
Chronic Binge Alcohol and Ovarian Hormone Loss Dysregulate Circulating Immune Cell SIV Co-Receptor Expression and Mitochondrial Homeostasis in SIV-Infected Rhesus Macaques

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
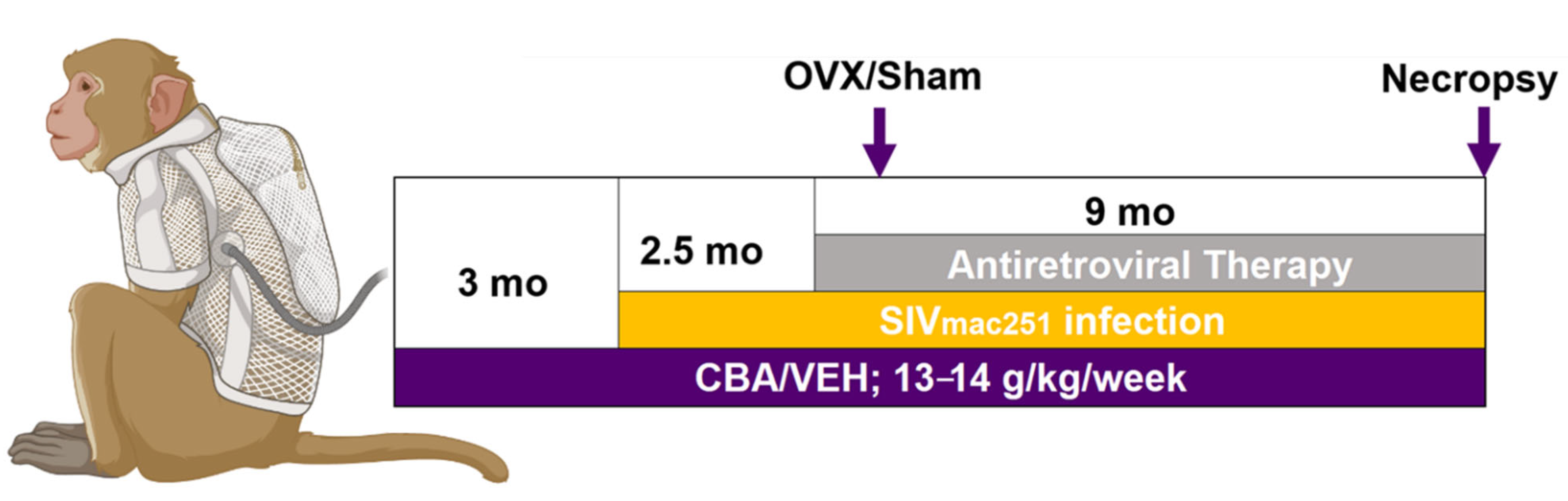
2.2. Animal Characteristics and Study Design
2.3. Plasma Viral Load and PBMC Viral DNA/RNA Measurements
2.4. RNA Isolation and Reverse Transcription Quantitative Polymerase Chain Reaction for Mitochondrial-Related Genes
2.5. Flow Cytometry Analysis
2.6. Statistical Analysis
3. Results
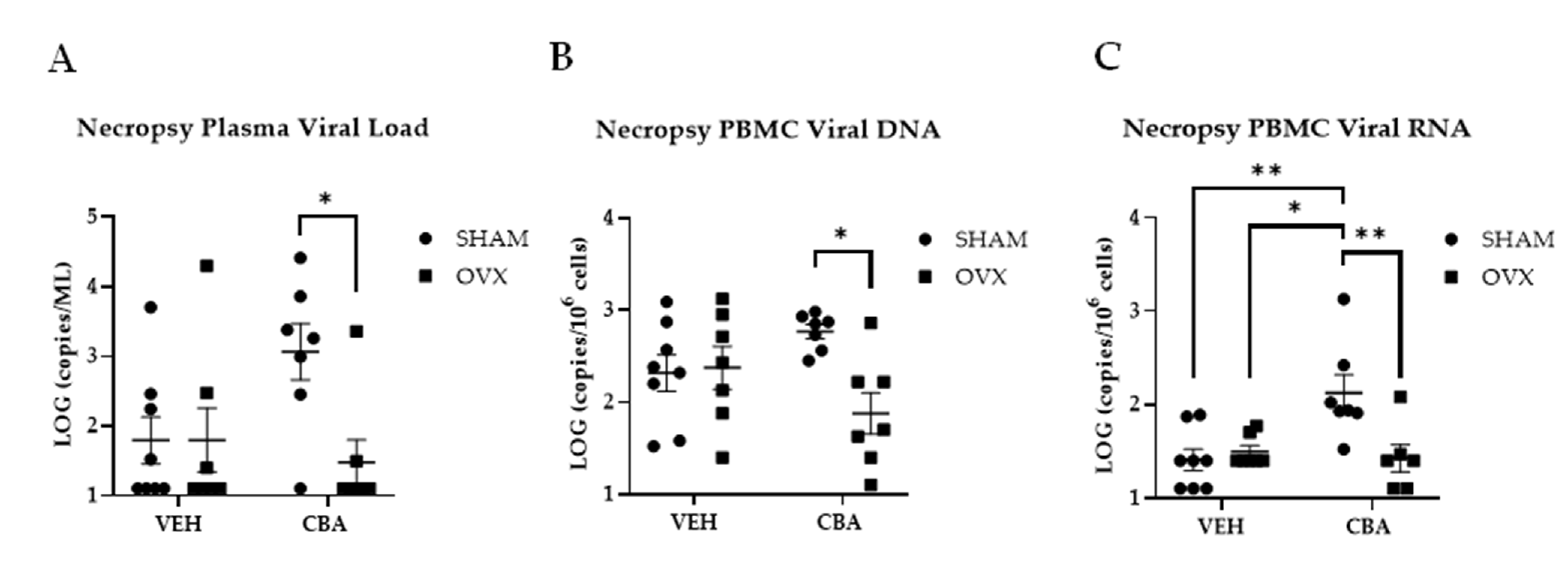
3.1. CBA/SHAM Animals Had Higher Plasma Viral Loads, and PBMC Viral DNA and RNA
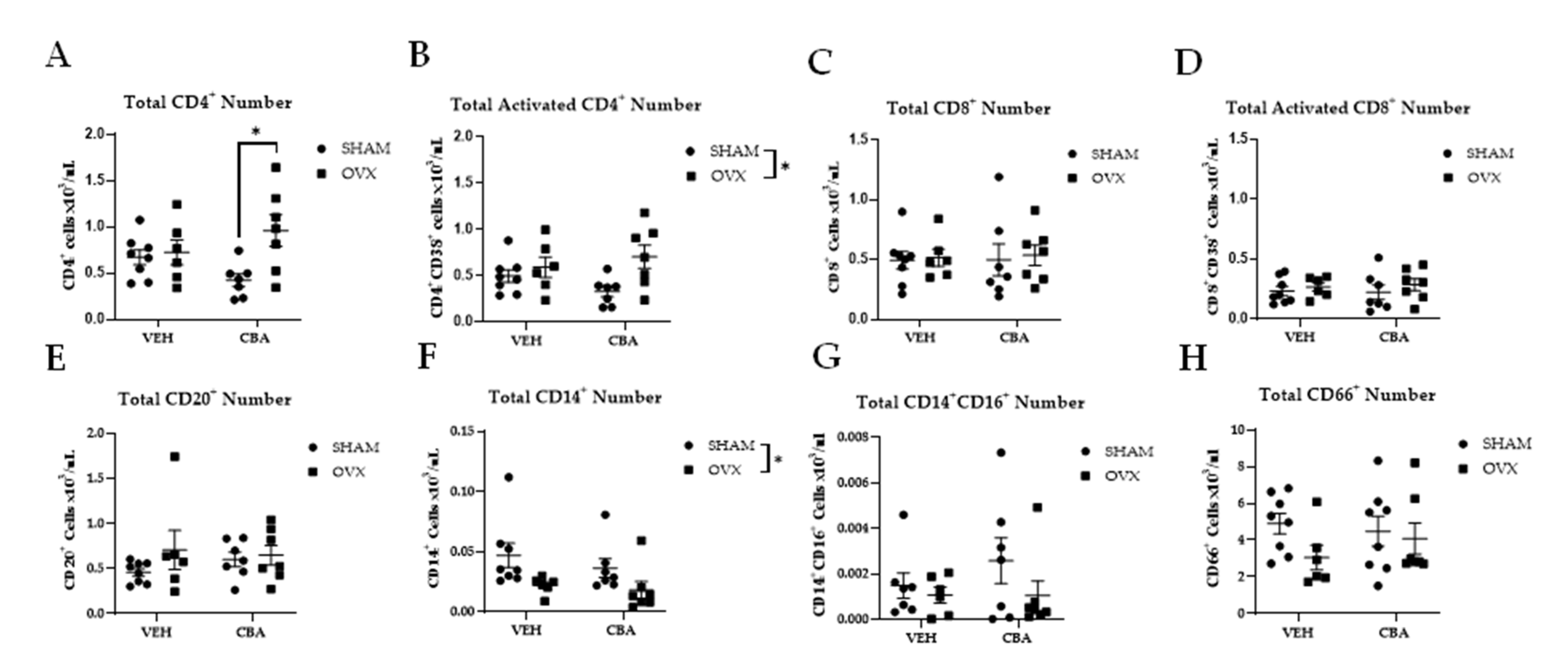
3.2. Immune Cell Phenotype
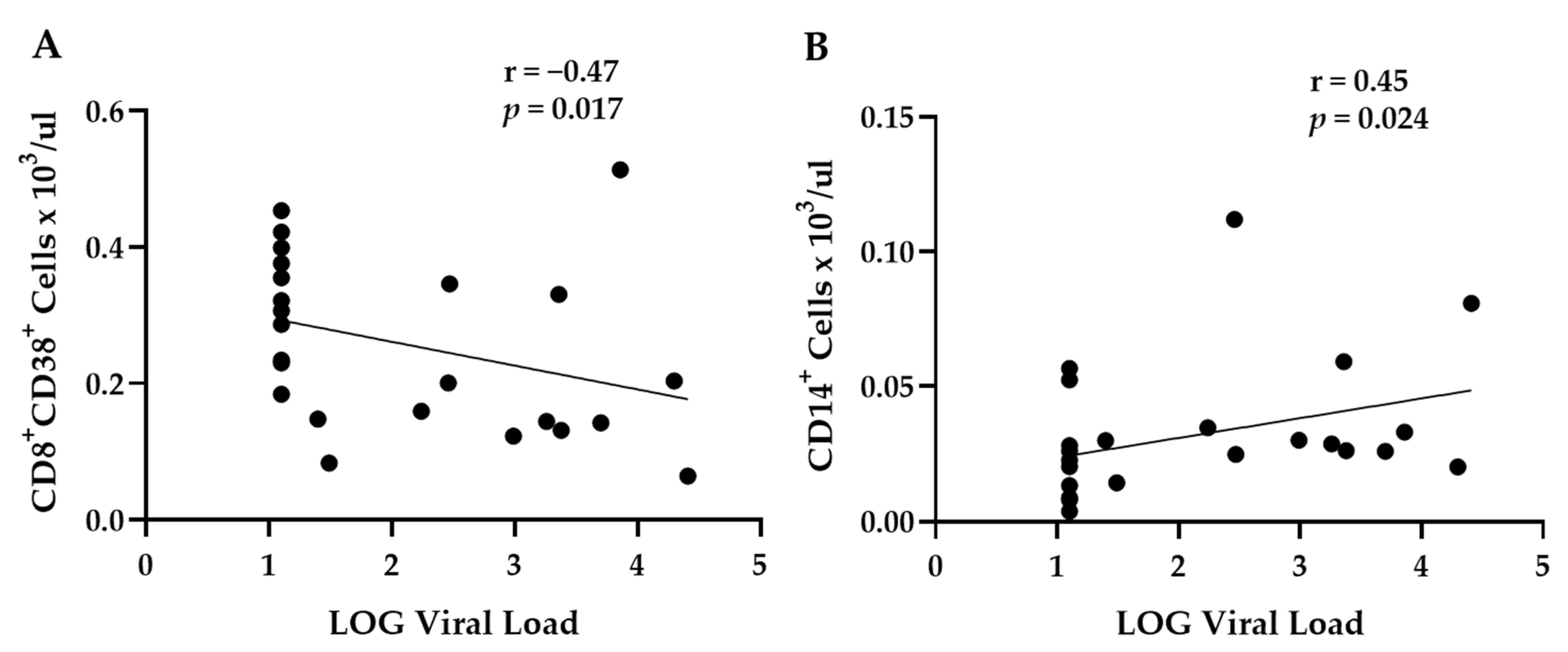
3.3. Correlation Analysis of Viral Data and Blood Cell Populations
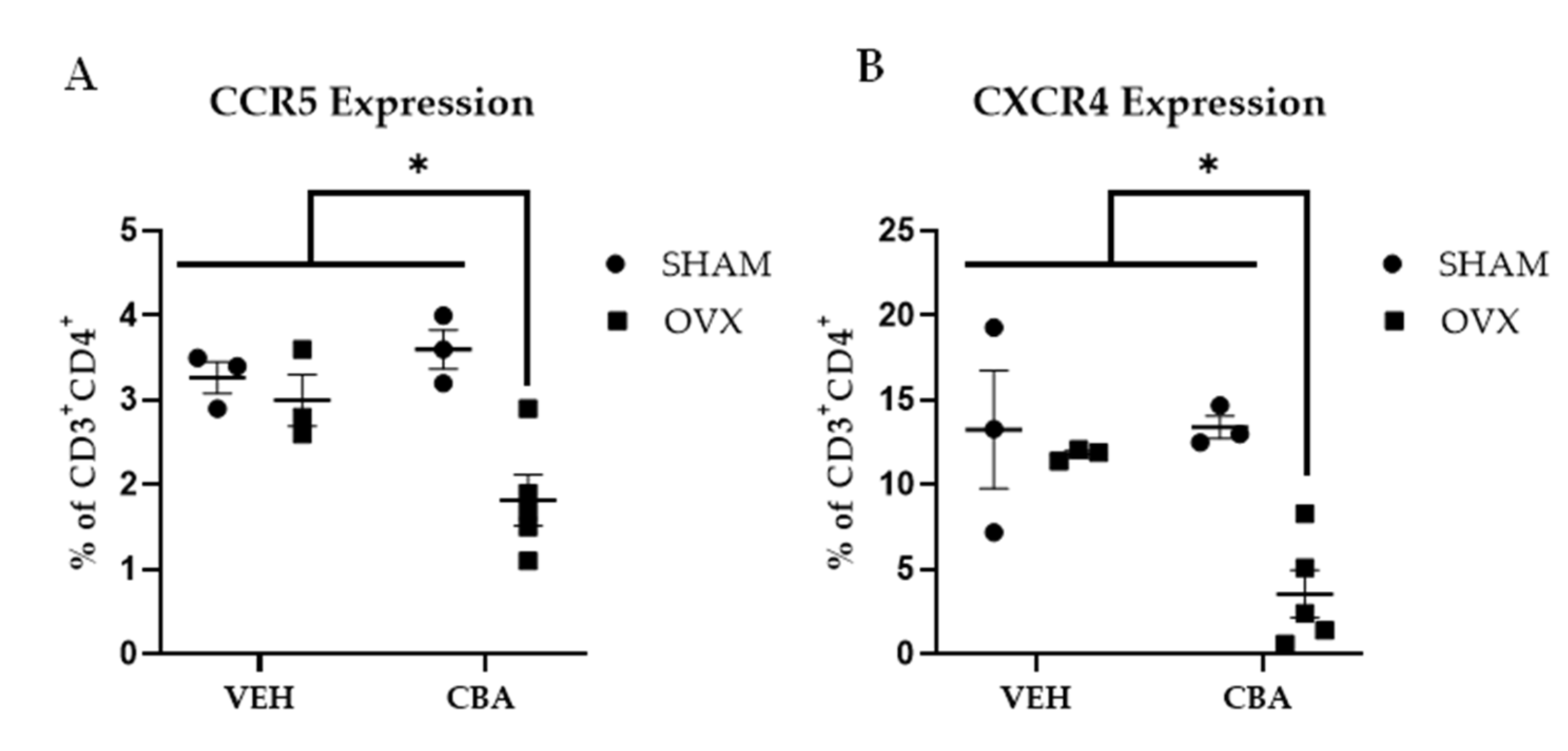
3.4. SIV Co-Receptor Expression
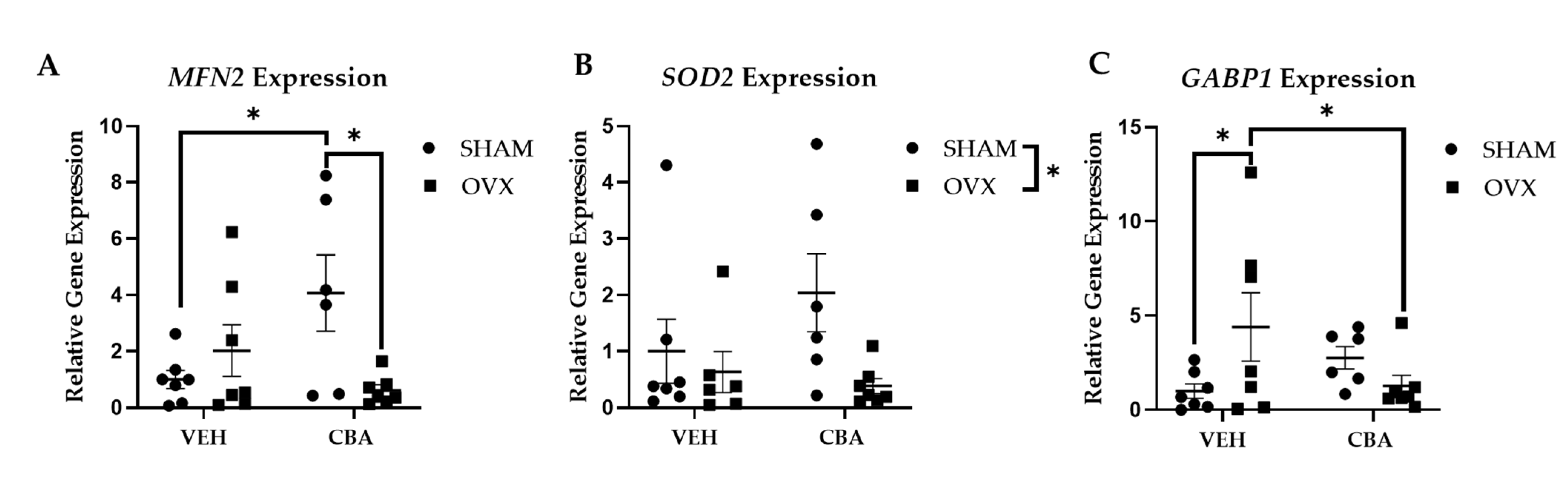
3.5. PBMC Mitochondrial-Related Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control and Prevention (CDC). HIV among People Aged 50 and Older. Available online: https://www.cdc.gov/hiv/group/age/olderamericans/index.html (accessed on 7 August 2021).
- HIV.gov. U.S. Statistics. Available online: https://www.hiv.gov/hiv-basics/overview/data-and-trends/statistics (accessed on 5 August 2021).
- Galvan, F.H.; Burnam, M.A.; Bing, E.G. Co-occurring Psychiatric Symptoms and Drug Dependence or Heavy Drinking Among HIV-Positive People. J. Psychoact. Drugs 2003, 35, 153–160. [Google Scholar] [CrossRef]
- Grant, B.F.; Dawson, D.A.; Stinson, F.S.; Chou, S.P.; Dufour, M.C.; Pickering, R.P. The 12-month prevalence and trends in DSM-IV alcohol abuse and dependence: United States, 1991–1992 and 2001–2002. Drug Alcohol Depend. 2004, 74, 223–234. [Google Scholar] [CrossRef]
- Grant, B.F.; Goldstein, R.B.; Saha, T.D.; Chou, S.P.; Jung, J.; Zhang, H.; Pickering, R.P.; Ruan, W.J.; Smith, S.M.; Huang, B.; et al. Epidemiology of DSM-5 Alcohol Use Disorder: Results from the National Epidemiologic Survey on Alcohol and Related Conditions III. JAMA Psychiatry 2015, 72, 757–766. [Google Scholar] [CrossRef] [Green Version]
- Molina, P.E.; Simon, L.; Amedee, A.M.; Welsh, D.A.; Ferguson, T.F. Impact of Alcohol on HIV Disease Pathogenesis, Comorbidities and Aging: Integrating Preclinical and Clinical Findings. Alcohol Alcohol. 2018, 53, 439–447. [Google Scholar] [CrossRef]
- Ferguson, T.F.; Theall, K.P.; Brashear, M.; Maffei, V.; Beauchamp, A.; Siggins, R.W.; Simon, L.; Mercante, D.; Nelson, S.; Welsh, D.A.; et al. Comprehensive Assessment of Alcohol Consumption in People Living with HIV (PLWH): The New Orleans Alcohol Use in HIV Study. Alcohol. Clin. Exp. Res. 2020, 44, 1261–1272. [Google Scholar] [CrossRef]
- Register, T.C.; Cline, J.M.; Shively, C.A. Health Issues in Postmenopausal Women Who Drink. Alcohol Res. Health J. Natl. Inst. Alcohol Abus. Alcohol. 2002, 26, 299–307. [Google Scholar]
- De Pommerol, M.; Hessamfar, M.; Lawson-Ayayi, S.; Neau, D.; Geffard, S.; Farbos, S.; Uwamaliya, B.; Vandenhende, M.-A.; Pellegrin, J.-L.; Blancpain, S.; et al. Menopause and HIV infection: Age at onset and associated factors, ANRS CO3 Aquitaine cohort. Int. J. STD AIDS 2011, 22, 67–72. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). HIV and Women: HIV Incidence. Available online: https://www.cdc.gov/hiv/group/gender/women/incidence.html (accessed on 31 August 2021).
- Bull, L.; Tittle, V.; Rashid, T.; Nwokolo, N. HIV and the menopause: A review. Post Reprod. Health 2017, 24, 19–25. [Google Scholar] [CrossRef]
- Simon, L.; Torres, D.; Saravia, A.; Levitt, D.E.; Stouwe, C.V.; Mc Garrah, H.; Coleman, L.; Dufour, J.P.; Amedee, A.M.; Molina, P.E. Chronic binge alcohol and ovariectomy-mediated impaired insulin responsiveness in SIV-infected female rhesus macaques. Am. J. Physiol. Integr. Comp. Physiol. 2021, 321, R699–R711. [Google Scholar] [CrossRef] [PubMed]
- Adeel, S.; Singh, K.; Vydareny, K.H.; Kumari, M.; Shah, E.; Weitzmann, M.N.; Tangpricha, V. Bone loss in surgically ovariectomized premenopausal women is associated with T lymphocyte activation and thymic hypertrophy. J. Investig. Med. 2013, 61, 1178–1183. [Google Scholar] [CrossRef]
- Trkola, A.; Dragic, T.; Arthos, J.; Binley, J.M.; Olson, W.C.; Allaway, G.P.; Cheng-Mayer, C.; Robinson, J.; Maddon, P.J.; Moore, J.P. CD4-dependent, antibody-sensitive interactions between HIV-1 and its co-receptor CCR-5. Nature 1996, 14, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Endres, M.J.; Clapham, P.R.; Marsh, M.; Ahuja, M.; Turner, J.D.; McKnight, A.; Thomas, J.F.; Stoebenau-Haggarty, B.; Choe, S.; Vance, P.J.; et al. CD4-Independent Infection by HIV-2 Is Mediated by Fusin/CXCR4. Cell 1996, 87, 745–756. [Google Scholar] [CrossRef] [Green Version]
- Bagby, G.J.; Zhang, P.; Purcell, J.E.; Didier, P.J.; Nelson, S. Chronic Binge Ethanol Consumption Accelerates Progression of Simian Immunodeficiency Virus Disease. Alcohol. Clin. Exp. Res. 2006, 30, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Katz, P.S.; Siggins, R.W.; Porretta, C.; Armstrong, M.L.; Zea, A.H.; Mercante, D.E.; Parsons, C.; Veazey, R.S.; Bagby, G.J.; Nelson, S.; et al. Chronic alcohol increases CD8+ T-cell immunosenescence in simian immunodeficiency virus-infected rhesus macaques. Alcohol 2015, 49, 759–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poonia, B.; Nelson, S.; Bagby, G.J.; Veazey, R.S. Intestinal Lymphocyte Subsets and Turnover Are Affected by Chronic Alcohol Consumption. JAIDS J. Acquir. Immune Defic. Syndr. 2006, 41, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Wang, Q.; Ghneim, K.; Wang, L.; Rampanelli, E.; Holley-Guthrie, E.; Cheng, L.; Garrido, C.; Margolis, D.M.; Eller, L.A.; et al. Multi-omics analyses reveal that HIV-1 alters CD4+ T cell immunometabolism to fuel virus replication. Nat. Immunol. 2021, 22, 423–433. [Google Scholar] [CrossRef]
- Abdallah, M.A.; Singal, A.K. Mitochondrial dysfunction and alcohol-associated liver disease: A novel pathway and therapeutic target. Signal Transduct. Target. Ther. 2020, 5, 26. [Google Scholar] [CrossRef] [Green Version]
- Singal, A.K.; Jampana, S.C.; Weinman, S.A. Antioxidants as therapeutic agents for liver disease. Liver Int. 2011, 31, 1432–1448. [Google Scholar] [CrossRef] [Green Version]
- Mansouri, A.; Gaou, I.; De Kerguenec, C.; Amsellem, S.; Haouzi, D.; Berson, A.; Moreau, A.; Feldmann, G.; Lettéron, P.; Pessayre, D.; et al. An alcoholic binge causes massive degradation of hepatic mitochondrial DNA in mice. Gastroenterology 1999, 117, 181–190. [Google Scholar] [CrossRef]
- Duplanty, A.A.; Simon, L.; Molina, P.E. Chronic Binge Alcohol-Induced Dysregulation of Mitochondrial-Related Genes in Skeletal Muscle of Simian Immunodeficiency Virus-Infected Rhesus Macaques at End-Stage Disease. Alcohol Alcohol. 2017, 52, 298–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadikot, R.T.; Bedi, B.; Li, J.; Yeligar, S.M. Alcohol-induced mitochondrial DNA damage promotes injurious crosstalk between alveolar epithelial cells and alveolar macrophages. Alcohol 2018, 80, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Hoek, J.B.; Cahill, A.; Pastorino, J.G. Alcohol and mitochondria: A dysfunctional relationship. Gastroenterology 2002, 122, 2049–2063. [Google Scholar] [CrossRef] [Green Version]
- Levitt, D.E.; Ferguson, T.F.; Primeaux, S.D.; Zavala, J.A.; Ahmed, J.; Marshall, R.H.; Simon, L.; Molina, P.E. Skeletal muscle bioenergetic health and function in people living with HIV: Association with glucose tolerance and alcohol use. Am. J. Physiol. Integr. Comp. Physiol. 2021, 321, R781–R790. [Google Scholar] [CrossRef] [PubMed]
- Poret, J.M.; Guidry, J.J.; Simon, L.; Molina, P.E. Chronic binge alcohol and ovariectomy dysregulate omental adipose tissue metaboproteome in simian immunodeficiency virus-infected female macaques. Physiol. Genom. 2021, 53, 358–371. [Google Scholar] [CrossRef]
- Molina, P.E.; Amedee, A.M.; Veazey, R.; Dufour, J.; Volaufova, J.; Bagby, G.J.; Nelson, S. Chronic binge alcohol consumption does not diminish effectiveness of continuous antiretroviral suppression of viral load in simian immunodeficiency virus-infected macaques. Alcohol. Clin. Exp. Res. 2014, 38, 2335–2344. [Google Scholar] [CrossRef] [Green Version]
- Robichaux, S.; Lacour, N.; Bagby, G.J.; Amedee, A.M. Validation of RPS13 as a reference gene for absolute quantification of SIV RNA in tissue of rhesus macaques. J. Virol. Methods 2016, 236, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Gulzar, N. CD8+ T-Cells: Function and Response to HIV Infection. Curr. HIV Res. 2004, 2, 23–37. [Google Scholar] [CrossRef]
- Abreu, C.M.; Veenhuis, R.T.; Avalos, C.R.; Graham, S.; Parrilla, D.R.; Ferreira, E.A.; Queen, S.E.; Shirk, E.N.; Bullock, B.T.; Li, M.; et al. Myeloid and CD4 T Cells Comprise the Latent Reservoir in Antiretroviral Therapy-Suppressed SIVmac251-Infected Macaques. mBio 2019, 10, e01659-19. [Google Scholar] [CrossRef] [Green Version]
- Igarashi, T.; Imamichi, H.; Brown, C.R.; Hirsch, V.M.; Martin, M.A. The emergence and characterization of macrophage-tropic SIV/HIV chimeric viruses (SHIVs) present in CD4+T cell-depleted rhesus monkeys. J. Leukoc. Biol. 2003, 74, 772–780. [Google Scholar] [CrossRef]
- Trifunovic, A.; Larsson, N.-G. Mitochondrial dysfunction as a cause of ageing. J. Intern. Med. 2008, 263, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Edinger, A.L.; Amedee, A.; Miller, K.; Doranz, B.J.; Endres, M.; Sharron, M.; Samson, M.; Lu, Z.-H.; Clements, J.E.; Murphey-Corb, M.; et al. Differential utilization of CCR5 by macrophage and T cell tropic simian immunodeficiency virus strains. Proc. Natl. Acad. Sci. USA 1997, 94, 4005–4010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, D.N.; Aldrovandi, G.M.; Kutsch, O.; Shaw, G.M. Dynamics of HIV-1 recombination in its natural target cells. Proc. Natl. Acad. Sci. USA 2004, 12, 4204–4209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loisel-Meyer, S.; Swainson, L.; Craveiro, M.; Oburoglu, L.; Mongellaz, C.; Costa, C.; Martinez, M.; Cosset, F.-L.; Battini, J.-L.; Herzenberg, L.A.; et al. Glut1-mediated glucose transport regulates HIV infection. Proc. Natl. Acad. Sci. USA 2012, 109, 2549–2554. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Tang, H. HIV-1 Infection and Glucose Metabolism Reprogramming of T Cells: Another Approach Toward Functional Cure and Reservoir Eradication. Front. Immunol. 2020, 11, 572677. [Google Scholar] [CrossRef]
- Cretenet, G.; Clerc, I.; Matias, M.; Loisel, S.; Craveiro, M.; Oburoglu, L.; Kinet, S.; Mongellaz, C.; Dardalhon, V.; Taylor, N. Cell surface Glut1 levels distinguish human CD4 and CD8 T lymphocyte subsets with distinct effector functions. Sci. Rep. 2016, 6, 24129. [Google Scholar] [CrossRef]
- Bárcena, C.; Mayoral, P.; Quirós, P.M. Mitohormesis, an Antiaging Paradigm. Int. Rev. Cell Mol. Biol. 2018, 340, 35–77. [Google Scholar] [CrossRef]
- Ivanov, A.V.; Valuev-Elliston, V.T.; Ivanova, O.N.; Kochetkov, S.N.; Starodubova, E.S.; Bartosch, B.; Isaguliants, M.G. Oxidative Stress during HIV Infection: Mechanisms and Consequences. Oxid. Med. Cell Longev. 2016, 2016, 8910396. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Targets |
|---|---|
| Mitochondrial-Related Genes and Primer Sequences | |
| MFN2—Mitofusin 2 | Forward: CTGTGCTGGTGGATGATTAC Reverse: CCCAGTCCTTCCTCTATGT |
| SOD2—Superoxide Dismutase 2 | Forward: GACAAACCTCAGCCCTAATG Reverse: CCGTCAGCTTCTCCTTAAAC |
| GABP1—GA-Binding Protein Subunit Beta-1 | Forward: GTGCAATCTGCTACACCTAC Reverse: GATAGAAGCTCACCTGGGA |
| House Keeping Gene | |
| RPS13—Ribosomal Protein S13 | Forward: TTGCTCATCTTCCTGA Reverse: CCGGCTCTCTATCAGAATCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McTernan, P.M.; Siggins, R.W.; Catinis, A.; Amedee, A.M.; Simon, L.; Molina, P.E. Chronic Binge Alcohol and Ovarian Hormone Loss Dysregulate Circulating Immune Cell SIV Co-Receptor Expression and Mitochondrial Homeostasis in SIV-Infected Rhesus Macaques. Biomolecules 2022, 12, 946. https://doi.org/10.3390/biom12070946
McTernan PM, Siggins RW, Catinis A, Amedee AM, Simon L, Molina PE. Chronic Binge Alcohol and Ovarian Hormone Loss Dysregulate Circulating Immune Cell SIV Co-Receptor Expression and Mitochondrial Homeostasis in SIV-Infected Rhesus Macaques. Biomolecules. 2022; 12(7):946. https://doi.org/10.3390/biom12070946
Chicago/Turabian StyleMcTernan, Patrick M., Robert W. Siggins, Anna Catinis, Angela M. Amedee, Liz Simon, and Patricia E. Molina. 2022. "Chronic Binge Alcohol and Ovarian Hormone Loss Dysregulate Circulating Immune Cell SIV Co-Receptor Expression and Mitochondrial Homeostasis in SIV-Infected Rhesus Macaques" Biomolecules 12, no. 7: 946. https://doi.org/10.3390/biom12070946