
Sporadic Amyotrophic Lateral Sclerosis Skeletal Muscle Transcriptome Analysis: A Comprehensive Examination of Differentially Expressed Genes
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microarray Dataset
2.2. Identification and Analysis of Differentially Expressed Genes (DEGs)
2.3. Functional and Enrichment Analysis of DEG Pathways
2.4. Protein–Protein Interaction (PPI) Network Construction and Subnetwork Identification
2.5. Analysis of Hub Genes and PPI Networks
2.6. Prediction of miRNAs Targeting Hub Genes
3. Results
3.1. Identification and Analysis of Differentially Expressed Genes (DEGs)
3.2. Functional and Enrichment Analysis of DEG Pathways
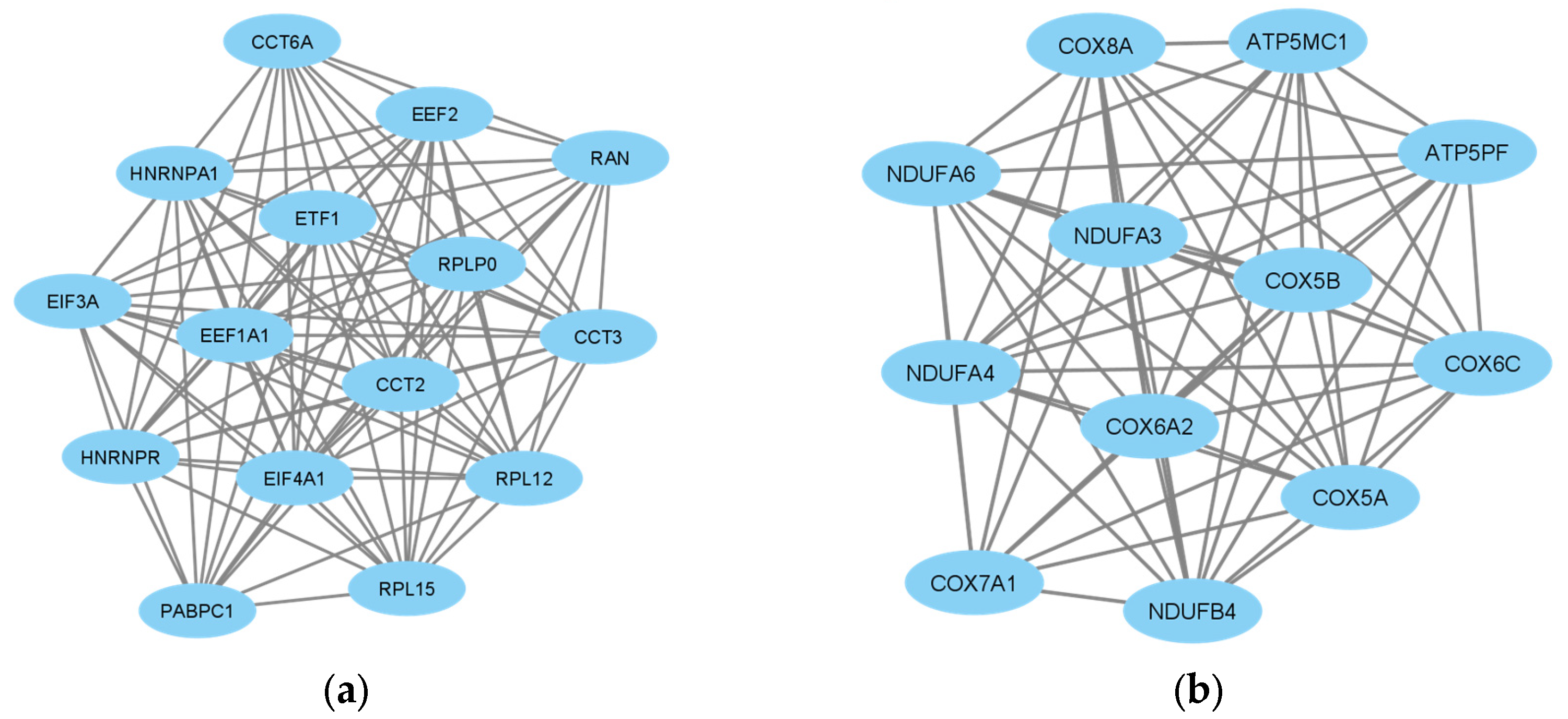
3.3. Protein–Protein Interaction (PPI) Network Construction and Subnetwork Identification
3.4. Identification of Hub Genes
3.5. Prediction of miRNAs Targeting Hub Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Longinetti, E.; Fang, F. Epidemiology of Amyotrophic Lateral Sclerosis: An Update of Recent Literature. Curr. Opin. Neurol. 2019, 32, 771–776. [Google Scholar] [CrossRef]
- Lin, J.; Huang, P.; Chen, W.; Ye, C.; Su, H.; Yao, X. Key Molecules and Pathways Underlying Sporadic Amyotrophic Lateral Sclerosis: Integrated Analysis on Gene Expression Profiles of Motor Neurons. Front. Genet. 2020, 11, 578143. [Google Scholar] [CrossRef]
- Šoltić, D.; Bowerman, M.; Stock, J.; Shorrock, H.K.; Gillingwater, T.H.; Fuller, H.R. Multi-Study Proteomic and Bioinformatic Identification of Molecular Overlap between Amyotrophic Lateral Sclerosis (Als) and Spinal Muscular Atrophy (Sma). Brain Sci. 2018, 8, 212. [Google Scholar] [CrossRef]
- Chia, R.; Chio, A.; Traynor, B.J. Novel Genes Associated with Amyotrophic Lateral Sclerosis: Diagnostic and Clinical Implications. Lancet Neurol. 2018, 17, 94–102. [Google Scholar] [CrossRef]
- Kumar, R.; Haider, S. Protein Network Analysis to Prioritize Key Genes in Amyotrophic Lateral Sclerosis. IBRO Neurosci. Rep. 2022, 12, 25–44. [Google Scholar] [CrossRef] [PubMed]
- Sever, B.; Ciftci, H.; DeMirci, H.; Sever, H.; Ocak, F.; Yulug, B.; Tateishi, H.; Tateishi, T.; Otsuka, M.; Fujita, M.; et al. Comprehensive Research on Past and Future Therapeutic Strategies Devoted to Treatment of Amyotrophic Lateral Sclerosis. Int. J. Mol. Sci. 2022, 23, 2400. [Google Scholar] [CrossRef] [PubMed]
- Butti, Z.; Patten, S.A. Rna Dysregulation in Amyotrophic Lateral Sclerosis. Front. Genet. 2018, 9, 712. [Google Scholar] [CrossRef] [PubMed]
- Cykowski, M.D.; Dickson, D.W.; Powell, S.Z.; Arumanayagam, A.S.; Rivera, A.L.; Appel, S.H. Dipeptide Repeat (Dpr) Pathology in the Skeletal Muscle of Als Patients with C9orf72 Repeat Expansion. Acta Neuropathol. 2019, 138, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Brenner, D.; Muller, K.; Wieland, T.; Weydt, P.; Bohm, S.; Lule, D.; Hubers, A.; Neuwirth, C.; Weber, M.; Borck, G.; et al. Nek1 Mutations in Familial Amyotrophic Lateral Sclerosis. Brain 2016, 139 Pt 5, e28. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Taylor, J.P. Lost in Transportation: Nucleocytoplasmic Transport Defects in Als and Other Neurodegenerative Diseases. Neuron 2017, 96, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Foerster, B.R.; Pomper, M.G.; Callaghan, B.C.; Petrou, M.; Edden, R.A.E.; Mohamed, M.A.; Welsh, R.C.; Carlos, R.C.; Barker, P.B.; Feldman, E.L. An Imbalance between Excitatory and Inhibitory Neurotransmitters in Amyotrophic Lateral Sclerosis Revealed by Use of 3-T Proton Magnetic Resonance Spectroscopy. JAMA Neurol. 2013, 70, 1009–1016. [Google Scholar] [CrossRef]
- Mitsumoto, H.; Santella, R.M.; Liu, X.; Bogdanov, M.; Zipprich, J.; Wu, H.C.; Mahata, J.; Kilty, M.; Bednarz, K.; Bell, D.; et al. Oxidative Stress Biomarkers in Sporadic Als. Amyotroph. Lateral Scler. 2008, 9, 177–183. [Google Scholar] [CrossRef] [PubMed]
- De Vos, K.J.; Chapman, A.L.; Tennant, M.E.; Manser, C.; Tudor, E.L.; Lau, K.F.; Brownlees, J.; Ackerley, S.; Shaw, P.J.; McLoughlin, D.M.; et al. Familial Amyotrophic Lateral Sclerosis-Linked Sod1 Mutants Perturb Fast Axonal Transport to Reduce Axonal Mitochondria Content. Hum. Mol. Genet. 2007, 16, 2720–2728. [Google Scholar] [CrossRef] [PubMed]
- Rosen, D.R.; Siddique, T.; Patterson, D.; Figlewicz, D.A.; Sapp, P.; Hentati, A.; Donaldson, D.; Goto, J.; O’Regan, J.P.; Deng, H.X.; et al. Mutations in Cu/Zn Superoxide Dismutase Gene Are Associated with Familial Amyotrophic Lateral Sclerosis. Nature 1993, 362, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowski, T.J., Jr.; Bosco, D.A.; Leclerc, A.L.; Tamrazian, E.; Vanderburg, C.R.; Russ, C.; Davis, A.; Gilchrist, J.; Kasarskis, E.J.; Munsat, T.; et al. Mutations in the Fus/Tls Gene on Chromosome 16 Cause Familial Amyotrophic Lateral Sclerosis. Science 2009, 323, 1205–1208. [Google Scholar] [CrossRef] [PubMed]
- DeJesus-Hernandez, M.; Mackenzie, I.R.; Boeve, B.F.; Boxer, A.L.; Baker, M.; Rutherford, N.J.; Nicholson, A.M.; Finch, N.A.; Flynn, H.; Adamson, J.; et al. Expanded Ggggcc Hexanucleotide Repeat in Noncoding Region of C9orf72 Causes Chromosome 9p-Linked Ftd and Als. Neuron 2011, 72, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Renton, A.E.; Majounie, E.; Waite, A.; Simón-Sánchez, J.; Rollinson, S.; Gibbs, J.R.; Schymick, J.C.; Laaksovirta, H.; van Swieten, J.C.; Myllykangas, L.; et al. A Hexanucleotide Repeat Expansion in C9orf72 Is the Cause of Chromosome 9p21-Linked Als-Ftd. Neuron 2011, 72, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Elden, A.C.; Kim, H.J.; Hart, M.P.; Chen-Plotkin, A.S.; Johnson, B.S.; Fang, X.; Armakola, M.; Geser, F.; Greene, R.; Lu, M.M.; et al. Ataxin-2 Intermediate-Length Polyglutamine Expansions Are Associated with Increased Risk for Als. Nature 2010, 466, 1069–1075. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, H.; Morino, H.; Ito, H.; Izumi, Y.; Kato, H.; Watanabe, Y.; Kinoshita, Y.; Kamada, M.; Nodera, H.; Suzuki, H.; et al. Mutations of Optineurin in Amyotrophic Lateral Sclerosis. Nature 2010, 465, 223–226. [Google Scholar] [CrossRef]
- Johnson, J.O.; Mandrioli, J.; Benatar, M.; Abramzon, Y.; Van Deerlin, V.M.; Trojanowski, J.Q.; Gibbs, J.R.; Brunetti, M.; Gronka, S.; Wuu, J.; et al. Exome Sequencing Reveals Vcp Mutations as a Cause of Familial Als. Neuron 2010, 68, 857–864. [Google Scholar] [CrossRef]
- Wu, C.H.; Fallini, C.; Ticozzi, N.; Keagle, P.J.; Sapp, P.C.; Piotrowska, K.; Lowe, P.; Koppers, M.; McKenna-Yasek, D.; Baron, D.M.; et al. Mutations in the Profilin 1 Gene Cause Familial Amyotrophic Lateral Sclerosis. Nature 2012, 488, 499–503. [Google Scholar] [CrossRef]
- Johnson, J.O.; Pioro, E.P.; Boehringer, A.; Chia, R.; Feit, H.; Renton, A.E.; Pliner, H.A.; Abramzon, Y.; Marangi, G.; Winborn, B.J.; et al. Mutations in the Matrin 3 Gene Cause Familial Amyotrophic Lateral Sclerosis. Nat. Neurosci. 2014, 17, 664–666. [Google Scholar] [CrossRef]
- Hirano, M.; Quinzii, C.M.; Mitsumoto, H.; Hays, A.P.; Roberts, J.K.; Richard, P.; Rowland, L.P. Senataxin Mutations and Amyotrophic Lateral Sclerosis. Amyotroph. Lateral Scler. 2011, 12, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.X.; Chen, W.; Hong, S.T.; Boycott, K.M.; Gorrie, G.H.; Siddique, N.; Yang, Y.; Fecto, F.; Shi, Y.; Zhai, H.; et al. Mutations in Ubqln2 Cause Dominant X-Linked Juvenile and Adult-Onset Als and Als/Dementia. Nature 2011, 477, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Gelfman, S.; Dugger, S.; de Araujo Martins Moreno, C.; Ren, Z.; Wolock, C.J.; Shneider, N.A.; Phatnani, H.; Cirulli, E.T.; Lasseigne, B.N.; Harris, T.; et al. A New Approach for Rare Variation Collapsing on Functional Protein Domains Implicates Specific Genic Regions in Als. Genome Res. 2019, 29, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Y.; Lin, J.L.; Feng, S.Y.; Che, C.H.; Huang, H.P.; Zou, Z.Y. Novel Variants in the Fig4 Gene Associated with Chinese Sporadic Amyotrophic Lateral Sclerosis with Slow Progression. J. Clin. Neurol. 2022, 18, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Daneshmandpour, Y.; Bahmanpour, Z.; Kazeminasab, S.; Moghadam, E.A.; Alehabib, E.; Chapi, M.; Tafakhori, A.; Aghaei, N.; Darvish, H.; Emamalizadeh, B. A Novel Mutation in the Als2 Gene in an Iranian Kurdish Family with Juvenile Amyotrophic Lateral Sclerosis. Amyotroph. Lateral Scler. Front. Degener. 2023, 24, 148–151. [Google Scholar] [CrossRef] [PubMed]
- González-Mingot, C.; Miana-Mena, F.J.; Iñarrea, P.J.; Iñiguez, C.; Capablo, J.L.; Osta, R.; Gil-Sánchez, A.; Brieva, L.; Larrodé, P. Mitochondrial Aconitase Enzymatic Activity: A Potential Long-Term Survival Biomarker in the Blood of Als Patients. J. Clin. Med. 2023, 12, 3560. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, H.M.; Dimachkie, M.M.; Agbas, A. Blood-Based Biomarkers for Amyotrophic Lateral Sclerosis. In Amyotrophic Lateral Sclerosis; Exon Publications: Brisbane, Australia, 2021. [Google Scholar]
- Zhang, P.; Feng, J.; Wu, X.; Chu, W.; Zhang, Y.; Li, P. Bioinformatics Analysis of Candidate Genes and Pathways Related to Hepatocellular Carcinoma in China: A Study Based on Public Databases. Pathol. Oncol. Res. 2021, 27, 588532. [Google Scholar] [CrossRef] [PubMed]
- Ganekal, P.; Vastrad, B.; Kavatagimath, S.; Vastrad, C.; Kotrashetti, S. Bioinformatics and Next-Generation Data Analysis for Identification of Genes and Molecular Pathways Involved in Subjects with Diabetes and Obesity. Medicina 2023, 59, 309. [Google Scholar] [CrossRef]
- Mohanan, E.M.; Jhala, D.; More, C.B.; Patel, A.K.; Joshi, C. Bioinformatics Analysis of Mirna and Its Associated Genes to Identify Potential Biomarkers of Oral Submucous Fibrosis and Oral Malignancy. Cancer Rep. 2023, 6, e1787. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Vats, A.; Ahuja, V.; Vats, K.; Khurana, S.; Vats, Y.; Gourie-Devi, M.; Wajid, S.; Ganguly, N.K.; Chakraborti, P.; et al. Functional Consequences of Familial Als-Associated Sod1(L84f) in Neuronal and Muscle Cells. FASEB J. 2024, 38, e23461. [Google Scholar] [CrossRef]
- Bernardini, C.; Censi, F.; Lattanzi, W.; Barba, M.; Calcagnini, G.; Giuliani, A.; Tasca, G.; Sabatelli, M.; Ricci, E.; Michetti, F. Mitochondrial Network Genes in the Skeletal Muscle of Amyotrophic Lateral Sclerosis Patients. PLoS ONE 2013, 8, e57739. [Google Scholar] [CrossRef] [PubMed]
- Clough, E.; Barrett, T. The Gene Expression Omnibus Database. In Statistical Genomics: Methods and Protocols; Mathé, E., Davis, S., Eds.; Springer: New York, NY, USA, 2016; pp. 93–110. [Google Scholar]
- Reimers, M.; Carey, V.J. Bioconductor: An Open Source Framework for Bioinformatics and Computational Biology. Methods Enzymol. 2006, 411, 119–134. [Google Scholar]
- Yu, G.C.; Wang, L.G.; Han, Y.Y.; He, Q.Y. Clusterprofiler: An R Package for Comparing Biological Themes among Gene Clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.A.-O.; Nastou, K.A.-O.; Mehryary, F.A.-O.; Hachilif, R.; Gable, A.L.; Fang, T.A.-O.; Doncheva, N.A.-O.; Pyysalo, S.; et al. The String Database in 2023: Protein-Protein Association Networks and Functional Enrichment Analyses for Any Sequenced Genome of Interest. Nucleic Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Bader, G.D.; Hogue, C.W.V. An Automated Method for Finding Molecular Complexes in Large Protein Interaction Networks. BMC Bioinform. 2003, 4, 2. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape Provides a Biologist-Oriented Resource for the Analysis of Systems-Level Datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. Cytohubba: Identifying Hub Objects and Sub-Networks from Complex Interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef]
- Chang, L.; Zhou, G.; Soufan, O.; Xia, J. Mirnet 2.0: Network-Based Visual Analytics for Mirna Functional Analysis and Systems Biology. Nucleic Acids Res. 2020, 48, W244–W251. [Google Scholar] [CrossRef]
- Wang, T.; Tian, X.; Kim, H.B.; Jang, Y.; Huang, Z.; Na, C.H.; Wang, J. Intracellular Energy Controls Dynamics of Stress-Induced Ribonucleoprotein Granules. Nat. Commun. 2022, 13, 5584. [Google Scholar] [CrossRef] [PubMed]
- Taminato, T.; Takeuchi, T.; Ueyama, M.; Mori, K.; Ikeda, M.; Mochizuki, H.; Nagai, Y. Therapeutic Reduction of Ggggcc Repeat Rna Levels by Hnrnpa3 Suppresses Neurodegeneration in Drosophila Models of C9orf72-Linked Als/Ftd. Hum. Mol. Genet. 2023, 32, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Naskar, A.; Nayak, A.; Salaikumaran, M.R.; Vishal, S.S.; Gopal, P.P. Phase Separation and Pathologic Transitions of Rnp Condensates in Neurons: Implications for Amyotrophic Lateral Sclerosis, Frontotemporal Dementia and Other Neurodegenerative Disorders. Front. Mol. Neurosci. 2023, 16, 1242925. [Google Scholar] [CrossRef]
- Peng, G.; Gu, A.; Niu, H.; Chen, L.; Chen, Y.; Zhou, M.; Zhang, Y.; Liu, J.; Cai, L.; Liang, D.; et al. Amyotrophic Lateral Sclerosis (Als) Linked Mutation in Ubiquilin 2 Affects Stress Granule Assembly Via Tia-1. CNS Neurosci. Ther. 2022, 28, 105–115. [Google Scholar] [CrossRef]
- Qin, S.; You, P.; Yu, H.; Su, B. Reep1 Preserves Motor Function in Sod1g93a Mice by Improving Mitochondrial Function Via Interaction with Ndufa4. Neurosci. Bull. 2023, 39, 929–946. [Google Scholar] [CrossRef]
- Aisha, Z.; Lei, J.; Zhang, Y.; Ma, J. Eef1a1 Is Involved the Regulating Neuroinflammatory Processes in Parkinson’s Disease. J. Integr. Neurosci. 2023, 22, 122. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Yan, J.; Huang, X.; Zou, C.; Chen, L.; Li, R.; Xie, J.; Pan, M.; Zou, D.; Liu, Y. Identification of Molecular Signatures Associated with Sleep Disorder and Alzheimer’s Disease. Front. Psychiatry 2022, 13, 925012. [Google Scholar] [CrossRef]
- Suzuki, M.; Tezuka, K.; Handa, T.; Sato, R.; Takeuchi, H.; Takao, M.; Tano, M.; Uchida, Y. Upregulation of Ribosome Complexes at the Blood-Brain Barrier in Alzheimer’s Disease Patients. J. Cereb. Blood Flow. Metab. 2022, 42, 2134–2150. [Google Scholar] [CrossRef]
- Wallach, T.; Mossmann, Z.J.; Szczepek, M.; Wetzel, M.; Machado, R.; Raden, M.; Miladi, M.; Kleinau, G.; Kruger, C.; Dembny, P.; et al. Microrna-100-5p and Microrna-298-5p Released from Apoptotic Cortical Neurons Are Endogenous Toll-Like Receptor 7/8 Ligands That Contribute to Neurodegeneration. Mol. Neurodegener. 2021, 16, 80. [Google Scholar] [CrossRef]
- Nogami, M.; Ishikawa, M.; Doi, A.; Sano, O.; Sone, T.; Akiyama, T.; Aoki, M.; Nakanishi, A.; Ogi, K.; Yano, M.; et al. Identification of Hub Molecules of Fus-Als by Bayesian Gene Regulatory Network Analysis of Ipsc Model: Ibrn. Neurobiol. Dis. 2021, 155, 105364. [Google Scholar] [CrossRef]
- Malacarne, C.; Galbiati, M.; Giagnorio, E.; Cavalcante, P.; Salerno, F.; Andreetta, F.; Cagnoli, C.; Taiana, M.; Nizzardo, M.; Corti, S.; et al. Dysregulation of Muscle-Specific Micrornas as Common Pathogenic Feature Associated with Muscle Atrophy in Als, Sma and Sbma: Evidence from Animal Models and Human Patients. Int. J. Mol. Sci. 2021, 22, 5673. [Google Scholar] [CrossRef]
- Ruffo, P.; Catalano, S.; La Bella, V.; Conforti, F.L. Deregulation of Plasma Microrna Expression in a Tardbp-Als Family. Biomolecules 2023, 13, 706. [Google Scholar] [CrossRef]
- Cheng, J.; Ho, W.K.; Wu, B.T.; Liu, H.P.; Lin, W.Y. Mirna Profiling as a Complementary Diagnostic Tool for Amyotrophic Lateral Sclerosis. Sci. Rep. 2023, 13, 13805. [Google Scholar] [CrossRef]
- Rasheed, M.; Asghar, R.; Firdoos, S.; Ahmad, N.; Nazir, A.; Ullah, K.M.; Li, N.; Zhuang, F.; Chen, Z.; Deng, Y. A Systematic Review of Circulatory Micrornas in Major Depressive Disorder: Potential Biomarkers for Disease Prognosis. Int. J. Mol. Sci. 2022, 23, 1294. [Google Scholar] [CrossRef]
- Keon, M.; Musrie, B.; Dinger, M.; Brennan, S.E.; Santos, J.; Saksena, N.K. Destination Amyotrophic Lateral Sclerosis. Front. Neurol. 2021, 12, 596006. [Google Scholar] [CrossRef]
- Abel, O.; Powell, J.F.; Andersen, P.M.; Al-Chalabi, A. Alsod: A User-Friendly Online Bioinformatics Tool for Amyotrophic Lateral Sclerosis Genetics. Hum. Mutat. 2012, 33, 1345–1351. [Google Scholar] [CrossRef]
- Shefner, J.M.; Musaro, A.; Ngo, S.T.; Lunetta, C.; Steyn, F.J.; Robitaille, R.; De Carvalho, M.; Rutkove, S.; Ludolph, A.C.; Dupuis, L. Skeletal Muscle in Amyotrophic Lateral Sclerosis. Brain 2023, 146, 4425–4436. [Google Scholar] [CrossRef]
- Pradat, P.F.; Barani, A.; Wanschitz, J.; Dubourg, O.; Lombes, A.; Bigot, A.; Mouly, V.; Bruneteau, G.; Salachas, F.; Lenglet, T.; et al. Abnormalities of Satellite Cells Function in Amyotrophic Lateral Sclerosis. Amyotroph. Lateral Scler. 2011, 12, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Feneberg, E.; Gordon, D.; Thompson, A.G.; Finelli, M.J.; Dafinca, R.; Candalija, A.; Charles, P.D.; Mager, I.; Wood, M.J.; Fischer, R.; et al. An Als-Linked Mutation in Tdp-43 Disrupts Normal Protein Interactions in the Motor Neuron Response to Oxidative Stress. Neurobiol. Dis. 2020, 144, 105050. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Banerjee, S.; Savran, D.; Rajes, C.; Wiese, S.; Girdhar, A.; Schwierz, N.; Lee, C.; Shorter, J.; Schmidt, M.; et al. Cryo-Em Structure of the Full-Length Hnrnpa1 Amyloid Fibril. J. Mol. Biol. 2023, 435, 168211. [Google Scholar] [CrossRef]
- Moreno-García, L.; Miana-Mena, F.J.; Moreno-Martínez, L.; de la Torre, M.; Lunetta, C.; Tarlarini, C.; Zaragoza, P.; Calvo, A.C.; Osta, R. Inflammasome in Als Skeletal Muscle: Nlrp3 as a Potential Biomarker. Int. J. Mol. Sci. 2021, 22, 2523. [Google Scholar] [CrossRef] [PubMed]
- Larson, K.C.; Martens, L.H.; Marconi, M.; Dejesus, C.; Bruhn, S.; Miller, T.A.; Tate, B.; Levenson, J.M. Preclinical Translational Platform of Neuroinflammatory Disease Biology Relevant to Neurodegenerative Disease. J. Neuroinflamm. 2024, 21, 37. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, C.; Zhan, X.; Li, B.; Zhang, Z.; Li, S.; Xie, Y.; Song, X.; Shen, Y.; Liu, J.; et al. R13 Preserves Motor Performance in Sod1(G93a) Mice by Improving Mitochondrial Function. Theranostics 2021, 11, 7294–7307. [Google Scholar] [CrossRef] [PubMed]
- Jhanji, R.; Behl, T.; Sehgal, A.; Bungau, S. Mitochondrial Dysfunction and Traffic Jams in Amyotrophic Lateral Sclerosis. Mitochondrion 2021, 58, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Toivonen, J.M.; Manzano, R.; Oliván, S.; Zaragoza, P.; García-Redondo, A.; Osta, R. Microrna-206: A Potential Circulating Biomarker Candidate for Amyotrophic Lateral Sclerosis. PLoS ONE 2014, 9, e89065. [Google Scholar] [CrossRef]
- Liu, H.; Lan, S.; Shi, X.J.; Fan, F.C.; Liu, Q.S.; Cong, L.; Cheng, Y. Systematic Review and Meta-Analysis on Micrornas in Amyotrophic Lateral Sclerosis. Brain Res. Bull. 2023, 194, 82–89. [Google Scholar] [CrossRef]
- Ju, H.Y.; Tang, S.S.; Li, B.J.; Luo, X.; Li, Q. The Expression Levels of Circulating Mir-140-3p, Mir-130a-3p, and Mir-320b as Diagnostic Biomarkers in Acute Ischemic Stroke. Kaohsiung J. Med. Sci. 2023, 39, 927–935. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | miRNA Accession | Gene Target | Tissue | Up/Down |
|---|---|---|---|---|
| hsa-let-7a-5p | MIMAT0000062 | EEF2 | Muscle | Up |
| hsa-mir-100-5p | MIMAT0000098 | EEF1A1 RPL15 RPL12 | Muscle | Up |
| EEF1A1 RPL15 RPL12 | ||||
| hsa-mir-125b-5p | MIMAT0000423 | EEF1A1 | Muscle | Up |
| RPLP0 | ||||
| PABPC1 | ||||
| EEF2 | ||||
| hsa-mir-133a-3p | MIMAT0000427 | EIF4A1 | Muscle | Up |
| hsa-mir-133b | MIMAT0000770 | EIF4A1 | Muscle | Up |
| hsa-mir-134-3p | MIMAT0026481 | RPL12 | Muscle | Up |
| hsa-mir-1-3p | MIMAT0000416 | HNRNPA1 HNRNPR EEF1A1 EIF4A1 RPL15 RPL12 | Muscle | Up |
| hsa-mir-140-3p | MIMAT0004597 | RPLP0 HNRNPA1 EIF4A1 EEF2 RPL15 | Muscle | Up |
| hsa-mir-193a-5p | MIMAT0004614 | PABPC1 | Muscle | Up |
| hsa-mir-206 | MIMAT0000462 | HNRNPA1 | Muscle | Up |
| hsa-mir-26a-5p | MIMAT0000082 | EEF2 | Muscle | Up |
| hsa-mir-27a-3p | MIMAT0000084 | PABPC1 HNRNPA1 EEF2 | Muscle | Up |
| hsa-let-7a-5p | MIMAT0000062 | NDUFB4 NDUFA3 COX8A | Muscle | Down |
| hsa-mir-100-5p | MIMAT0000098 | COX5A | Muscle | Down |
| hsa-mir-134-5p | MIMAT0000447 | COX8A | Muscle | Down |
| hsa-mir-1-3p | MIMAT0000416 | NDUFB4 COX6C NDUFA3 COX5A NDUFA6 | Muscle | Down |
| hsa-mir-26a-5p | MIMAT0000082 | COX5A COX5B COX8A | Muscle | Down |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gascón, E.; Zaragoza, P.; Calvo, A.C.; Osta, R. Sporadic Amyotrophic Lateral Sclerosis Skeletal Muscle Transcriptome Analysis: A Comprehensive Examination of Differentially Expressed Genes. Biomolecules 2024, 14, 377. https://doi.org/10.3390/biom14030377
Gascón E, Zaragoza P, Calvo AC, Osta R. Sporadic Amyotrophic Lateral Sclerosis Skeletal Muscle Transcriptome Analysis: A Comprehensive Examination of Differentially Expressed Genes. Biomolecules. 2024; 14(3):377. https://doi.org/10.3390/biom14030377
Chicago/Turabian StyleGascón, Elisa, Pilar Zaragoza, Ana Cristina Calvo, and Rosario Osta. 2024. "Sporadic Amyotrophic Lateral Sclerosis Skeletal Muscle Transcriptome Analysis: A Comprehensive Examination of Differentially Expressed Genes" Biomolecules 14, no. 3: 377. https://doi.org/10.3390/biom14030377