uracil | 98 ± 14.1 | 60.1 ± 24 | 0.0387 | 1.6 |
| UDP-N-Acetylglucosamine | 6.9 ± 2.1 | 3.9 ± 3.4 | 0.1266 | 1.8 |
| Other | ||||
| Glutathione | 69.6 ± 2.1 | 41.7 ± 6.7 | 0.0011 | 1.7 |
| Citrulline | 81.9 ± 5.1 | 63.9 ± 13 | 0.0438 | 1.3 |
| Cystine | 81.4 ± 6.3 | 58.4 ± 19 | 0.0582 | 1.4 |
| N-Acetylglucosamine | 21.9 ± 9.3 | 12.8 ± 5.2 | 0.1065 | 1.7 |
| Formate | 294.3 ± 68.5 | 312.8 ± 8.9 | 0.3334 | 0.9 |
| Fumarate | 25 ± 3.2 | 27.7 ± 5 | 0.2363 | 0.9 |
| Metabolite Name | Mean ± SD (Control) | Mean ± SD (Treatment) | p-Value | Fold Changes Control/Trt |
| Amino Acids | ||||
| Aspartate | 105.5 ± 3.5 | 77.4 ± 4.3 | 0.0010 | 1.4 |
| Glutamate | 80.1 ± 5.7 | 49.3 ± 6.2 | 0.0033 | 1.6 |
| Leucine | 31.8 ± 1.3 | 18.3 ± 0.8 | <0.0001 | 1.7 |
| Glycine | 28.2 ± 4.7 | 18.1 ± 3.2 | 0.0561 | 1.6 |
| Alanine | 28.8 ± 2.2 | 18.2 ± 2.3 | 0.0044 | 1.6 |
| Glutamine | 75.3 ± 5.1 | 53.7 ± 8.4 | 0.0177 | 1.4 |
| Histidine | ND | ND | ||
| Asparagine | 105 ± 21 | 84 ± 23.3 | 0.1986 | 1.3 |
| Taurine | ND | ND | ||
| Valine | 28.8 ± 4.9 | 21.7 ± 5.6 | 0.1706 | 1.3 |
| Tryptophan | 36.8 ± 2 | 17.8 ± 11.4 | 0.0401 | 2.1 |
| Proline | 90.2 ± 39.3 | 74.3 ± 34.9 | 0.3453 | 1.2 |
| Lysine | 38.8 ± 11.3 | 19.4 ± 7.1 | 0.0547 | 2 |
| Isoleucine | 37.2 ± 4.9 | 23.8 ± 2.7 | 0.0138 | 1.6 |
| Methionine | 8.7 ± 0.8 | 6.7 ± 1.9 | 0.1247 | 1.3 |
| Arginine | 43.8 ± 2.7 | 28.4 ± 6.6 | 0.0189 | 1.5 |
| Intermediate of TCA Cycle and Energy Metabolism | ||||
| Lactate | 125.8 ± 7.3 | 122 ± 15.4 | 0.3857 | 1 |
| 2-Oxoglutarate | 32.5 ± 7.9 | 17.2 ± 1.5 | 0.0272 | 1.9 |
| AMP | 27.5 ± 0.2 | 13.7 ± 2 | 0.0003 | 2 |
| Glutaric acid monomethyl ester | 27.4 ± 0 | 20.6 ± 7.4 | 1.3 | |
| Malate | 130.9 ± 7.8 | 84.7 ± 9 | 0.0027 | 1.5 |
| Succinate | 13.9 ± 1.7 | 5.3 ± 3.8 | 0.0215 | 2.6 |
| Glucose | 196.4 ± 50.1 | 147.1 ± 19.4 | 0.1324 | 1.3 |
| ADP | 33.6 ± 5.1 | 14.9 ± 7.7 | 0.0227 | 2.3 |
| Citrate | 35.2 ± 0.8 | 25.6 ± 4.3 | 0.0183 | 1.4 |
| NADH | 65.3 ± 11.7 | 43.7 ± 30.7 | 0.2024 | 1.5 |
| NADPH | 48.6 ± 11.1 | 38.1 ± 23.5 | 0.2996 | 1.3 |
| ATP | 43.5 ± 7.8 | 22.2 ± 5.5 | 0.0171 | 2 |
| Nucleic acid Associated Metabolites | ||||
| Uracil | 88.5 ± 11.9 | 40.2 ± 16.3 | 0.0139 | 2.2 |
| UDP-N-Acetylglucosamine | ND | |||
| Other | ||||
| Glutathione | 42.3 ± 4.5 | 28 ± 6.5 | 0.0319 | 1.5 |
| Citrulline | 65.4 ± 20.6 | 53.4 ± 25.4 | 0.3156 | 1.2 |
| Cystine | 61 ± 7.2 | 26.3 ± 14.1 | 0.0338 | 2.3 |
| N-Acetylglucosamine | ||||
| Fumarate | ||||
| Formate | 354.5 ± 90.9 | 346.7 ± 41 | 0.4585 | 1 |
| Tyrosine | 12.9 ± 0.6 | 67.8 ± 9.1 | 0.0134 | 0.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajasinghe, L.D.; Hutchings, M.; Gupta, S.V. Delta-Tocotrienol Modulates Glutamine Dependence by Inhibiting ASCT2 and LAT1 Transporters in Non-Small Cell Lung Cancer (NSCLC) Cells: A Metabolomic Approach. Metabolites 2019, 9, 50. https://doi.org/10.3390/metabo9030050
Rajasinghe LD, Hutchings M, Gupta SV. Delta-Tocotrienol Modulates Glutamine Dependence by Inhibiting ASCT2 and LAT1 Transporters in Non-Small Cell Lung Cancer (NSCLC) Cells: A Metabolomic Approach. Metabolites. 2019; 9(3):50. https://doi.org/10.3390/metabo9030050
Chicago/Turabian StyleRajasinghe, Lichchavi Dhananjaya, Melanie Hutchings, and Smiti Vaid Gupta. 2019. "Delta-Tocotrienol Modulates Glutamine Dependence by Inhibiting ASCT2 and LAT1 Transporters in Non-Small Cell Lung Cancer (NSCLC) Cells: A Metabolomic Approach" Metabolites 9, no. 3: 50. https://doi.org/10.3390/metabo9030050