

Abscisic Acid and Cytokinins Are Not Involved in the Regulation of Stomatal Conductance of Scots Pine Saplings during Post-Drought Recovery

 , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- -
- analysis of changes in ABA and CK contents in pine plants during water stress and recovery (Experiment 1);
- -
- analysis of the effects of exogenous CK treatment on stomatal conductance in pine plants (Experiment 2).
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Design
2.2.1. Experiment 1
2.2.2. Experiment 2
- “Drought”—plants under continuous drought, which were not rewatered at Day 0 and thereafter.
- “Recovery”—plants rewatered to full soil capacity and sprayed with water (12.5 mL per plant) supplemented with 0.02% Silwet™ 408 as the surfactant.
- “Recovery + 0.1 BAP”—plants rewatered to full soil capacity and sprayed with 0.1 µM 6-benzylaminopurine (6-BAP) in water (12.5 mL per plant) + 0.02% Silwet™ 408.
- “Recovery + 1.0 BAP”—plants rewatered to full soil capacity and sprayed with 1.0 µM BAP in water (12.5 mL per plant) + 0.02% Silwet™ 408.
- “Recovery + 10 BAP”—plants rewatered to full soil capacity and sprayed with 10 µM BAP (12.5 mL per plant) + 0.02% Silwet™ 408.
- “Recovery + 100 BAP”—plants rewatered to full soil capacity and sprayed with 100 µM BAP (12.5 mL per plant) +0.02% Silwet™ 408.
2.3. Stomatal Conductance to Water Vapour
2.4. Leaf Water Potential (Ψleaf)
2.5. Absolute and Relative Needle Water Content
2.6. Analysis of Plant Hormones and Related Compounds
2.7. Statistical Analysis
3. Results
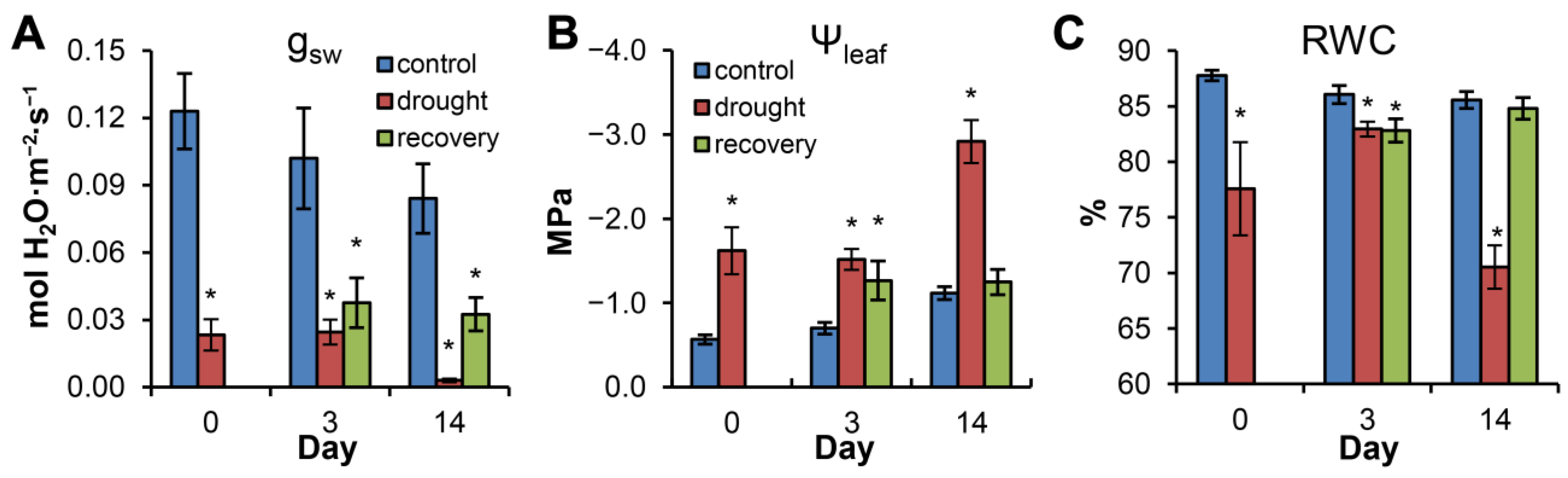
3.1. Changes in Needle Water Status in Water-Stressed and Recovering Plants (Experiment 1)
3.2. Changes in Hormonal Balance in Water-Stressed and Recovering Plants (Experiment 1)
3.3. Effects of Exogenous Cytokinin on Recovery of Needle Water Status (Experiment 2)
4. Discussion
4.1. ABA Accumulates during Water Stress and Decreases Rapidly in Recovering Plants
4.2. Dynamics of Cytokinins and CK/ABA Ratio Do Not Coincide with Recovery of Stomatal Conductance
4.3. Possible Reasons for Delayed Recovery of Stomatal Conductance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Beguería, S.; Trigo, R.; López-Moreno, J.I.; Azorín-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J. Response of Vegetation to Drought Time-Scales across Global Land Biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruehr, N.K.; Grote, R.; Mayr, S.; Arneth, A. Beyond the Extreme: Recovery of Carbon and Water Relations in Woody Plants Following Heat and Drought Stress. Tree Physiol. 2019, 39, 1285–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannenberg, S.A.; Schwalm, C.R.; Anderegg, W.R. Ghosts of the Past: How Drought Legacy Effects Shape Forest Functioning and Carbon Cycling. Ecol. Lett. 2020, 23, 891–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderegg, W.R.; Schwalm, C.; Biondi, F.; Camarero, J.J.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Shevliakova, E.; Williams, A.P. Pervasive Drought Legacies in Forest Ecosystems and Their Implications for Carbon Cycle Models. Science 2015, 349, 528–532. [Google Scholar] [CrossRef] [Green Version]
- Anderegg, W.R.; Trugman, A.T.; Badgley, G.; Konings, A.G.; Shaw, J. Divergent Forest Sensitivity to Repeated Extreme Droughts. Nat. Clim. Chang. 2020, 10, 1091–1095. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Sánchez-Salguero, R.; Vicente-Serrano, S.M.; Serra-Maluquer, X.; Gutiérrez, E.; de Luis, M.; Sangüesa-Barreda, G.; Novak, K.; Rozas, V. Drought Legacies Are Short, Prevail in Dry Conifer Forests and Depend on Growth Variability. J. Ecol. 2020, 108, 2473–2484. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Sánchez-Salguero, R.; Zavala, M.A.; Serra-Maluquer, X.; Gutiérrez, E.; De Luis, M.; Sangüesa-Barreda, G.; Novak, K.; Rozas, V. Tree Growth Response to Drought Partially Explains Regional-Scale Growth and Mortality Patterns in Iberian Forests. Ecol. Appl. 2022, 32, e2589. [Google Scholar] [CrossRef]
- DeSoto, L.; Cailleret, M.; Sterck, F.; Jansen, S.; Kramer, K.; Robert, E.M.; Aakala, T.; Amoroso, M.M.; Bigler, C.; Camarero, J.J. Low Growth Resilience to Drought Is Related to Future Mortality Risk in Trees. Nat. Commun. 2020, 11, 545. [Google Scholar] [CrossRef] [Green Version]
- Jiang, P.; Liu, H.; Piao, S.; Ciais, P.; Wu, X.; Yin, Y.; Wang, H. Enhanced Growth after Extreme Wetness Compensates for Post-Drought Carbon Loss in Dry Forests. Nat. Commun. 2019, 10, 195. [Google Scholar] [CrossRef] [Green Version]
- Fatichi, S.; Pappas, C.; Ivanov, V.Y. Modeling Plant–Water Interactions: An Ecohydrological Overview from the Cell to the Global Scale. Wiley Interdiscip. Rev. Water 2016, 3, 327–368. [Google Scholar] [CrossRef]
- Chmura, D.J.; Guzicka, M.; McCulloh, K.A.; Żytkowiak, R. Limited Variation Found among Norway Spruce Half-Sib Families in Physiological Response to Drought and Resistance to Embolism. Tree Physiol. 2016, 36, 252–266. [Google Scholar] [CrossRef] [Green Version]
- Rehschuh, R.; Cecilia, A.; Zuber, M.; Faragó, T.; Baumbach, T.; Hartmann, H.; Jansen, S.; Mayr, S.; Ruehr, N. Drought-Induced Xylem Embolism Limits the Recovery of Leaf Gas Exchange in Scots Pine. Plant Physiol. 2020, 184, 852–864. [Google Scholar] [CrossRef]
- Hájíčková, M.; Plichta, R.; Urban, J.; Volařík, D.; Gebauer, R. Low Resistance but High Resilience to Drought of Flushing Norway Spruce Seedlings. Tree Physiol. 2021, 41, 1848–1860. [Google Scholar] [CrossRef]
- Zlobin, I.E.; Kartashov, A.V.; Ivanov, Y.V.; Ivanova, A.I.; Kuznetsov, V.V. Stem Notching Decreases Stem Hydraulic Conductance but Does Not Influence Drought Impacts and Post-Drought Recovery in Scots Pine and Norway Spruce. Physiol. Plant. 2022, 174, e13813. [Google Scholar] [CrossRef]
- Han, Y.; Deng, J.; Zhou, W.; Wang, Q.-W.; Yu, D. Seasonal Responses of Hydraulic Function and Carbon Dynamics in Spruce Seedlings to Continuous Drought. Front. Plant Sci. 2022, 13, 868108. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Cochard, H. Hydraulic Failure Defines the Recovery and Point of Death in Water-Stressed Conifers. Plant Physiol. 2009, 149, 575–584. [Google Scholar] [CrossRef] [Green Version]
- Brodribb, T.J.; McAdam, S.A.; Jordan, G.J.; Martins, S.C. Conifer Species Adapt to Low-Rainfall Climates by Following One of Two Divergent Pathways. Proc. Natl. Acad. Sci. USA 2014, 111, 14489–14493. [Google Scholar] [CrossRef] [Green Version]
- Brodribb, T.J.; McAdam, S.A. Abscisic Acid Mediates a Divergence in the Drought Response of Two Conifers. Plant Physiol. 2013, 162, 1370–1377. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.M.; Gong, L.; Nie, Z.-F.; Li, F.-P.; Ahammed, G.J.; Fang, X.-W. ABA-Induced Stomatal Movements in Vascular Plants during Dehydration and Rehydration. Environ. Exp. Bot. 2021, 186, 104436. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sano, T.; Tamaoki, M.; Nakajima, N.; Kondo, N.; Hasezawa, S. Cytokinin and Auxin Inhibit Abscisic Acid-Induced Stomatal Closure by Enhancing Ethylene Production in Arabidopsis. J. Exp. Bot. 2006, 57, 2259–2266. [Google Scholar] [CrossRef] [Green Version]
- Verslues, P.E. ABA and Cytokinins: Challenge and Opportunity for Plant Stress Research. Plant Mol. Biol. 2016, 91, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Skalák, J.; Černỳ, M.; Jedelskỳ, P.; Dobrá, J.; Ge, E.; Novák, J.; Hronková, M.; Dobrev, P.; Vanková, R.; Brzobohatỳ, B. Stimulation of Ipt Overexpression as a Tool to Elucidate the Role of Cytokinins in High Temperature Responses of Arabidopsis Thaliana. J. Exp. Bot. 2016, 67, 2861–2873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prerostova, S.; Dobrev, P.I.; Gaudinova, A.; Knirsch, V.; Körber, N.; Pieruschka, R.; Fiorani, F.; Brzobohatỳ, B.; Černỳ, M.; Spichal, L. Cytokinins: Their Impact on Molecular and Growth Responses to Drought Stress and Recovery in Arabidopsis. Front. Plant Sci. 2018, 9, 655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farber, M.; Attia, Z.; Weiss, D. Cytokinin Activity Increases Stomatal Density and Transpiration Rate in Tomato. J. Exp. Bot. 2016, 67, 6351–6362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daszkowska-Golec, A.; Szarejko, I. Open or Close the Gate–Stomata Action under the Control of Phytohormones in Drought Stress Conditions. Front. Plant Sci. 2013, 4, 138. [Google Scholar] [CrossRef] [Green Version]
- Zlobin, I.E.; Ivanov, Y.V.; Kartashov, A.V.; Kuznetsov, V.V. Impact of Drought Stress Induced by Polyethylene Glycol on Growth, Water Relations and Cell Viability of Norway Spruce Seedlings. Environ. Sci. Pollut. Res. 2018, 25, 8951–8962. [Google Scholar] [CrossRef]
- Fox, H.; Doron-Faigenboim, A.; Kelly, G.; Bourstein, R.; Attia, Z.; Zhou, J.; Moshe, Y.; Moshelion, M.; David-Schwartz, R. Transcriptome Analysis of Pinus Halepensis under Drought Stress and during Recovery. Tree Physiol. 2018, 38, 423–441. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.-L.; Yin, X.; Xiong, C.-F.; Cai, B.-D.; Wu, Y.; Zhang, X.-Y.; Wei, Z.; Ye, T.; Feng, Y.-Q. Neophaseic Acid Catabolism in the 9′-Hydroxylation Pathway of Abscisic Acid in Arabidopsis Thaliana. Plant Commun. 2022, 3, 100340. [Google Scholar] [CrossRef]
- Abrams, S.R.; Loewen, M.C. Chemistry and chemical biology of ABA. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2019; Volume 92, pp. 315–339. [Google Scholar]
- Seo, M.; Marion-Poll, A. Abscisic acid metabolism and transport. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2019; Volume 92, pp. 1–49. [Google Scholar]
- Jadhav, A.S.; Taylor, D.C.; Giblin, M.; Ferrie, A.M.; Ambrose, S.J.; Ross, A.R.; Nelson, K.M.; Zaharia, L.I.; Sharma, N.; Anderson, M. Hormonal Regulation of Oil Accumulation in Brassica Seeds: Metabolism and Biological Activity of ABA, 7′-, 8′-and 9′-Hydroxy ABA in Microspore Derived Embryos of B. Napus. Phytochemistry 2008, 69, 2678–2688. [Google Scholar] [CrossRef]
- Kepka, M.; Benson, C.L.; Gonugunta, V.K.; Nelson, K.M.; Christmann, A.; Grill, E.; Abrams, S.R. Action of Natural Abscisic Acid Precursors and Catabolites on Abscisic Acid Receptor Complexes. Plant Physiol. 2011, 157, 2108–2119. [Google Scholar] [CrossRef] [Green Version]
- Pashkovskiy, P.P.; Vankova, R.; Zlobin, I.E.; Dobrev, P.; Kartashov, A.V.; Ivanova, A.I.; Ivanov, V.P.; Marchenko, S.I.; Nartov, D.I.; Ivanov, Y.V. Hormonal Responses to Short-Term and Long-Term Water Deficit in Native Scots Pine and Norway Spruce Trees. Environ. Exp. Bot. 2022, 195, 104789. [Google Scholar] [CrossRef]
- Pashkovskiy, P.P.; Vankova, R.; Zlobin, I.E.; Dobrev, P.; Ivanov, Y.V.; Kartashov, A.V.; Kuznetsov, V.V. Comparative Analysis of Abscisic Acid Levels and Expression of Abscisic Acid-Related Genes in Scots Pine and Norway Spruce Seedlings under Water Deficit. Plant Physiol. Biochem. 2019, 140, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Lomin, S.N.; Savelieva, E.M.; Arkhipov, D.V.; Pashkovskiy, P.P.; Myakushina, Y.A.; Heyl, A.; Romanov, G.A. Cytokinin Perception in Ancient Plants beyond Angiospermae. Int. J. Mol. Sci. 2021, 22, 13077. [Google Scholar] [CrossRef] [PubMed]
- Martins, S.C.; McAdam, S.A.; Deans, R.M.; DaMatta, F.M.; Brodribb, T.J. Stomatal Dynamics Are Limited by Leaf Hydraulics in Ferns and Conifers: Results from Simultaneous Measurements of Liquid and Vapour Fluxes in Leaves. Plant Cell Environ. 2016, 39, 694–705. [Google Scholar] [CrossRef] [Green Version]
- Scoffoni, C.; Sack, L.; Ort, D. The Causes and Consequences of Leaf Hydraulic Decline with Dehydration. J. Exp. Bot. 2017, 68, 4479–4496. [Google Scholar] [CrossRef] [Green Version]
- Scoffoni, C.; Albuquerque, C.; Brodersen, C.R.; Townes, S.V.; John, G.P.; Bartlett, M.K.; Buckley, T.N.; McElrone, A.J.; Sack, L. Outside-Xylem Vulnerability, Not Xylem Embolism, Controls Leaf Hydraulic Decline during Dehydration. Plant Physiol. 2017, 173, 1197–1210. [Google Scholar] [CrossRef] [Green Version]
- Creek, D.; Lamarque, L.J.; Torres-Ruiz, J.M.; Parise, C.; Burlett, R.; Tissue, D.T.; Delzon, S. Xylem Embolism in Leaves Does Not Occur with Open Stomata: Evidence from Direct Observations Using the Optical Visualization Technique. J. Exp. Bot. 2020, 71, 1151–1159. [Google Scholar] [CrossRef]
- Skelton, R.P.; Brodribb, T.J.; McAdam, S.A.; Mitchell, P.J. Gas Exchange Recovery Following Natural Drought Is Rapid Unless Limited by Loss of Leaf Hydraulic Conductance: Evidence from an Evergreen Woodland. New Phytol. 2017, 215, 1399–1412. [Google Scholar] [CrossRef] [Green Version]
- Brodribb, T.; Brodersen, C.R.; Carriqui, M.; Tonet, V.; Rodriguez Dominguez, C.; McAdam, S. Linking Xylem Network Failure with Leaf Tissue Death. New Phytol. 2021, 232, 68–79. [Google Scholar] [CrossRef]
- Cardoso, A.A.; Batz, T.A.; McAdam, S.A. Xylem Embolism Resistance Determines Leaf Mortality during Drought in Persea Americana. Plant Physiol. 2020, 182, 547–554. [Google Scholar] [CrossRef] [Green Version]
- John, G.P.; Henry, C.; Sack, L. Leaf Rehydration Capacity: Associations with Other Indices of Drought Tolerance and Environment. Plant Cell Environ. 2018, 41, 2638–2653. [Google Scholar] [CrossRef]
- Trueba, S.; Pan, R.; Scoffoni, C.; John, G.P.; Davis, S.D.; Sack, L. Thresholds for Leaf Damage Due to Dehydration: Declines of Hydraulic Function, Stomatal Conductance and Cellular Integrity Precede Those for Photochemistry. New Phytol. 2019, 223, 134–149. [Google Scholar] [CrossRef]
- Virlouvet, L.; Fromm, M. Physiological and Transcriptional Memory in Guard Cells during Repetitive Dehydration Stress. New Phytol. 2015, 205, 596–607. [Google Scholar] [CrossRef] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zlobin, I.E.; Vankova, R.; Dobrev, P.I.; Gaudinova, A.; Kartashov, A.V.; Ivanov, Y.V.; Ivanova, A.I.; Kuznetsov, V.V. Abscisic Acid and Cytokinins Are Not Involved in the Regulation of Stomatal Conductance of Scots Pine Saplings during Post-Drought Recovery. Biomolecules 2023, 13, 523. https://doi.org/10.3390/biom13030523
Zlobin IE, Vankova R, Dobrev PI, Gaudinova A, Kartashov AV, Ivanov YV, Ivanova AI, Kuznetsov VV. Abscisic Acid and Cytokinins Are Not Involved in the Regulation of Stomatal Conductance of Scots Pine Saplings during Post-Drought Recovery. Biomolecules. 2023; 13(3):523. https://doi.org/10.3390/biom13030523
Chicago/Turabian StyleZlobin, Ilya E., Radomira Vankova, Petre I. Dobrev, Alena Gaudinova, Alexander V. Kartashov, Yury V. Ivanov, Alexandra I. Ivanova, and Vladimir V. Kuznetsov. 2023. "Abscisic Acid and Cytokinins Are Not Involved in the Regulation of Stomatal Conductance of Scots Pine Saplings during Post-Drought Recovery" Biomolecules 13, no. 3: 523. https://doi.org/10.3390/biom13030523
APA StyleZlobin, I. E., Vankova, R., Dobrev, P. I., Gaudinova, A., Kartashov, A. V., Ivanov, Y. V., Ivanova, A. I., & Kuznetsov, V. V. (2023). Abscisic Acid and Cytokinins Are Not Involved in the Regulation of Stomatal Conductance of Scots Pine Saplings during Post-Drought Recovery. Biomolecules, 13(3), 523. https://doi.org/10.3390/biom13030523