Chronic Alcohol Dysregulates Skeletal Muscle Myogenic Gene Expression after Hind Limb Immobilization in Female Rats

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
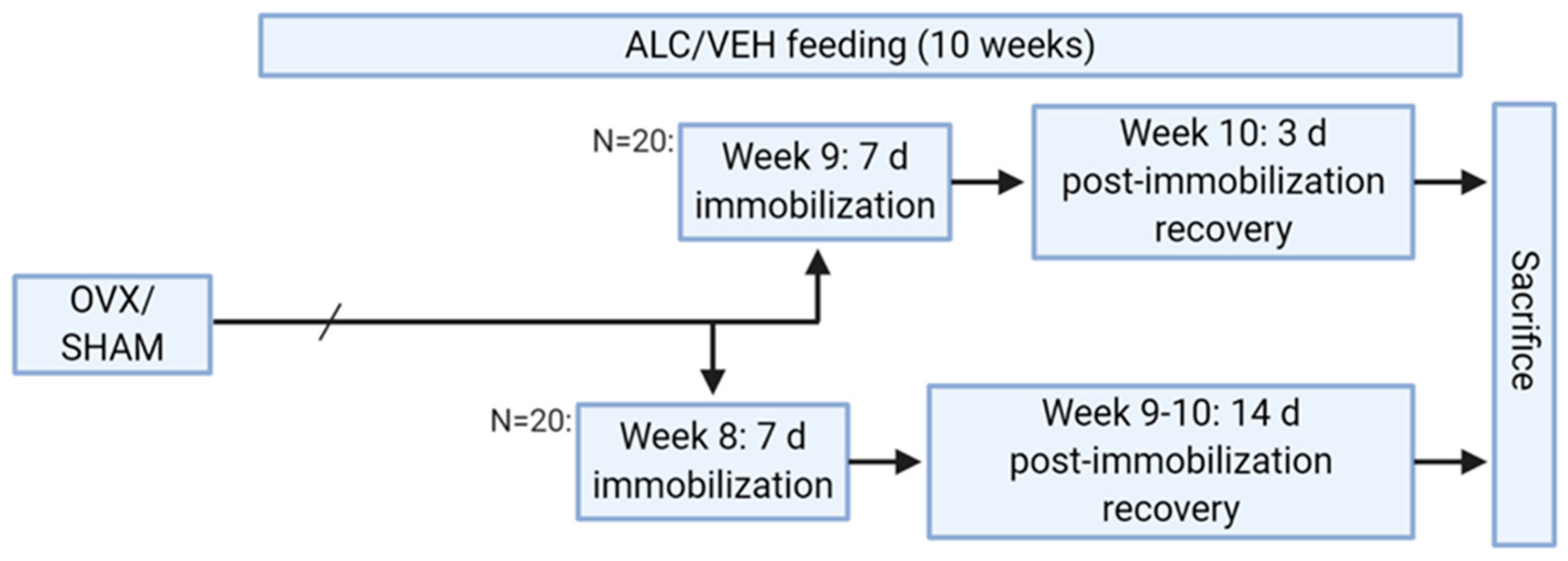
2.1. Animal Study Design
2.2. Ovariectomy
2.3. Alcohol Feeding
2.4. Unilateral Hind Limb Immobilization
2.5. RNA Isolation and qPCR
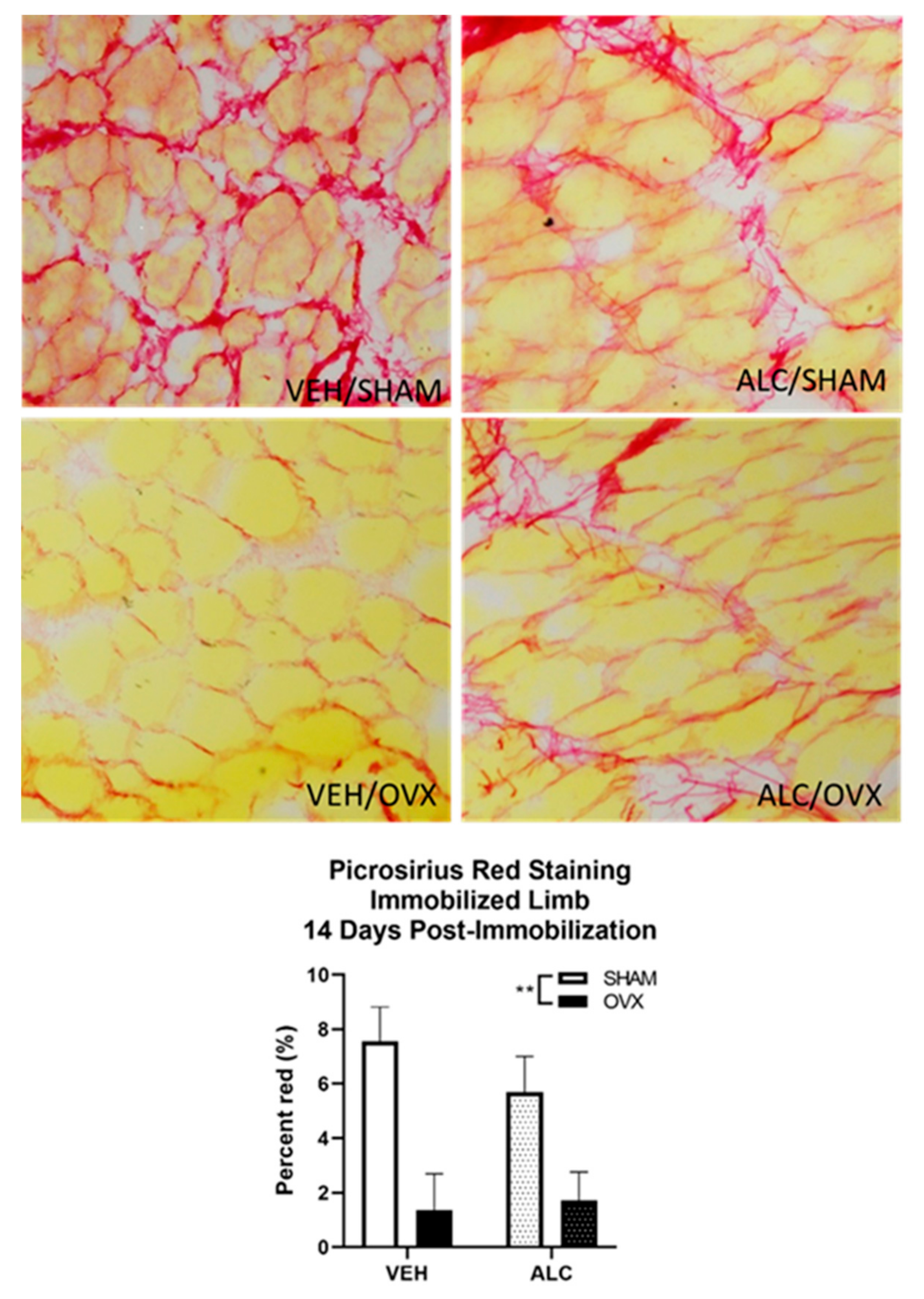
2.6. Picrosirius Staining for Collagen Expression
2.7. Statistical Analyses
3. Results
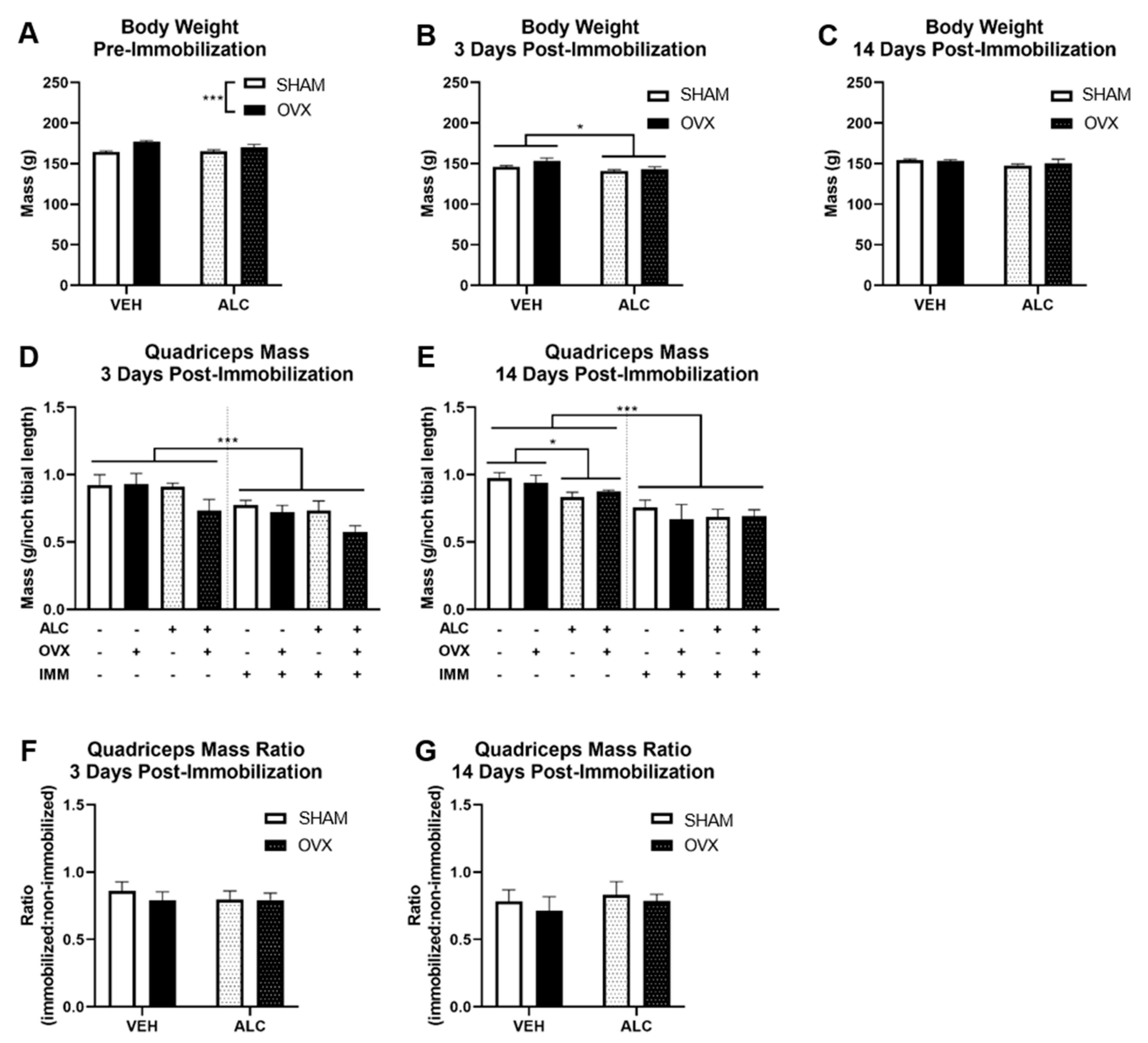
3.1. Body and Quadriceps Weight
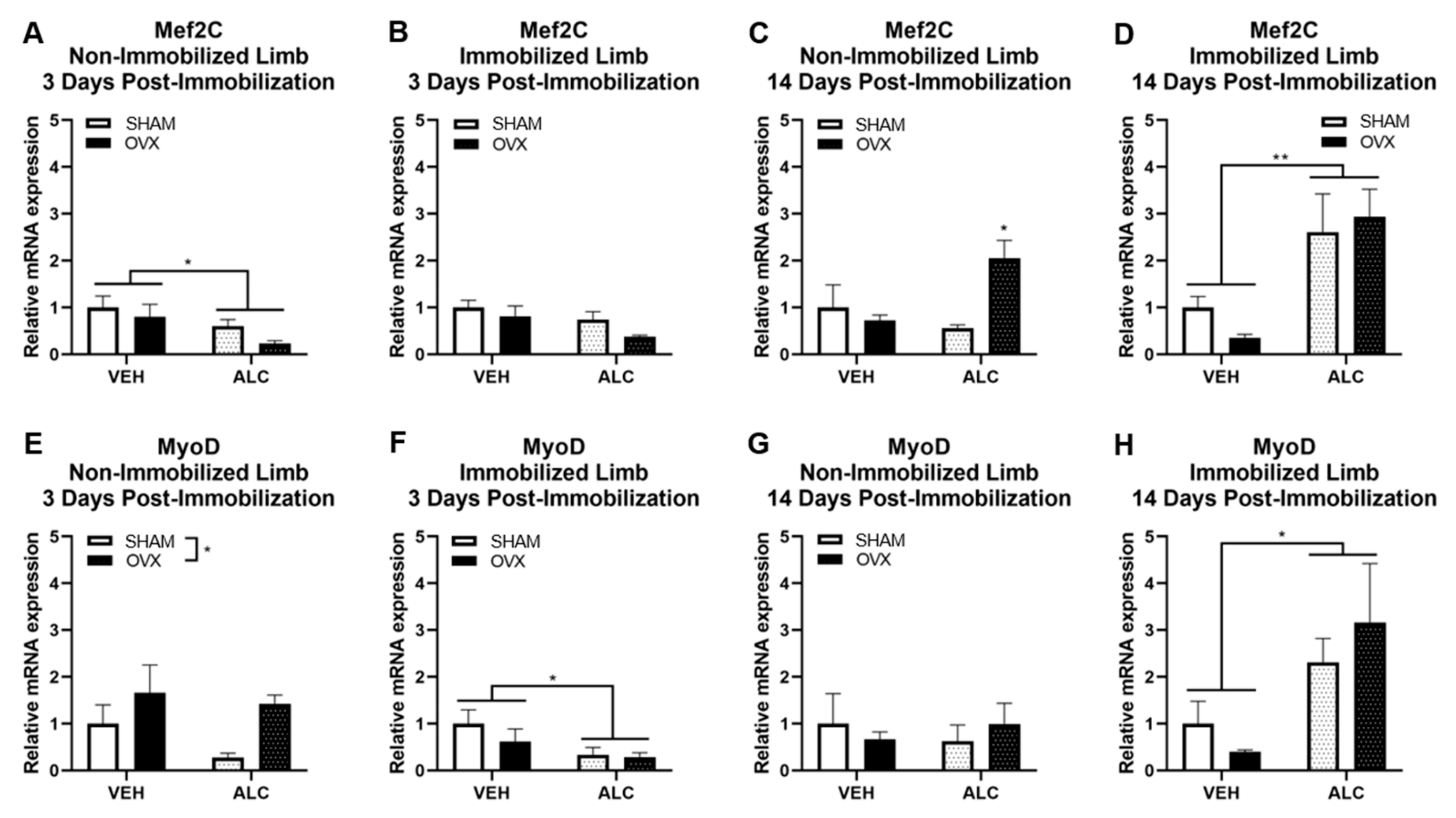
3.2. Myogenic Gene Expression
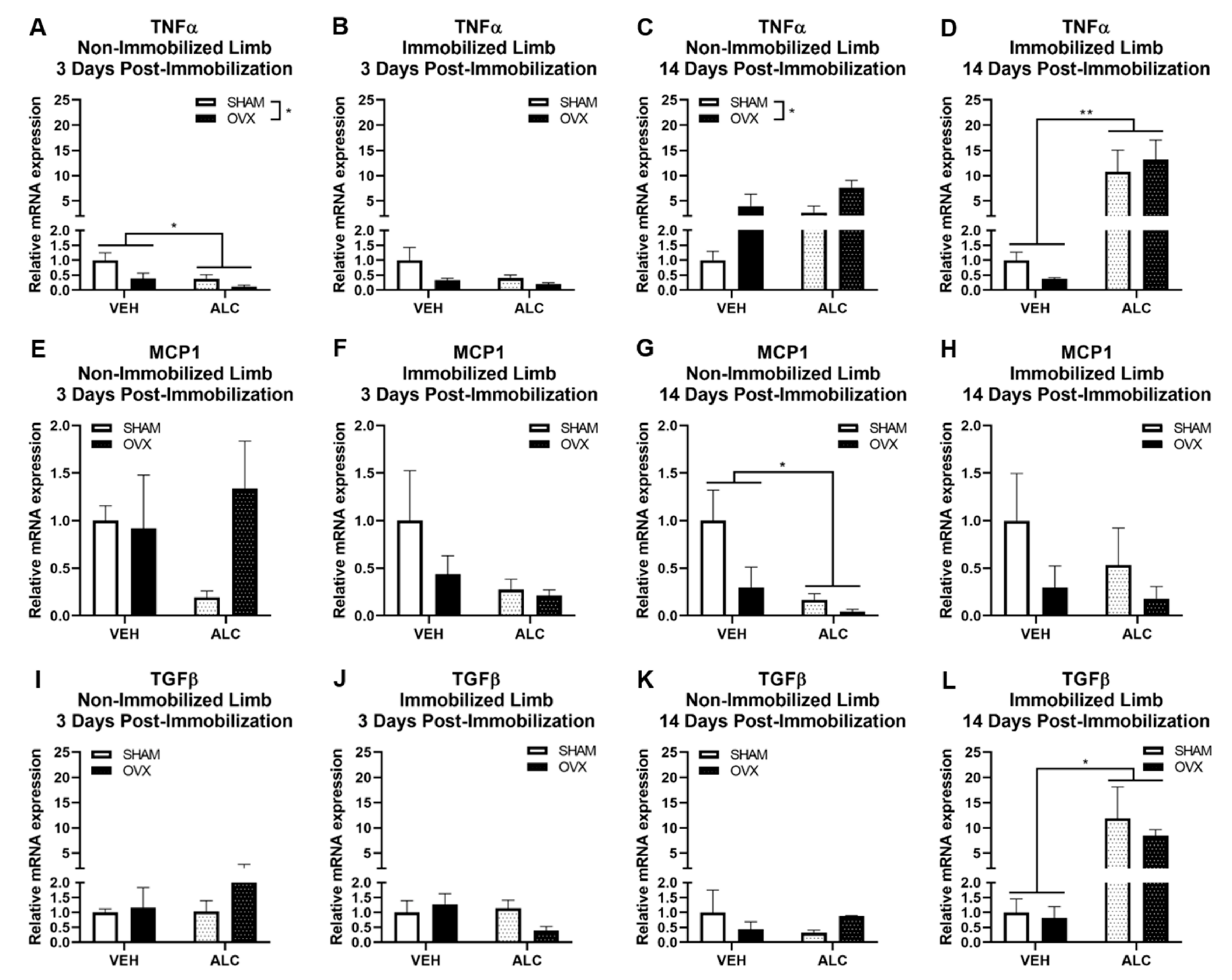
3.3. Inflammatory and Fibrotic Gene Expression
3.4. Collagen Expression
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Lukaszyk, C.; Harvey, L.; Sherrington, C.; Keay, L.; Tiedemann, A.; Coombes, J.; Clemson, L.; Ivers, R. Risk factors, incidence, consequences and prevention strategies for falls and fall-injury within older indigenous populations: a systematic review. Aust. New Zealand J. Public Heal. 2016, 40, 564–568. [Google Scholar] [CrossRef]
- Frech, T.; Ma, K.-N.; Ferrucci, E.D.; Lanier, A.P.; McFadden, M.; Tom-Orme, L.; Slattery, M.L.; Murtaugh, M. Prevalence of fracture and osteoporosis risk factors in American Indian and Alaska Native people. J. Heal. Care Poor Underserved 2012, 23, 1157–1173. [Google Scholar] [CrossRef] [Green Version]
- DeKeyser, G.J.; Clary, C.R.; Otis, J.S. Chronic alcohol ingestion delays skeletal muscle regeneration following injury. Regen. Med. Res. 2013, 1, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, H.; Chin, A.-V.; Cunningham, C.; Lawlor, B. Alcohol use disorders in elderly people—redefining an age old problem in old age. BMJ 2003, 327, 664–667. [Google Scholar] [CrossRef] [PubMed]
- Honkanen, R.; Visuri, T. Blood alcohol levels in a series of injured patients with special reference to accident and type of injury. Ann. Chir. Gynaecol. 1976, 65, 287–294. [Google Scholar] [PubMed]
- Felson, D.T.; Kiel, D.P.; Anderson, J.J.; Kannel, W.B. Alcohol Consumption And Hip Fractures: The Framingham Study. Am. J. Epidemiol. 1988, 128, 1102–1110. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Avila, M.; Colditz, G.; Stampfer, M.J.; Rosner, B.; Speizer, F.E.; Willett, W.C. Caffeine, moderate alcohol intake, and risk of fractures of the hip and forearm in middle-aged women. Am. J. Clin. Nutr. 1991, 54, 157–163. [Google Scholar] [CrossRef]
- Høidrup, S.; Grønbæk, M.; Gottschau, A.; Lauritzen, J.B.; Schroll, M. Alcohol Intake, Beverage Preference, and Risk of Hip Fracture in Men and Women. Am. J. Epidemiol. 1999, 149, 993–1001. [Google Scholar] [CrossRef] [Green Version]
- Baron, J.; Farahmand, B.; Weiderpass, E.; Michaëlsson, K.; Alberts, A.; Persson, I.; Ljunghall, S. Cigarette smoking, alcohol consumption, and risk of hip fracture in women. Arch. Intern. Med. 2001, 161, 983–988. [Google Scholar] [CrossRef] [Green Version]
- Kaukonen, J.-P.; Nurmi-Lüthje, I.; Lüthje, P.; Naboulsi, H.; Tanninen, S.; Kataja, M.; Kallio, M.-L.; Leppilampi, M. Acute alcohol use among patients with acute hip fractures: A descriptive incidence study in southeastern Finland. Alcohol Alcohol. 2006, 41, 345–348. [Google Scholar] [CrossRef] [Green Version]
- Orces, C. Emergency department visits for fall-related fractures among older adults in the USA: a retrospective cross-sectional analysis of the National Electronic Injury Surveillance System All Injury Program, 2001–2008. BMJ Open 2013, 3, e001722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ka, H.; Van Lieshout, E.M.M.; Polinder, S.; Panneman, M.J.; Van Der Cammen, T.J.; Patka, P. Rapid Increase in Hospitalizations Resulting from Fall-Related Traumatic Head Injury in Older Adults in the Netherlands 1986–2008. J. Neurotrauma 2011, 28, 739–744. [Google Scholar]
- Looker, A.C.; Melton, L.J.; Harris, T.B.; Borrud, L.G.; Shepherd, J.A. Prevalence and Trends in Low Femur Bone Density Among Older US Adults: NHANES 2005–2006 Compared With NHANES III. J. Bone Miner. Res. 2009, 25, 64–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syddall, H.; Roberts, H.C.; Evandrou, M.; Cooper, C.; Bergman, H.; Sayer, A.A. Prevalence and correlates of frailty among community-dwelling older men and women: findings from the Hertfordshire Cohort Study. Age Ageing 2009, 39, 197–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsiaras, A.; Newman, A.B.; Kriska, A.; Brach, J.S.; Krishnaswami, S.; Feingold, E.; Kritchevsky, S.; Li, R.; Harris, T.B.; Schwartz, A.; et al. Skeletal muscle fatigue, strength, and quality in the elderly: the Health ABC Study. J. Appl. Physiol. 2005, 99, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Solà, J.; Preedy, V.R.; Lang, C.H.; Gonzalez-Reimers, E.; Arno, M.; Lin, J.C.I.; Wiseman, H.; Zhou, S.; Emery, P.W.; Nakahara, T.; et al. Molecular and Cellular Events in Alcohol-Induced Muscle Disease. Alcohol. Clin. Exp. Res. 2007, 31, 1953–1962. [Google Scholar] [CrossRef] [PubMed]
- Preedy, V.R.; Ohlendieck, K.; Adachi, J.; Koll, M.; Sneddon, A.; Hunter, R.; Rajendram, R.; Mantle, D.; Peters, T.J. The importance of alcohol-induced muscle disease. J. Muscle Res. Cell Motil. 2003, 24, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Junqué, A.; Estruch, R.; Nicolás, J.M.; Villegas, E.; Urbano-Márquez, A. Relationship Between Ethanol-Related Diseases And Nutritional Status In Chronically Alcoholic Men. Alcohol Alcohol. 1993, 28, 543–550. [Google Scholar]
- Urbano-Márquez, A.; Fernández-Solà, J. Effects of alcohol on skeletal and cardiac muscle. Muscle Nerve 2004, 30, 689–707. [Google Scholar] [CrossRef]
- Simon, L.; Jolley, S.E.; Molina, P.E. Alcoholic Myopathy: Pathophysiologic Mechanisms and Clinical Implications. Alcohol Res. Curr. Rev. 2017, 38, 207–217. [Google Scholar]
- González-Reimers, E.; Fernández-Rodríguez, C.M.; Santolaria-Fernández, F.; De La Vega-Prieto, M.J.; Martín-González, C.; Gómez-Rodríguez, M.Á.; Alemán-Valls, M.; Rodríguez-Gaspar, M. Interleukin-15 and Other Myokines in Chronic Alcoholics. Alcohol Alcohol. 2011, 46, 529–533. [Google Scholar] [CrossRef] [Green Version]
- Steiner, J.L.; Lang, C.H. Dysregulation of skeletal muscle protein metabolism by alcohol. Am. J. Physiol. Metab. 2015, 308, E699–E712. [Google Scholar] [CrossRef] [Green Version]
- Simon, L.; LeCapitaine, N.; Berner, P.; Stouwe, C.V.; Mussell, J.C.; Allerton, T.; Primeaux, S.D.; Dufour, J.; Nelson, S.; Bagby, G.J.; et al. Chronic binge alcohol consumption alters myogenic gene expression and reduces in vitro myogenic differentiation potential of myoblasts from rhesus macaques. Am. J. Physiol. Integr. Comp. Physiol. 2014, 306, R837–R844. [Google Scholar] [CrossRef] [PubMed]
- Simon, L.; Ford, S.M.; Song, K.; Berner, P.; Stouwe, C.V.; Nelson, S.; Bagby, G.J.; Molina, P.E. Decreased myoblast differentiation in chronic binge alcohol-administered simian immunodeficiency virus-infected male macaques: role of decreased miR-206. Am. J. Physiol. Integr. Comp. Physiol. 2017, 313, R240–R250. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.L.; Muñoz-Cánoves, P. Regulation and dysregulation of fibrosis in skeletal muscle. Exp. Cell Res. 2010, 316, 3050–3058. [Google Scholar] [CrossRef]
- Grounds, M.D.; Sorokin, L.; White, J.D. Strength at the extracellular matrix-muscle interface. Scand. J. Med. Sci. Sports 2005, 15, 381–391. [Google Scholar] [CrossRef]
- Kaariainen, M.; Jarvinen, T.; Jarvinen, M.; Rantanen, J.; Kalimo, H. Relation between myofibers and connective tissue during muscle injury repair. Scand. J. Med. Sci. Sports 2000, 10, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Dodd, T.; Simon, L.; LeCapitaine, N.J.; Zabaleta, J.; Mussell, J.; Berner, P.; Ford, S.; Dufour, J.; Bagby, G.J.; Nelson, S.; et al. Chronic binge alcohol administration accentuates expression of pro-fibrotic and inflammatory genes in the skeletal muscle of simian immunodeficiency virus-infected macaques. Alcohol. Clin. Exp. Res. 2014, 38, 2697–2706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, R.; Lang, C.H. Alcohol Accelerates Loss of Muscle and Impairs Recovery of Muscle Mass Resulting From Disuse Atrophy. Alcohol. Clin. Exp. Res. 2007, 32, 128–137. [Google Scholar] [CrossRef]
- Jones, J.H. Resource Book for the Design of Animal Exercise Protocols. Am. J. Veter- Res. 2007, 68, 583. [Google Scholar] [CrossRef]
- Chacon-Cabrera, A.; Lund-Palau, H.; Gea, J.; Barreiro, E. Time-Course of Muscle Mass Loss, Damage, and Proteolysis in Gastrocnemius following Unloading and Reloading: Implications in Chronic Diseases. PLoS ONE 2016, 11, e0164951. [Google Scholar] [CrossRef] [PubMed]
- Chacon-Cabrera, A.; Gea, J.; Barreiro, E. Short- and Long-Term Hindlimb Immobilization and Reloading: Profile of Epigenetic Events in Gastrocnemius. J. Cell. Physiol. 2016, 232, 1415–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sitnick, M.; Foley, A.M.; Brown, M.; Spangenburg, E.E. Ovariectomy prevents the recovery of atrophied gastrocnemius skeletal muscle mass. J. Appl. Physiol. 2006, 100, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.; Foley, A.; Ferreria, J.A. Ovariectomy, hindlimb unweighting, and recovery effects on skeletal muscle in adult rats. Aviat. Space Environ. Med. 2005, 76, 1012–1018. [Google Scholar]
- Robichaux, S.; Lacour, N.; Bagby, G.J.; Amedee, A.M. Validation of RPS13 as a reference gene for absolute quantification of SIV RNA in tissue of rhesus macaques. J. Virol. Methods 2016, 236, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Benyam Yoseph, S.S. Redefining the satellite cell as the motor of skeletal muscle regeneration. J. Sci. Appl. Biomed. 2015, 5, 76–82. [Google Scholar]
- Ye, F.; Mathur, S.; Liu, M.; Borst, S.E.; Walter, G.A.; Sweeney, H.L.; Vandenborne, K. Overexpression of IGF-1 attenuates skeletal muscle damage and accelerates muscle regeneration and functional recovery after disuse. Exp. Physiol. 2013, 98, 1038–1052. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.V.; Hughes, S.M. Mef2 and the skeletal muscle differentiation program. Semin. Cell Dev. Boil. 2017, 72, 33–44. [Google Scholar] [CrossRef]
- Adler, K.; Molina, P.E.; Simon, L. Epigenomic mechanisms of alcohol-induced impaired differentiation of skeletal muscle stem cells; role of Class IIA histone deacetylases. Physiol. Genom. 2019, 51, 471–479. [Google Scholar] [CrossRef]
- Arya, M.A.; Tai, A.K.; Wooten, E.; Parkin, C.D.; Kudryavtseva, E.; Huggins, G. Notch Pathway Activation Contributes to Inhibition of C2C12 Myoblast Differentiation by Ethanol. PLoS ONE 2013, 8, e71632. [Google Scholar] [CrossRef] [Green Version]
- Guitart, M.; Lloreta, J.; García, L.M.; Barreiro, E. Muscle regeneration potential and satellite cell activation profile during recovery following hindlimb immobilization in mice. J. Cell. Physiol. 2018, 233, 4360–4372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shireman, P.K.; Contreras-Shannon, V.; Ochoa, O.; Karia, B.P.; Michalek, J.E.; McManus, L.M. MCP-1 deficiency causes altered inflammation with impaired skeletal muscle regeneration. J. Leukoc. Boil. 2006, 81, 775–785. [Google Scholar] [CrossRef]
- Li, Y.-P.; Schwartz, R.J. TNF-α regulates early differentiation of C2C12 myoblasts in an autocrine fashion. FASEB J. 2001, 15, 1413–1415. [Google Scholar] [CrossRef] [PubMed]
- Costamagna, D.; Costelli, P.; Sampaolesi, M.; Penna, F. Role of Inflammation in Muscle Homeostasis and Myogenesis. Mediat. Inflamm. 2015, 2015, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrianjafiniony, T.; Dupré-Aucouturier, S.; Letexier, M.; Couchoux, H.; Desplanches, M. Oxidative stress, apoptosis, and proteolysis in skeletal muscle repair after unloading. Am. J. Physiol. Physiol. 2010, 299, C307–C315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, J.; Hasser, E.M.; Brown, M. Effects of ovariectomy and hindlimb unloading on skeletal muscle. J. Appl. Physiol. 1998, 85, 1316–1321. [Google Scholar] [CrossRef] [Green Version]
- Kaasik, P.; Riso, E.-M.; Seene, T. Extracellular Matrix and Myofibrils During Unloading and Reloading of Skeletal Muscle. Int. J. Sports Med. 2011, 32, 247–253. [Google Scholar] [CrossRef]
- Heinemeier, K.M.; Olesen, J.; Haddad, F.; Schjerling, P.; Baldwin, K.M.; Kjaer, M. Effect of unloading followed by reloading on expression of collagen and related growth factors in rat tendon and muscle. J. Appl. Physiol. 2009, 106, 178–186. [Google Scholar] [CrossRef] [Green Version]
- Miller, T.A.; Lesniewski, L.A.; Muller-Delp, J.; Majors, A.K.; Scalise, D.; Delp, M. Hindlimb unloading induces a collagen isoform shift in the soleus muscle of the rat. Am. J. Physiol. Integr. Comp. Physiol. 2001, 281, R1710–R1717. [Google Scholar] [CrossRef]
- Fonseca, H.; Powers, S.K.; Gonçalves, D.; Santos, A.; Mota, M.P.; Duarte, J.A. Physical Inactivity is a Major Contributor to Ovariectomy-Induced Sarcopenia. Int. J. Sports Med. 2012, 33, 268–278. [Google Scholar] [CrossRef]
- Braggion, G.F.; Ornelas, E.; Cury, J.C.S.; Lima, N.E.A.; Aquino, R.C.; Fonseca, F.L.A.; Maifrino, L.B.M. Morphological and Biochemical Effects on the Skeletal Muscle of Ovariectomized Old Female Rats Submitted to the Intake of Diets with Vegetable or Animal Protein and Resistance Training. Oxidative Med. Cell. Longev. 2016, 2016, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, S.M.; Kazi, A.A.; Hong-Brown, L.; Lang, C.H. Delayed Recovery of Skeletal Muscle Mass following Hindlimb Immobilization in mTOR Heterozygous Mice. PLoS ONE 2012, 7, e38910. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| Myod | CAGGTGTAACCATACCC | CTGGCCAAGCAACTCTTAT |
| Mef2c | ATCTCTCCCTGCCTTCTAC | CGTGTGTTGTGGGTATCTC |
| Myh1 | CCGTGAACTTGAAGGAGAAG | CCTCTTCAGTTTGGTAAG |
| Myogenin | CCACCGTCCATTCACATAAG | GGACTCCATCTTTCTCTCCT |
| Mcp1 | CTCAGCCAGATGCAGTTAAT | CTGCTGGTGATTCTCTTGTAG |
| Tnfa | CGTGTTCATCCGTTCTCTAC | GAGCCACAATTCCCTTTCT |
| Il-1b | CTGACAGGCAACCACTTAC | CTGTGCACTGGTCCAAAT |
| Il-10 | CGACGCTGTCATCGATTT | GGCCTTGTAGACACCTTTG |
| Tgf b | GAACCAAGGAGACGGAATAC | GGGACTGATCCCATTGATTT |
| Rps13 | GCACCTTGAGAGGAACAGAA | GAGCACCCGCTTAGTCTTATAG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levitt, D.E.; Yeh, A.Y.; Prendergast, M.J.; Budnar, Jr., R.G.; Adler, K.A.; Cook, G.; Molina, P.E.; Simon, L. Chronic Alcohol Dysregulates Skeletal Muscle Myogenic Gene Expression after Hind Limb Immobilization in Female Rats. Biomolecules 2020, 10, 441. https://doi.org/10.3390/biom10030441
Levitt DE, Yeh AY, Prendergast MJ, Budnar, Jr. RG, Adler KA, Cook G, Molina PE, Simon L. Chronic Alcohol Dysregulates Skeletal Muscle Myogenic Gene Expression after Hind Limb Immobilization in Female Rats. Biomolecules. 2020; 10(3):441. https://doi.org/10.3390/biom10030441
Chicago/Turabian StyleLevitt, Danielle E., Alice Y. Yeh, Matthew J. Prendergast, Ronald G. Budnar, Jr., Katherine A. Adler, Garth Cook, Patricia E. Molina, and Liz Simon. 2020. "Chronic Alcohol Dysregulates Skeletal Muscle Myogenic Gene Expression after Hind Limb Immobilization in Female Rats" Biomolecules 10, no. 3: 441. https://doi.org/10.3390/biom10030441