
Two Modulators of Skeletal Development: BMPs and Proteoglycans

Department of Anatomy, Physiology and Pharmacology (APP), College of Medicine, University of Saskatchewan, 2D01-107 Wiggins Road, Saskatoon, SK S7N 5E5, Canada
*
Author to whom correspondence should be addressed.
J. Dev. Biol. 2022, 10(2), 15; https://doi.org/10.3390/jdb10020015
Submission received: 1 February 2022
/
Revised: 21 March 2022
/
Accepted: 23 March 2022
/
Published: 6 April 2022
(This article belongs to the Special Issue Zebrafish—a Model System for Developmental Biology II)
Abstract
:During embryogenesis, skeletal development is tightly regulated by locally secreted growth factors that interact with proteoglycans (PGs) in the extracellular matrix (ECM). Bone morphogenetic proteins (BMPs) are multifunctional growth factors that play critical roles in cartilage maturation and bone formation. BMP signals are transduced from plasma membrane receptors to the nucleus through both canonical Smad and noncanonical p38 mitogen-activated protein kinase (MAPK) pathways. BMP signalling is modulated by a variety of endogenous and exogenous molecular mechanisms at different spatiotemporal levels and in both positive and negative manners. As an endogenous example, BMPs undergo extracellular regulation by PGs, which generally regulate the efficiency of ligand-receptor binding. BMP signalling can also be exogenously perturbed by a group of small molecule antagonists, such as dorsomorphin and its derivatives, that selectively bind to and inhibit the intracellular kinase domain of BMP type I receptors. In this review, we present a current understanding of BMPs and PGs functions in cartilage maturation and osteoblast differentiation, highlighting BMP–PG interactions. We also discuss the identification of highly selective small-molecule BMP receptor type I inhibitors. This review aims to shed light on the importance of BMP signalling and PGs in cartilage maturation and bone formation.
1. Introduction
Bone morphogenetic proteins (BMPs) are multifunctional growth factors with over 20 family members that play a key role in a variety of biological processes, from embryonic stage to adulthood. However, they are best known for their ability to induce bone formation [1,2,3]. BMPs are the biggest subfamily of the Transforming Growth Factor (TGF-β) superfamily, with highly conserved amino acid sequences from insects to humans [4]. According to sequence and structural homology, BMP ligands have been divided into different groups: BMP-2 and 4; BMP-5, 6, 7, 8, and 8B; BMP-9/GDF-2 (Growth Differentiation Factor-2) and BMP-10; BMP-11/GDF-11 and GDF-8; BMP-12/GDF-7, BMP-13/GDF-6, and BMP-14/GDF-5; BMP-15/GDF-9 and GDF-9b; GDF-1 and 3; and BMP-3 and BMP-3b/GDF-10. BMP family members may form homodimeric or heterodimeric proteins to signal, except for BMP-3 and BMP-15, which biologically act as monomers. However, the osteoinduction ability and signalling activity of homodimers, such as BMP-2, BMP-4, BMP-5, BMP-6, and BMP-7, increase 30- to 50-fold when they are in heteromeric forms, such as BMP2/5, BMP2/6, BMP2/7, and BMP4/7. Higher binding affinity for BMP receptors explain the increased signalling activity of BMP heterodimers [5,6,7].
BMP signalling is mediated through BMP receptors (BMPRs) (Figure 1). A total of four BMP receptors type I (BMPRI) have been identified for BMPs: activin receptor-like kinase 1 (ACVRL1 or ALK1), activin receptor IA (ACVR1 or ALK2), type 1A BMP receptor (BMPRIA or ALK3), and type 1B BMP receptor (BMPRIB or ALK6). Three BMP receptors type II (BMPRII) are known to interact with BMPs: type 2 BMP receptor (BMPRII or BMPR2), activin type 2A receptor (ActRIIA or ACVR2A), and activin type 2B receptor (ActRIIB or ACVR2B). While BMPRIA, BMPRIB, and BMPR2 are specific to BMPs, ALK2, ACVR2A, and ACVR2B are also shared with activins, a member of the TGF-β superfamily [8]. Activin A has a high affinity for the BMPRs type 2, ACVR2A and ACVR2B, which suppresses BMP-6 and BMP-9 signalling by competing for binding to ACVR2A and ACVR2B receptors in conjunction with ALK-2 [9]. BMP dimers bind to BMPRs with varying degrees of affinity [10]. Although TGF-β and activin do not interact with their type I receptors, BMPs can bind to either BMPR I or II on their own; however, their interaction with both receptors increases the ligand’s binding affinity. BMP-9 has the highest affinity for ALK1 and binds poorly to ALK2 [11]; BMP-10 preferentially binds to ALK1, over ALK3 and ALK6; BMP-5, BMP-6, and BMP-7 signal through ALK2, but BMP-6 can also bind to ALK3 and ALK6; and BMP-2 and BMP-4 connect most strongly with ALK3 and ALK6 [12,13]. Interestingly, the different binding affinities of BMPs can determine the mechanism of heterotetrameric signalling complex formation. BMP-6 and BMP-7, for example, bind type II receptors and recruit type I receptors, whereas BMP-2 and BMP-4 primarily bind type I receptors, then recruit type II receptors [14].
BMPs bind to a heterotetrameric complex composed of two BMPRI and two BMPRII transmembrane serine/threonine kinase receptors, acting through them to elicit the downstream transcriptional changes that control cartilage maturation and osteoblast differentiation. BMPs attach to type I and type II receptor complexes, activating the type II receptor, which cross-phosphorylates specific serine and threonine residues at the juxtamembrane glycine-serine-rich (GS) domain of the type I receptor. BMP signalling is transduced intracellularly by a canonical Smad pathway and a noncanonical p38 pathway. In the canonical pathway, activated BMPRs phosphorylate and activate receptor-specific Smad 1, 5, and 8, which bind and recruit Smad 4 (co-Smad) proteins to form heteromeric complexes that translocate into the nucleus. During chondrocyte maturation, BMP-activated Smad complexes control the expression of important osteogenic genes and recruit transcription factors, such as Hoxc-8, FAST-1, OAZ, Runx2, AP-1, and STAT [15,16]. Smad1/Runx2 activity generated by BMP has been demonstrated to upregulate the expression of GADD45β protein, which is present in the nucleus of late hypertrophic chondrocytes and acts as an activator of matrix metalloproteinase-13 (MMP-13) expression. GADD45β inhibition slows terminal chondrocyte differentiation [17,18]. In the noncanonical pathway, activated BMPRs phosphorylate TAK1, which recruits TAB1 and initiates a p38 MAPK signalling cascade. In this cascade, the phosphorylated form of p38 travels into the nuclei to regulate gene expression. In pre-osteoblasts, Runx2, Dlx5, and Sp7 are phosphorylated by p38 to increase their transcriptional activity (Figure 1) [19,20,21].
Diverse interaction of BMPs with molecules in the extracellular matrix (ECM) is a crucial aspect of their biology. For example, fibrillins, the main structural components of microfibrils, regulate BMP signalling pathways by sequestering them [22,23,24]. Furthermore, Drosophila collagen type IV interacts with BMP receptors through binding to Decapentaplegic (Dpp), a functional ortholog of BMP-2 and BMP-4 in vertebrates [25]. Collagen type II features a chordin-like Von Willebrand factor type C (VWC) domain that binds to BMP-2, acting as a negative regulator for this essential chondrogenic growth factor. Many ECM proteins, as well as recognised BMP regulators, have the VWC/chordin domain, which acts as a negative regulator of BMP activities [26]. Moreover, some data suggest that hyaluronan (HA) can mediate and modulate BMP-7 responses [27,28,29]. Another example of the ECM’s participation in BMP biological activities and cell signalling are proteoglycans (PGs). PGs can operate as a coordinator for BMP signalling at the cell surface, where ligands bind to signalling receptors. This review intends to include literature on BMPs, and then PGs, on their individual roles in regulating bone formation, and ends by discussing how these two regulating molecules interact together and how their interaction further modulates skeletal development.
2. Role of BMPs in Skeletal Development
Bone formation, or ossification, begins during the early stages of embryonic life. Intramembranous ossification and endochondral ossification are the two main ways of bone formation. Endochondral ossification, ultimately forming what are called chondral bones, begins when mesenchymal stem cells (MSCs) condense and differentiate into chondrocytes to produce a cartilage template [30,31,32]. A sheath of cells that encapsulates the cartilage differentiates into the perichondrium. Chondrocytes proliferate and produce many matrix molecules, such as collagen type 2 and aggrecan, the latter of which is the most abundant PG in cartilage [33,34,35,36]. Then, a group of chondrocytes, generally in the center region of the developing cartilage, passes through a developmental transition termed maturation. During chondrocyte maturation, cells undergo hypertrophy and express maturation genes, such as Indian hedgehog (Ihh) and col10a1, while changing and mineralizing their ECM. Cells in the perichondrium surrounding the hypertrophic zone differentiate into osteoblasts at the same time that hypertrophic differentiation begins. Because it ossifies and produces osteoblasts, this area of the perichondrium is called the periosteum [37]. The cartilage ECM they construct is then degraded by invading blood vessels and gradually replaced mostly by marrow, but also by some trabecular bone [38,39].
One of the key signalling systems involved in cell commitment to the chondrogenic lineage and subsequent progression within the growth plate is the BMP pathway (Figure 2). BMPs control the expression of numerous chondrocyte-specific genes and matrix production and have a role in all stages of chondrogenesis, including early patterning and MSCs condensation, chondrocyte proliferation, and hypertrophic differentiation in the growth plate [40,41]. Multiple BMPs, including BMPs 2, 4, 5, and 7, as well as GDF5, are expressed surrounding or within early condensing mesenchyme in limb buds and developing somites, implying a function for BMP pathways in the early phases of condensation [42,43,44,45,46]. In response to BMP signalling, Sox9, a significant effector of chondrogenesis, induces the production of chondrocytic genes, such as type II collagen [47,48,49]. After the development of a perichondrium, many BMPs, including BMPs 2, 4, and 5, become strongly expressed there [50,51,52,53,54,55]. BMPs 2 and 6 are produced by hypertrophic chondrocytes [52,54]. BMP7 is found in proliferating chondrocytes, especially in the vicinity of the perichondrium [44,54]. Finally, GDFs 5, 6, and 7 are overexpressed in areas where joints are formed. As a result, every area of the growth plate exhibits overlapping expression of several BMP ligands [56,57,58].
BMP signalling induces Ihh expression, and both signals cooperate to regulate cartilage maturation [59,60]. Mice missing the Ihh gene have significant skeletal defects, including decreased chondrocyte proliferation and maturation, as well as a lack of mature osteoblasts, both of which are deleterious to bone growth [61]. Moreover, Ihh released by maturing chondrocytes induces the perichondrium to ossify [37,59]. During this process, undifferentiated cells in the perichondrium differentiate to the osteoblasts, and cortical bone is formed around the cartilage template. Mouse Ihh and zebrafish ihha mutants both display delayed perichondral ossification [62,63]. Direct effects on chondrocytes and upregulation of Ihh expression are two ways that BMP promotes chondrocytes’ proliferation. According to a study that used a ChIP-based cloning approach, the promoter region of Ihh contains numerous motifs that bind to Smad 4 and are essential for BMP-dependent activation [64]. Ihh, in turn, keeps BMP levels constant, demonstrating the existence of a positive feedback loop [59,65]. This impact may be direct since Gli transcription factors, which are downstream mediators of Ihh signalling, directly upregulate BMP-4 and BMP-7 promoter activity [66].
BMPs are strong osteoblast differentiation and bone formation inducers (Figure 2) [67,68,69]. BMPs maintain bone mass after birth by promoting the differentiation of MSCs into osteoblasts and controlling their differentiation potential [70,71,72,73]. BMP-2, 4, 5, 6, and 7 are known as powerful osteogenic factors [74]. Most BMPs can successfully promote the terminal differentiation of committed osteoblast precursors and osteoblasts; however, BMP-2, 6, and 9 may be the most powerful agents for promoting the osteoblast lineage-specific differentiation of mesenchymal progenitor cells among all of the BMPs investigated [40]. BMP-2 induces or promotes the expression of Runx2 and Sp7 (Osx), which are essential transcription factors for osteoblast differentiation and bone formation, as well as osteoblast differentiation markers, such as alkaline phosphatase (ALP), type I collagen, and osteocalcin [49,75,76,77,78,79,80]. BMP-7 increases matrix mineralization and promotes ALP activity [81,82].
3. PGs’ Function in Cell-ECM Crosstalk and Skeletal Development
PGs are structural molecules in ECM that offer novel perspectives in cell-ECM crosstalk by regulating the availability of signalling molecules. PGs are sugar-coated proteins that are made up of a core protein, a tetrasaccharide linkage region, and one or more covalently connected repeating disaccharide side chains, which are called glycosaminoglycans (GAGs) [83]. Chondroitin sulfate (CS), heparan sulfate (HS), keratan sulfate (KS), dermatan sulfate (DS), and heparin (HP) are different types of GAGs in PGs’ structure [84]. Together with HA and link proteins, PGs have a tendency to assemble into massive supramolecular complexes > 200 MDa [85]. PGs were thought to be passive, structural molecules, but recent studies have drastically changed that perception by revealing nonstructural, biological roles for PGs through binding and release of numerous signalling molecules and modulating the activity and bioavailability of growth factors and morphogens [86,87].
Although chondrocytes create several minor PGs, aggrecan is the most abundant PG expressed during endochondral bone development [88]. Versican, a large CSPG, is expressed in the early limb bud’s undifferentiated mesenchymal cells and during the beginning of prechondrogenic condensation, before disappearing with chondrocyte differentiation [89]. Concurrently with versican downregulation, a substantial aggrecan expression occurs during the development and maturation of the chondrocyte [90]. The large CSPG aggrecan is the most common PG found in cartilage. Aggrecan includes 100 CS and 25–30 KS chains in adult articular cartilage [91]. The expression pattern of aggrecan mRNA across the growth plate of normal limbs shows that various phases of chondrocyte differentiation need differing amounts of aggrecan in the ECM. The findings that pre-hypertrophic chondrocytes in the wild-type growth plate produce the greatest amounts of aggrecan is consistent with the evidence that the loss of aggrecan across the growth plate impacts chondrocytes during the transition from the pre-hypertrophic to the hypertrophic state [92].
Aggrecan plays a vital role for endochondral bone development and the function of permanent cartilage structures in both human and animal species. Human patients with Kashin–Beck disease, an endemic osteochondropathy found in regions of China, are characterised by low levels of aggrecan, short stature and abnormalities of the limbs and fingers, deformed growth plates, and chondrocyte apoptosis [93,94]. In addition, mutations in the aggrecan coding genes, such as ACAN or AGC1, are responsible for Spondyloepimetaphyseal dysplasia (SEMD), osteochondritis dissecans with early and severe onset of osteoarthritis in humans, and a variety of short-stature syndromes with rapid bone maturation. AGC1 mutations induce human aggrecanopathies by causing haploinsufficiency or cartilage structural disturbance [95,96]. Studies on mice have shown that a single 7-bp deletion in exon 5 of the Agc1 gene results in a premature stop codon in exon 6 and formation of shortened aggrecan molecules which causes cartilage matrix deficit (cmd). Homozygous cmd mice (cmd/cmd) exhibited phenotypes including dwarf-like characteristics, chondrodysplasia, aberrant collagen fibrillogenesis, and a cleft palate, which demonstrate aggrecan’s critical role in cartilage maturation. Furthermore, homozygous cmd mouse’s articular cartilage was found to have tightly packed chondrocytes surrounded by a little matrix, while the growth plate cartilage had chondrocytes arranged in disorganised columns of diminished length in severely diminished proliferative, pre-hypertrophic zones, consistent with the mouse’s lower proportions [97,98,99,100]. In other studies, in chicks, due to a premature stop codon, chondrocytes produce a truncated aggrecan core protein precursor that is not translocated to the Golgi apparatus for processing, resulting in the absence of aggrecan in cartilage, chondrodysplasia, disrupted organisation of the hyaline and growth plate cartilages, and severe skeletal stature reduction [26,92,101,102,103,104].
During endochondral ossification, aggrecan interacts with growth factors and morphogens to control chondrocyte proliferation and differentiation. It was shown that an aggrecan-rich matrix and the correct sulfation of aggrecan’s CS chains are required for the proper interaction with growth factors to establish a morphogen gradient, which modulates the coordination of numerous different signalling pathways during growth plate morphogenesis. Aggrecan is required for creating an appropriate Ihh gradient; these results are supported by the findings that Ihh binds CS chains in vitro and that the Ihh gradient is reduced in the undersulfated CSPG matrix of the brachymorphic mouse growth plate. Furthermore, wingless-related proteins (Wnts) and fibroblast growth factors (FGFs) are among the cell signalling pathways that can be influenced by CS [86,105,106,107]. In another study, a severe chondrodysplasia characterized by the substantial upregulation of TGF-β signalling was revealed in a gene trap mutation in the chondroitin-4-sulfotransferase 1 (C4st1) gene, which causes downregulation of 4-O-sulfated chondroitin production [108]. The lack of aggrecan is accompanied by the deregulation of multiple genes previously implicated in hypertrophic chondrocyte development and osteoblast differentiation. The absence of aggrecan in the matrix may also interfere with growth factor availability, which is required for communication between the perichondrium and growing chondrocytes [92]. The early and enhanced invasion of the growth plate hypertrophic zone by blood vessels and osteoblasts may result from an aggrecan-induced matrix deficit; indeed, an anti-angiogenic role for the aggrecan matrix has been suggested [109]. Angiogenic factors, basic fibroblast growth factor (bFGF) and Vascular endothelial growth factor (VEGF), produced by growth plate chondrocytes, induce endothelial cells to move towards hypertrophic cells [39,110]. However, in the lack of aggrecan, a recognised diffusion barrier to several factors in cartilage, this mechanism may be changed (Figure 3) [111].
Perlecan, also known as HSPG-2 (heparan sulfate proteoglycan 2), is a multifunctional, modular PG that promotes chondrocyte proliferation, differentiation, and matrix synthesis by interacting with a wide range of ligands, such as growth factors, morphogens, and ECM-stabilizing glycoproteins. HS, one of the GAGs found in perlecan, is an essential extracellular component. The release of HS-bound cytokines, growth factors, morphogens, proteases, and inhibitory proteins induces matrix remodelling, which regulates numerous cellular pathological and physiological processes. Perlecan possesses chondrogenic capabilities and can influence cell signalling, matrix assembly, and new tissue development via its HS chains. Perlecan’s HS side chains can bind and store growth factors, including FGF-1, 2, 4, and 9, and operate as low-affinity co-receptors, emphasizing the protein’s relevance in growth and development. FGF-7 has also been found to attach to domain III of the perlecan core protein, which appears to control the growth factor’s activity [112]. Perlecan binds, stores, and sequesters FGF-1 in the cartilage matrix, preventing proper Fgfr3 signalling activity and influencing normal cartilage and bone growth. Perlecan’s presence in the pericellular area may help this process. Other HS-containing cell surface receptors may be involved in Fgfr3 signalling, collaborating with perlecan in proper cartilage formation. Moreover, FGF-2 can bind to perlecan and act as a mechanotransducer in chondrocytes. In human articular chondrocytes, FGF-2 can also upregulate the transcription of matrix metallopeptidases 1 and 13 (MMP1 and MMP13), two enzymes that play significant roles in cartilage degradation. These findings suggest that perlecan plays various roles throughout embryonic development, emphasising the relevance of matrix structure in cellular activities [113,114,115].
In addition to regulating cartilage maturation, PGs have been identified as modulators of osteoblast differentiation [116]. It was shown that CS chains enhance osteoblast differentiation by binding to both cadherin-1 and n-cadherin, decreasing extracellular signal-regulated kinase 1/2 (ERK1/2) phosphorylation, activating Smad 3 and Smad 1/5/8 signalling pathways, and increasing the expression of osteoblastic differentiation markers, such as ALP [117]. Another regulator of osteoblast differentiation are syndecans, which are cell-surface HSPGs that can act as low-affinity co-receptors to help ligands dock and concentrate. Syndecans also influence intracellular signalling by interacting with high-affinity receptors and integrins [118,119]. Out of the four syndecans, Syndecan-2, was found to be particularly associated with osteoblast differentiation during mouse development and in adult bone. Syndecan-2 is also expressed in the periosteum at the start of endochondral ossification, and its expression rises as osteoblast differentiation progresses [120]. BMP-2 and Runx2 are osteogenic mediators that tightly upregulate Syndecan-2 in osteoblast. As a result, the amount of syndecan-2 in osteoblasts appears to be strictly regulated [121,122]. Syndecan-2 overexpression enriches the bone surface with HS and results in an increased bone mass due to a potent inhibition of resorption. Furthermore, it leads to increased bone marrow cell death and lower populations of osteoblast and osteoclast precursors. Multiple pathways, including phosphoinositide 3-kinase (PI3K), MAPK, nuclear factor kappa-B (NF-κB), and protein kinase C, as well as canonical and noncanonical Wnt pathways, are altered by the overexpression of syndecan-2 in osteosarcoma cells. Moreover, it alters the osteoblast environment by downregulating the Wnt/β-catenin/T-cell factor (TCF) pathway [123,124,125]. This family’s syndecan-3 is expressed during limb cartilage development. Syndecan-3 is expressed transiently during the pre-cartilage condensation of the skeletal components of the limb and afterwards in differentiating periosteum osteoblasts. Anti-syndecan-3 antibodies have been demonstrated to decrease limb cartilage development in vitro [126].
Small leucine-rich PGs (SLRPs) are involved in all phases of bone formation, including osteogenesis, mineral deposition, and bone remodeling, by interacting with cell surface receptors and growth factors. Skeletal growth, craniofacial structure, dentin production, and collagen fibrillogenesis are all affected by SLRPs. Biglycan and decorin are both class I subtypes of the SLRPs with CS/DS side chains [127]. Biglycan can regulate the activity of multifunctional growth factors, such as TGF-β, BMP4, and Wnt, which all play a role in the osteogenic program [128,129,130]. Biglycan has been shown to activate the Wnt pathway by binding to Wnt3a, the canonical Wnt ligand, and Wnt receptor low-density lipoprotein receptor-related protein 6 (LRP6). Both glycosylated and non-glycosylated forms of biglycan stimulate this signalling pathway. Biglycan is also implicated in ERK phosphorylation and signal transmission via the transcription factor Runx2. The activation of ERK is mediated by the GAG chains, as phosphorylation of ERK is not identified when only the core protein of biglycan is given [128]. Bone marrow stroma cells (BMSCs) from biglycan/decorin-deficient mice had increased TGF-β signalling due to the inability of biglycan and decorin to sequester TGF-β in the ECM, resulting in a switch in fate from growth to apoptosis. In biglycan/decorin-deficient animals, the early death of osteogenic stem cells and osteoblast precursors resulted in a reduction in the number of mature osteoblasts, contributing to reduced osteogenesis and an osteoporosis-like phenotype. These investigations show that these two SLRPs function in controlling bone mass by modulating the proliferation and survival of osteogenic stem cells via regulating TGF-β activity. Due to a reduction in bone production, mice with a targeted disruption of biglycan develop age-dependent osteoporosis, with smaller trabecular volume and thinner cortices than their wild-type counterparts. Compared to wild-type littermates, biglycan knock-out mice have a considerably reduced capacity to form bone marrow stromal cells (Figure 3) [131].
The relevance of the biological role of PGs in skeletal development has been further demonstrated by using N-Ethyl-N-nitrosourea (ENU) mutagenesis screen that yielded zebrafish mutants that were deficient in cartilage and bone formation. Mutations in two genes, xylt1 and fam20b, were discovered using restriction site associated DNA (RAD) mapping, followed by meiotic mapping and sequencing in a class of mutants with reduced alcian blue staining of PGs in their cartilage matrix [36,132,133]. Xylt1 induces GAG side chain modifications to PG core proteins [134]. Fam20b is a kinase, phosphorylating xylose in the GAG side chain [135]. FAM20B is one of three fam20 family members with sequence similarity 20, also containing FAM20A and FAM20C in mammals. Fam20C is a kinase in this family that phosphorylates hundreds of secreted proteins with a highly conserved Ser-X-Glu/pSer motif. The pseudokinase FAM20A lacks a critical residue for catalysis and forms an evolutionarily conserved homodimer or heterodimer functional complex with FAM20C and activates it. Raine syndrome is caused by FAM20C mutations, which induce bone and craniofacial/dental anomalies, whereas Amelogenesis Imperfecta (AI) is caused by FAM20A mutations. Xylose kinases with a unique active site for binding Gal1-4Xyl1, the initiator disaccharide inside the tetrasaccharide linker region of PGs, are encoded by FAM20B. In HeLa cells, the overexpression of fam20b increases the quantity of HS and CS, whereas fam20b RNA interference diminishes their amount. This essential gene increases the number of GAG chains in PGs by phosphorylating the initiator xylose residue inside the tetrasaccharide linkage region. This phosphorylation is necessary for the elongation of the tetrasaccharide bridge and the assembly of GAG [136].
The discovery of zebrafish xylt1 and fam20b mutants with PG synthesis defects, followed by less cartilage matrix and early perichondral bone formation in developing embryos, led to the theory that cartilage PGs impede endochondral ossification. The fact that perichondral bone development begins sooner than wild-type siblings in these mutants indicates a physiologic function for cartilage PGs. Zebrafish with a fam20b gene mutation did not create wild-type amounts of CS and HS, typically prevalent in the cartilage matrix [35,36]. Moreover, in xylt1 and fam20b mutant chondrocytes, sox9a expression was reduced, whereas Runx2 transcripts increased. Ihh, which is expressed in mature chondrocytes but not in the perichondrium, mediates the inductive event for perichondral bone development. Ihh transcripts were upregulated early in xylt1 and fam20b mutant chondrocytes, and genetic epistasis tests revealed that Ihh function was required for these PG mutants’ early bone development [36].
4. PGs Modulate BMP Signal Transduction
In several in vivo and in vitro settings, BMPs have been discovered to interact with cell-surface and matrix-bound PGs, such perlecan, syndecan, glypican, and betaglycan [137,138]. For example, Xenopus syndecan-1 controls ectoderm dorsoventral patterning by modulating BMP signalling [139]. Glypican-3 has been linked to the modulation of the BMP-4 effects on renal branching morphogenesis [140]. In embryonic kidney explants, glypican-3 loss prevents BMP-2- and BMP-7-dependent ureteric bud formation [141]. In addition, betaglycan, a TGF-β type III receptor, is a membrane HS/CS sulfate PG discovered by its capacity to bind multiple TGF-β family members. Betaglycan can bind a broad spectrum of BMP ligands, including BMP-2, BMP-4, BMP-7, and GDF-5 [142].
PGs may have a key role in skeletal development by regulating BMP signalling (Figure 4). Perlecan works with BMP-2 to increase hypertrophic chondrocyte markers’ expression and osteogenesis [143,144]. Syndecan-3 has been shown to control BMPs activity during chondrogenesis by reducing the effective concentration of BMPs accessible for signalling [126]. Glypican-3 has been linked to the modulation of the BMP-4 effects on vertebrate limb patterning and skeletal development [140]. In vitro, a lack of biglycan inhibits BMP-4-induced osteoblast differentiation due to diminished BMP-4 binding to the receptors, which is totally restored by viral transfection of biglycan [129]. Biglycan has also been discovered to bind directly to BMP-2 and influence BMP-2-induced osteoblast differentiation [145]. In addition, BMP-2 biological activity is modulated by biglycan and decorin, which sequester it in the ECM. In BMSCs from mice with inactivated biglycan and decorin, the expression of BMP-2 signalling components, Smad 1, and phosphorylated Smad 1 were greater than in wild-type cells [131]. In articular chondrocytes, HS deficiency leads to chondrocyte hypertrophy, and enhanced BMP/Smad signalling plays a role in this phenotype [146]. Furthermore, it has been shown that heterozygous loss-of-function mutations in HS synthesis genes, EXT1 or EXT2, which cause multiple hereditary exostoses, a genetic bone condition, results in the upregulation of BMP signalling in the perichondrium. In vitro, BMP signalling is upregulated in Ext1-deficient perichondrium-derived mesenchymal progenitor cells (PDPCs), which leads to increased BMP-induced chondrogenic differentiation [147].
PGs have been discovered to have opposing effects on BMP activity, boosting it in some and decreasing it in others. For instance, HSPGs may either affect BMP activity by functioning as co-receptors, allowing BMPs and their receptors to interact more easily, or serve as a sink for BMPs, preventing them from interacting with BMP receptors, and consequently decrease rather than promote BMP signalling [126,148,149]. Division abnormally delayed (dally), a Drosophila homolog of the glypican family of GPI-linked cell-surface HSPGs, controls cell sensitivity to Dpp signalling by acting as a co-receptor for Dpp [150]. On the other hand, Dally can modulate Dpp distribution by sequestering it, assisting in the morphogen gradient in developing the wing disc [151]. In addition, BMP-4 biological activity is restricted in the Xenopus embryo when it binds to HS chains [152]. In vertebrate, polydactyly and other skeletal anomalies are associated with a lack of cellular response to BMP4 due to a loss-of-function mutation in glypican-3, which leads to Simpson–Golabi–Behmel dysmorphia syndrome in humans and mice [140]. Betaglycan functions as a BMP co-receptor, enhancing BMPs binding to the BMP signalling receptors, ALK3 and ALK6, and hence BMP signalling [142]. In vitro studies on a rat osteoblast cell line, ROS 17/2.8, shows that BMP-7 attaches to HS chains based on their sulfate structures, and the digestion of cell-surface HS hinders BMP-7 binding to the cells, which leads to inhibition of BMP-7-mediated Smad 1/5/8 phosphorylation [153]. In addition, natural BMP antagonists, such as Noggin and Chordin, bind to HS, which enhances their activity. HSPGs that bind to Noggin result in reduced BMP-4 activity [154,155]. Grem1, a member of the Dan family of BMP antagonists that directly binds BMPs to impede signal transduction, can also attach to HSPGs [156].
PG and BMP interactions are predominantly mediated by PG modifications and the domain’s amino acid sequence of BMP ligands. PG modifications affect cell signalling by changing the affinity of ligands and receptors. Sulf1, a secreted sulfatase enzyme, can alter the HSPG structure by removing a sulfate group from the 6-O position of glucosamine in HS chains at the cell surface [157,158,159]. In zebrafish, Sulf1 is an important regulator of BMP signalling that is necessary for appropriate somite development. In knock-down Sulf1 zebrafish embryos, the pharmacological suppression of BMP signalling rescues the development of the horizontal myoseptum and restores the normal migration of the posterior lateral line (PLL) primordium and pigment cells [160]. While HSPGs have well-established functional roles in BMP cell signalling, CSPGs’ participation in this process has often been overlooked. Nevertheless, an analysis of a gene trap mutation in the chondroitin-4-sulfotransferase 1 (C4st1) gene that causes the downregulation of 4-O-sulfated chondroitin synthesis leads to a severe chondrodysplasia associated with the downregulation of BMP signalling [108]. On the other hand, domains rich in basic amino acids govern BMP ligands’ distribution and function. One such domain has been found in the n-terminal region of BMP-2 and BMP-4. Changing the basic residues in the n-terminal domains of BMP-2 and BMP-4 inhibits HS binding and changes the mutant proteins’ bioactivity [138,152,161]. Furthermore, a domain with significant HS-binding capacity is in the C-terminal sections of mature BMP-5, BMP-6, and BMP-7. This domain’s amino acid sequence is nearly similar in all three BMPs and has been extremely conserved throughout evolution [162].
5. Exogenous Mechanisms to Modulate BMP Signalling Pathway
A variety of experimental techniques to modify the BMP signalling pathway have been developed in recent years. According to these findings, BMP signalling can be perturbed by powerful tools and at various molecular levels including: (1) ligand traps and natural soluble antagonists to limit ligand availability to receptors, (2) BMP receptors’ expression or kinase activity suppressors, and (3) intracellular inhibitors [163,164,165,166,167]. In this review, we will concentrate on a group of highly selective small-molecule inhibitors, including dorsomorphin and its analogs, which specifically antagonize the intracellular kinase domain of BMP type I receptors (Figure 4).
Figure 4.
Endogenous and exogenous modulation of BMP signalling transduction by PGs and dorsomorphin analogs. PGs have been shown to regulate the activity or availability of BMPs, either sequestering them from or presenting them to target cell receptors. Dorsomorphin (DM) and its analogs, LDN-193189 (LDN), DMH1, DMH2, and DMH3, specifically antagonize the intracellular kinase domain of BMP type I receptors. “Created with BioRender.com (Accessed on 31 January 2022)”.
Figure 4.
Endogenous and exogenous modulation of BMP signalling transduction by PGs and dorsomorphin analogs. PGs have been shown to regulate the activity or availability of BMPs, either sequestering them from or presenting them to target cell receptors. Dorsomorphin (DM) and its analogs, LDN-193189 (LDN), DMH1, DMH2, and DMH3, specifically antagonize the intracellular kinase domain of BMP type I receptors. “Created with BioRender.com (Accessed on 31 January 2022)”.

Dorsomorphin, also known as Compound C, was the first effective small-molecule BMP type I receptor inhibitor, which was found in a 7500-compound zebrafish library screen based on its ability to disrupt dorsoventral patterning in early zebrafish [168]. Dorsomorphin treatment phenocopies the effect of BMP antagonists, such as Noggin, Chordin, and Follistatin, which induce embryonic dorsalization endogenously. Dorsomorphin’s heterocyclic core structure interacts with the ATP binding site in the kinase domain of BMP type I receptors (Alk2, Alk3, and Alk6) with varying affinities, inhibiting their kinase activity [169,170,171]. The heterocyclic core of dorsomorphin, pyrazolo[1,5-a]pyrimidine core, has been used to synthesize multiple generations of dorsomorphin derivatives, including LDN-193189, DMH1, DMH2, JL5, DMH3, and DMH4. The changes made at the R-positions, R1 and R2, of the pyrazole[1,5-a]pyrimidine core varied amongst analogs of dorsomorphin, resulting in different affinities to the kinase domain of the BMP type I and type II receptors (Table 1) [172].
Based primarily on its effects on zebrafish embryos, LDN-193189 was found as a new compound with enhanced pharmacokinetic characteristics in a structure–activity relationship (SAR) test of dorsomorphin analogs. LDN-193189 has a significantly better selectivity for BMP receptors than dorsomorphin and can be utilised at lower dosages. The signalling activity of the BMP type I receptors ALK2 and ALK3 was effectively suppressed by LDN-193189 [173]. BMP-4 induction of ALP activity is inhibited by LDN-193189 in the C2C12 cell line, which expresses BMP receptors and can be induced to become osteoblasts by BMPs. This indicates that LDN-193189 may influence BMP-induced osteoblast differentiation [174,175]. In another study, LDN-193189 was utilised to demonstrate that overactive BMP receptor signalling contributes to the development of Fibrodysplasia ossificans progressiva (FOP) [176]. FOP is a rare, severe developmental musculoskeletal disease that is caused by activating mutations of the ALK2 gene and marked by heterotopic ossification (HO), endochondral bone development in non-skeletal sites, and a congenital deformity of the great toe [177,178,179]. Intraperitoneal administration of LDN-193189 could partially prevent heterotopic ossification without inducing osteopenia or bone fractures in a mouse model of FOP characterised by a constitutively active version of ALK2 [176]. Furthermore, in Col2a1-Ext1CKO and Fsp1-Ext1CKO, the two multiple hereditary exostoses models, the pharmacological suppression of BMP signalling by LDN-193189 reduces osteochondromagenesis [147]. Both dorsomorphin and LDN-193189 are known to have a variety of “off-target” effects. In addition to inhibiting BMP type I receptor kinase activity, dorsomorphin and LDN-193189 inhibit other cellular kinases, including the TGF-β pathway, AMP-activated kinase (AMPK), receptor tyrosine kinases for platelet-derived growth factor (PDGFR), VEGF, and many other kinases. These off-target effects restrict these drugs’ therapeutic potential and value [180]. The disadvantageous “off-target” effects of dorsomorphin and LDN-193189 were overcome by the identification of dorsomorphin homolog 1 (DMH1), a highly selective small-molecule BMP inhibitor, in a second in vivo zebrafish dorsalization assay with 21 dorsomorphin analogs to generate specific BMP receptor kinase inhibitors. DMH1 inhibits BMP receptors, but not the VEGF pathway, so DMH1 dorsalizes the embryonic axis without disrupting angiogenic processes. Furthermore, DMH1 had no inhibitory effects on TGF-β, activin-induced Smad 2/3 activation, KDR (VEGFR2), ALK5, AMPK, or PDGFR. DMH1 blocks signalling through the ALK2 and ALK3 receptors, with no impact on the ALK6 receptor and no additional side effects [172,181].
DMH2 and DMH3 also belong to this family and are expected to be pan-type I BMP receptor inhibitors with fewer side effects than dorsomorphin and LDN-193189. DMH2 has been reported as the most effective dorsalizing chemical; however, it was less selective than DMH1 and DHM3, since greater doses produced nonspecific developmental effects. JL5 is a DMH2 analog that has better pharmacokinetic features than DMH2. Although all of these BMP inhibitors block BMP type I receptors, JL5 and DMH2 also inhibit BMPR2, while DMH1 and LDN-193189 have no effect on BMPR2 [182,183]. The anti-angiogenic effect of this family’s DMH4 member was elevated, whereas dorsomorphin‘s dorsalization activity was decreased [172]. Dorsomorphin derivatives are inhibitors of BMP signalling pathways and Id family members. Studies have shown that dorsomorphin and DMH1 inhibit BMP-induced Smad 1/5/8 pathway phosphorylation in a dose-dependent manner, while they have no impact on BMP-induced p38 activation [170,172]. However, other studies have shown that all known BMP-induced signallings, canonical and noncanonical pathways, are affected by dorsomorphin and, more effectively, LDN-193189 inhibition of BMP receptors [184]. Moreover, dorsomorphin, DMH1, and DMH2 dramatically reduced Id1, Id2, and Id3 expression, which are the direct mediators of BMP signalling [185]. Other studies have shown reduced BMP-2 and BMP-4 mRNA expression by DMH1, but not ALK2 or ALK3 mRNA expression [186].
6. Conclusions and Perspectives
The highly spaciotemporal regulations of skeletal development imply that the mechanisms controlling cell behavior during chondrogenesis and bone formation must be regulated precisely. In this review, we concluded that BMPs and PGs act in concert to orchestrate proper skeletogenesis. Changes in BMP ligands’ availability, as well as PGs’ quantity or structure, can have devastating effects on cartilage maturation and osteoblast differentiation in the embryo. Understanding the BMP signalling system and PGs’ interaction is essential to understanding skeletogenesis mechanisms and for the development of future treatments for osteochondrodysplasias. The data summarized in this review indicate that more research is needed to determine the mechanism of BMPs’ and PGs’ interaction during cartilage maturation, osteoblast differentiation, and its effectiveness for proper skeletal development. Furthermore, while HSPGs have well-established functional roles in BMP cell signalling, CSPGs participation in this process has often been overlooked. In future studies, a greater focus should be given to determine the impact of mutation in PG synthesis genes on the diffusion of BMPs in ECM and their effects in skeletal development. In addition, further research is needed to determine the extent to which BMPs use canonical pathways, noncanonical pathways, or both in different phases of chondrogenesis and bone formation. It will also be intriguing to fathom the situations in which these pathways function synergistically or antagonistically. It is also a paucity of information about the effects of highly selective BMP signalling inhibitors, such as DMH1, on skeletal development. The discovery of BMP signalling inhibitors can pave the way for a deeper understanding of skeletogenesis mechanisms and the development of effective new treatments for skeletal diseases.
Funding
This research received no external funding.
Acknowledgments
This work was funded by the College of Medicine at the University of Saskatchewan and Canadian Institutes of Health Research (CIHR) grant PJT 148683. The authors declare no potential conflict of interest with respect to the authorship and/or publication of this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Urist, M.R. Bone: Formation by autoinduction. Science 1965, 150, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Zhao, M.; Mundy, G.R. Bone morphogenetic proteins. Growth Factors 2004, 22, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Wozney, J.M. Overview of bone morphogenetic proteins. Spine 2002, 27, S2–S8. [Google Scholar] [CrossRef] [PubMed]
- Newfeld, S.J.; Wisotzkey, R.G.; Kumar, S. Molecular evolution of a developmental pathway: Phylogenetic analyses of transforming growth factor-β family ligands, receptors and Smad signal transducers. Genetics 1999, 152, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Aono, A.; Hazama, M.; Notoya, K.; Taketomi, S.; Yamasaki, H.; Tsukuda, R.; Sasaki, S.; Fujisawa, Y. Potent ectopic bone-inducing activity of bone morphogenetic protein-4/7 heterodimer. Biochem. Biophys. Res. Commun. 1995, 210, 670–677. [Google Scholar] [CrossRef]
- Israel, D.I.; Nove, J.; Kerns, K.M.; Kaufman, R.J.; Rosen, V.; Cox, K.A.; Wozney, J.M. Heterodimeric bone morphogenetic proteins show enhanced activity in vitro and in vivo. Growth Factors 1996, 13, 291–300. [Google Scholar] [CrossRef]
- Kaito, T.; Morimoto, T.; Mori, Y.; Kanayama, S.; Makino, T.; Takenaka, S.; Sakai, Y.; Otsuru, S.; Yoshioka, Y.; Yoshikawa, H. BMP-2/7 heterodimer strongly induces bone regeneration in the absence of increased soft tissue inflammation. Spine J. 2018, 18, 139–146. [Google Scholar] [CrossRef]
- Gomez-Puerto, M.C.; Iyengar, P.V.; García de Vinuesa, A.; ten Dijke, P.; Sanchez-Duffhues, G. Bone morphogenetic protein receptor signal transduction in human disease. J. Pathol. 2019, 247, 9–20. [Google Scholar] [CrossRef]
- Olsen, O.E.; Wader, K.F.; Hella, H.; Mylin, A.K.; Turesson, I.; Nesthus, I.; Waage, A.; Sundan, A.; Holien, T. Activin A inhibits BMP-signaling by binding ACVR2A and ACVR2B. Cell Commun. Signal. 2015, 13, 27. [Google Scholar] [CrossRef] [Green Version]
- Mueller, T.D.; Nickel, J. Promiscuity and specificity in BMP receptor activation. FEBS Lett. 2012, 586, 1846–1859. [Google Scholar] [CrossRef]
- Scharpfenecker, M.; van Dinther, M.; Liu, Z.; van Bezooijen, R.L.; Zhao, Q.; Pukac, L.; Löwik, C.W.; ten Dijke, P. BMP-9 signals via ALK1 and inhibits bFGF-induced endothelial cell proliferation and VEGF-stimulated angiogenesis. J. Cell Sci. 2007, 120, 964–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebisawa, T.; Tada, K.; Kitajima, I.; Tojo, K.; Sampath, T.K.; Kawabata, M.; Miyazono, K.; Imamura, T. Characterization of bone morphogenetic protein-6 signaling pathways in osteoblast differentiation. J. Cell Sci. 1999, 112, 3519–3527. [Google Scholar] [CrossRef] [PubMed]
- Ten Dijke, P.; Yamashita, H.; Sampath, T.K.; Reddi, A.H.; Estevez, M.; Riddle, D.L.; Ichijo, H.; Heldin, C.-H.; Miyazono, K. Identification of type I receptors for osteogenic protein-1 and bone morphogenetic protein-4. J. Biol. Chem. 1994, 269, 16985–16988. [Google Scholar] [CrossRef]
- De Caestecker, M. The transforming growth factor-β superfamily of receptors. Cytokine Growth Factor Rev. 2004, 15, 1–11. [Google Scholar] [CrossRef]
- Derynck, R.; Zhang, Y.E. Smad-dependent and Smad-independent pathways in TGF-β family signalling. Nature 2003, 425, 577–584. [Google Scholar] [CrossRef]
- Massagué, J.; Seoane, J.; Wotton, D. Smad transcription factors. Genes Dev. 2005, 19, 2783–2810. [Google Scholar] [CrossRef] [Green Version]
- Goldring, M.B.; Tsuchimochi, K.; Ijiri, K. The control of chondrogenesis. J. Cell. Biochem. 2006, 97, 33–44. [Google Scholar] [CrossRef]
- Ijiri, K.; Zerbini, L.F.; Peng, H.; Correa, R.G.; Lu, B.; Walsh, N.; Zhao, Y.; Taniguchi, N.; Huang, X.-L.; Otu, H.; et al. A novel role for GADD45β as a mediator of MMP-13 gene expression during chondrocyte terminal differentiation. J. Biol. Chem. 2005, 280, 38544–38555. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, R.; Hata, K.; Ikeda, F.; Matsubara, T.; Yamashita, K.; Ichida, F.; Yoneda, T. The role of Smads in BMP signaling. Front. Biosci. 2003, 8, s275–s284. [Google Scholar] [CrossRef] [Green Version]
- Nohe, A.; Keating, E.; Knaus, P.; Petersen, N.O. Signal transduction of bone morphogenetic protein receptors. Cell. Signal. 2004, 16, 291–299. [Google Scholar] [CrossRef]
- Rosen, V. BMP2 signaling in bone development and repair. Cytokine Growth Factor Rev. 2009, 20, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Dietz, H.C.; Pyeritz, R.E. Mutations in the human gene for fibrillin-1 (FBN1) in the Marfan syndrome and related disorders. Hum. Mol. Genet. 1995, 4 (Suppl. 1), 1799–1809. [Google Scholar] [CrossRef] [PubMed]
- Gregory, K.E.; Ono, R.N.; Charbonneau, N.L.; Kuo, C.-L.; Keene, D.R.; Bächinger, H.P.; Sakai, L.Y. The prodomain of BMP-7 targets the BMP-7 complex to the extracellular matrix. J. Biol. Chem. 2005, 280, 27970–27980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, L.Y.; Keene, D.R.; Engvall, E. Fibrillin, a new 350-kD glycoprotein, is a component of extracellular microfibrils. J. Cell Biol. 1986, 103, 2499–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Harris, R.E.; Bayston, L.J.; Ashe, H.L. Type IV collagens regulate BMP signalling in Drosophila. Nature 2008, 455, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Abreu, J.G.; Coffinier, C.; Larraın, J.; Oelgeschläger, M.; De Robertis, E. Chordin-like CR domains and the regulation of evolutionarily conserved extracellular signaling systems. Gene 2002, 287, 39–47. [Google Scholar] [CrossRef]
- Andhare, R.; Takahashi, N.; Knudson, W.; Knudson, C. Hyaluronan promotes the chondrocyte response to BMP-7. Osteoarthr. Cartil. 2009, 17, 906–916. [Google Scholar] [CrossRef] [Green Version]
- Peterson, R.S.; Andhare, R.A.; Rousche, K.T.; Knudson, W.; Wang, W.; Grossfield, J.B.; Thomas, R.O.; Hollingsworth, R.E.; Knudson, C.B. CD44 modulates Smad1 activation in the BMP-7 signaling pathway. J. Cell Biol. 2004, 166, 1081–1091. [Google Scholar] [CrossRef]
- Selbi, W.; De La Motte, C.; Hascall, V.; Phillips, A. BMP-7 modulates hyaluronan-mediated proximal tubular cell-monocyte interaction. J. Am. Soc. Nephrol. 2004, 15, 1199–1211. [Google Scholar] [CrossRef] [Green Version]
- Erlebacher, A.; Filvaroff, E.H.; Gitelman, S.E.; Derynck, R. Toward a molecular understanding of skeletal development. Cell 1995, 80, 371–378. [Google Scholar] [CrossRef] [Green Version]
- Olsen, B.R.; Reginato, A.M.; Wang, W. Bone Development. Annu. Rev. Cell Dev. Biol. 2000, 16, 191–220. [Google Scholar] [CrossRef] [PubMed]
- Soltanoff, C.S.; Chen, W.; Yang, S.; Li, Y.-P. Signaling networks that control the lineage commitment and differentiation of bone cells. Crit. Rev. Eukaryot. Gene Expr. 2009, 19, 1–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplan, A. Endochondral bone formation: The lineage cascade. Mech. Bone Dev. Growth 1994, 1–46. [Google Scholar]
- Cormack, D.H.; Ham, A.W. Ham’s Histology; Lippincott: New York, NY, USA, 1987. [Google Scholar]
- Eames, B.F.; De La Fuente, L.; Helms, J.A. Molecular ontogeny of the skeleton. Birth Defects Res. Part C Embryo Today Rev. 2003, 69, 93–101. [Google Scholar] [CrossRef]
- Eames, B.F.; Yan, Y.-L.; Swartz, M.E.; Levic, D.S.; Knapik, E.W.; Postlethwait, J.H.; Kimmel, C.B. Mutations in fam20b and xylt1 reveal that cartilage matrix controls timing of endochondral ossification by inhibiting chondrocyte maturation. PLoS Genet. 2011, 7, e1002246. [Google Scholar] [CrossRef] [Green Version]
- Kronenberg, H.M. Developmental regulation of the growth plate. Nature 2003, 423, 332–336. [Google Scholar] [CrossRef]
- Hendriks, M.; Ramasamy, S.K. Blood vessels and vascular niches in bone development and physiological remodeling. Front. Cell Dev. Biol. 2020, 8, 1481. [Google Scholar] [CrossRef]
- Gerber, H.-P.; Vu, T.H.; Ryan, A.M.; Kowalski, J.; Werb, Z.; Ferrara, N. VEGF couples hypertrophic cartilage remodeling, ossification and angiogenesis during endochondral bone formation. Nat. Med. 1999, 5, 623–628. [Google Scholar] [CrossRef]
- Cheng, H.; Jiang, W.; Phillips, F.M.; Haydon, R.C.; Peng, Y.; Zhou, L.; Luu, H.H.; An, N.; Breyer, B.; Vanichakarn, P.; et al. Osteogenic activity of the fourteen types of human bone morphogenetic proteins (BMPs). J. Bone Jt. Surg. 2003, 85, 1544–1552. [Google Scholar] [CrossRef]
- Satoh, A.; Suzuki, M.; Amano, T.; Tamura, K.; Ide, H. Joint development in Xenopus laevis and induction of segmentations in regenerating froglet limb (spike). Dev. Dyn. 2005, 233, 1444–1453. [Google Scholar] [CrossRef] [PubMed]
- Dudley, A.T.; Robertson, E.J. Overlapping expression domains of bone morphogenetic protein family members potentially account for limited tissue defects in BMP7 deficient embryos. Dev. Dyn. 1997, 208, 349–362. [Google Scholar] [CrossRef]
- Kingsley, D.M. What do BMPs do in mammals? Clues from the mouse short-ear mutation. Trends Genet. 1994, 10, 16–21. [Google Scholar] [CrossRef]
- Lyons, K.M.; Hogan, B.L.; Robertson, E.J. Colocalization of BMP 7 and BMP 2 RNAs suggests that these factors cooperatively mediate tissue interactions during murine development. Mech. Dev. 1995, 50, 71–83. [Google Scholar] [CrossRef]
- McMahon, J.A.; Takada, S.; Zimmerman, L.B.; Fan, C.-M.; Harland, R.M.; McMahon, A.P. Noggin-mediated antagonism of BMP signaling is required for growth and patterning of the neural tube and somite. Genes Dev. 1998, 12, 1438–1452. [Google Scholar] [CrossRef] [Green Version]
- Winnier, G.; Blessing, M.; Labosky, P.A.; Hogan, B. Bone morphogenetic protein-4 is required for mesoderm formation and patterning in the mouse. Genes Dev. 1995, 9, 2105–2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, J.; Hall, B.K. Chondrogenic cell differentiation from membrane bone periostea. Anat. Embryol. 1997, 196, 349–362. [Google Scholar] [CrossRef]
- Nakashima, K.; de Crombrugghe, B. Transcriptional mechanisms in osteoblast differentiation and bone formation. Trends Genet. 2003, 19, 458–466. [Google Scholar] [CrossRef]
- Nakashima, K.; Zhou, X.; Kunkel, G.; Zhang, Z.; Deng, J.M.; Behringer, R.R.; De Crombrugghe, B. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell 2002, 108, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.C.; Hoang, B.; Thomas, J.T.; Vukicevic, S.; Luyten, F.P.; Ryba, N.; Kozak, C.A.; Reddi, A.H.; Moos, M. Cartilage-derived morphogenetic proteins. New members of the transforming growth factor-beta superfamily predominantly expressed in long bones during human embryonic development. J. Biol. Chem. 1994, 269, 28227–28234. [Google Scholar] [CrossRef]
- Jones, C.M.; Lyons, K.M.; Hogan, B. Involvement of Bone Morphogenetic Protein-4 (BMP-4) and Vgr-1 in morphogenesis and neurogenesis in the mouse. Development 1991, 111, 531–542. [Google Scholar] [CrossRef]
- Lyons, K.M.; Pelton, R.; Hogan, B. Organogenesis and pattern formation in the mouse: RNA distribution patterns suggest a role for bone morphogenetic protein-2A (BMP-2A). Development 1990, 109, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Macias, D.; Ganan, Y.; Sampath, T.; Piedra, M.; Ros, M.; Hurle, J. Role of BMP-2 and OP-1 (BMP-7) in programmed cell death and skeletogenesis during chick limb development. Development 1997, 124, 1109–1117. [Google Scholar] [CrossRef] [PubMed]
- Minina, E.; Schneider, S.; Rosowski, M.; Lauster, R.; Vortkamp, A. Expression of Fgf and Tgfβ signaling related genes during embryonic endochondral ossification. Gene Expr. Patterns 2005, 6, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Wozney, J.M.; Rosen, V.; Celeste, A.J.; Mitsock, L.M.; Whitters, M.J.; Kriz, R.W.; Hewick, R.M.; Wang, E.A. Novel regulators of bone formation: Molecular clones and activities. Science 1988, 242, 1528–1534. [Google Scholar] [CrossRef] [PubMed]
- Settle, S.H., Jr.; Rountree, R.B.; Sinha, A.; Thacker, A.; Higgins, K.; Kingsley, D.M. Multiple joint and skeletal patterning defects caused by single and double mutations in the mouse Gdf6 and Gdf5 genes. Dev. Biol. 2003, 254, 116–130. [Google Scholar] [CrossRef] [Green Version]
- Storm, E.E.; Kingsley, D.M. GDF5 coordinates bone and joint formation during digit development. Dev. Biol. 1999, 209, 11–27. [Google Scholar] [CrossRef] [Green Version]
- Wolfman, N.M.; Hattersley, G.; Cox, K.; Celeste, A.J.; Nelson, R.; Yamaji, N.; Dube, J.L.; DiBlasio-Smith, E.; Nove, J.; Song, J.J.; et al. Ectopic induction of tendon and ligament in rats by growth and differentiation factors 5, 6, and 7, members of the TGF-beta gene family. J. Clin. Investig. 1997, 100, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Minina, E.; Wenzel, H.M.; Kreschel, C.; Karp, S.; Gaffield, W.; McMahon, A.P.; Vortkamp, A. BMP and Ihh/PTHrP signaling interact to coordinate chondrocyte proliferation and differentiation. Development 2001, 128, 4523–4534. [Google Scholar] [CrossRef]
- Minina, E.; Kreschel, C.; Naski, M.C.; Ornitz, D.M.; Vortkamp, A. Interaction of FGF, Ihh/Pthlh, and BMP signaling integrates chondrocyte proliferation and hypertrophic differentiation. Dev. Cell 2002, 3, 439–449. [Google Scholar] [CrossRef] [Green Version]
- Ohba, S. Hedgehog signaling in endochondral ossification. J. Dev. Biol. 2016, 4, 20. [Google Scholar] [CrossRef]
- Long, F.; Chung, U.-I.; Ohba, S.; McMahon, J.; Kronenberg, H.M.; McMahon, A.P. Ihh signaling is directly required for the osteoblast lineage in the endochondral skeleton. Development 2004, 131, 1309–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St-Jacques, B.; Hammerschmidt, M.; McMahon, A.P. Indian hedgehog signaling regulates proliferation and differentiation of chondrocytes and is essential for bone formation. Genes Dev. 1999, 13, 2072–2086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seki, K.; Hata, A. Indian hedgehog gene is a target of the bone morphogenetic protein signaling pathway. J. Biol. Chem. 2004, 279, 18544–18549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathi, S.; Rutenberg, J.B.; Johnson, R.L.; Vortkamp, A. Interaction of Ihh and BMP/Noggin signaling during cartilage differentiation. Dev. Biol. 1999, 209, 239–253. [Google Scholar] [CrossRef] [Green Version]
- Kawai, S.; Sugiura, T. Characterization of human bone morphogenetic protein (BMP)-4 and -7 gene promoters: Activation of BMP promoters by Gli, a sonic hedgehog mediator. Bone 2001, 29, 54–61. [Google Scholar] [CrossRef]
- Katagiri, T.; Yamaguchi, A.; Komaki, M.; Abe, E.; Takahashi, N.; Ikeda, T.; Rosen, V.; Wozney, J.M.; Fujisawa-Sehara, A.; Suda, T. Bone morphogenetic protein-2 converts the differentiation pathway of C2C12 myoblasts into the osteoblast lineage. J. Cell Biol. 1994, 127, 1755–1766. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, A.; Katagiri, T.; Ikeda, T.; Wozney, J.M.; Rosen, V.; Wang, E.A.; Kahn, A.J.; Suda, T.; Yoshiki, S. Recombinant human bone morphogenetic protein-2 stimulates osteoblastic maturation and inhibits myogenic differentiation in vitro. J. Cell Biol. 1991, 113, 681–687. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, A.; Komori, T.; Suda, T. Regulation of osteoblast differentiation mediated by bone morphogenetic proteins, hedgehogs, and Cbfa1. Endocr. Rev. 2000, 21, 393–411. [Google Scholar] [CrossRef]
- Chen, G.; Deng, C.; Li, Y.-P. TGF-β and BMP signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272. [Google Scholar] [CrossRef] [Green Version]
- Dorman, L.J.; Tucci, M.; Benghuzzi, H. In vitro effects of bmp-2, bmp-7, and bmp-13 on proliferation and differentation of mouse mesenchymal stem cells. Biomed. Sci. Instrum. 2012, 48, 81–87. [Google Scholar]
- Hata, K.; Nishimura, R.; Ikeda, F.; Yamashita, K.; Matsubara, T.; Nokubi, T.; Yoneda, T. Differential roles of Smad1 and p38 kinase in regulation of peroxisome proliferator-activating receptor γ during bone morphogenetic protein 2-induced adipogenesis. Mol. Biol. Cell 2003, 14, 545–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Q.; Song, W.-X.; Luo, Q.; Tang, N.; Luo, J.; Luo, X.; Chen, J.; Bi, Y.; He, B.-C.; Park, J.K.; et al. A comprehensive analysis of the dual roles of BMPs in regulating adipogenic and osteogenic differentiation of mesenchymal progenitor cells. Stem Cells Dev. 2009, 18, 545–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Ren, P.-G.; Ma, T.; Smith, R.L.; Goodman, S.B. Modulating osteogenesis of mesenchymal stem cells by modifying growth factor availability. Cytokine 2010, 51, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-S.; Kim, H.-J.; Li, Q.-L.; Chi, X.-Z.; Ueta, C.; Komori, T.; Wozney, J.M.; Kim, E.-G.; Choi, J.-Y.; Ryoo, H.-M.; et al. Runx2 is a common target of transforming growth factor β1 and bone morphogenetic protein 2, and cooperation between Runx2 and Smad5 induces osteoblast-specific gene expression in the pluripotent mesenchymal precursor cell line C2C12. Mol. Cell. Biol. 2000, 20, 8783–8792. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Javed, A.; Kim, H.J.; Shin, H.I.; Gutierrez, S.; Choi, J.Y.; Rosen, V.; Stein, J.L.; Van Wijnen, A.J.; Stein, G.S.; et al. Transient upregulation of CBFA1 in response to bone morphogenetic protein-2 and transforming growth factor β1 in C2C12 myogenic cells coincides with suppression of the myogenic phenotype but is not sufficient for osteoblast differentiation. J. Cell. Biochem. 1999, 73, 114–125. [Google Scholar] [CrossRef]
- Lee, M.-H.; Kim, Y.-J.; Kim, H.-J.; Park, H.-D.; Kang, A.-R.; Kyung, H.-M.; Sung, J.-H.; Wozney, J.M.; Ryoo, H.-M. BMP-2-induced Runx2 expression is mediated by Dlx5, and TGF-β1 opposes the BMP-2-induced osteoblast differentiation by suppression of Dlx5 expression. J. Biol. Chem. 2003, 278, 34387–34394. [Google Scholar] [CrossRef] [Green Version]
- Luo, T.; Matsuo-Takasaki, M.; Lim, J.; Sargent, T.D. Differential regulation of Dlx gene expression by a BMP morphogenetic gradient. Int. J. Dev. Biol. 2004, 45, 681–684. [Google Scholar]
- Miyama, K.; Yamada, G.; Yamamoto, T.S.; Takagi, C.; Miyado, K.; Sakai, M.; Ueno, N.; Shibuya, H. A BMP-inducible gene, dlx5, regulates osteoblast differentiation and mesoderm induction. Dev. Biol. 1999, 208, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Noël, D.; Gazit, D.; Bouquet, C.; Apparailly, F.; Bony, C.; Plence, P.; Millet, V.; Turgeman, G.; Perricaudet, M.; Sany, J.; et al. Short-term BMP-2 expression is sufficient for in vivo osteochondral differentiation of mesenchymal stem cells. Stem Cells 2004, 22, 74–85. [Google Scholar] [CrossRef]
- Gu, K.; Zhang, L.; Jin, T.; Rutherford, R.B. Identification of potential modifiers of Runx2/Cbfa1 activity in C2C12 cells in response to bone morphogenetic protein-7. Cells Tissues Organs 2004, 176, 28–40. [Google Scholar] [CrossRef]
- Shen, B.; Wei, A.; Whittaker, S.; Williams, L.A.; Tao, H.; Ma, D.D.; Diwan, A.D. The role of BMP-7 in chondrogenic and osteogenic differentiation of human bone marrow multipotent mesenchymal stromal cells in vitro. J. Cell. Biochem. 2010, 109, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Hardingham, T.E.; Fosang, A.J. Proteoglycans: Many forms and many functions. FASEB J. 1992, 6, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Iozzo, R.V.; Murdoch, A.D. Proteoglycans of the extracellular environment: Clues from the gene and protein side offer novel perspectives in molecular diversity and function. FASEB J. 1996, 10, 598–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wight, T.N.; Toole, B.P.; Hascall, V.C. Hyaluronan and the aggregating proteoglycans. In The Extracellular Matrix: An Overview; Mecham, R.P., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 147–195. [Google Scholar]
- Cortes, M.; Baria, A.T.; Schwartz, N.B. Sulfation of chondroitin sulfate proteoglycans is necessary for proper Indian hedgehog signaling in the developing growth plate. Development 2009, 136, 1697–1706. [Google Scholar] [CrossRef] [Green Version]
- Mouw, J.K.; Ou, G.; Weaver, V.M. Extracellular matrix assembly: A multiscale deconstruction. Nat. Rev. Mol. Cell Biol. 2014, 15, 771–785. [Google Scholar] [CrossRef]
- Hardingham, T.E.; MuIR, H. Binding of oligosaccharides of hyaluronic acid to proteoglycans. Biochem. J. 1973, 135, 905–908. [Google Scholar] [CrossRef] [Green Version]
- Kimata, K.; Oike, Y.; Tani, K.; Shinomura, T.; Yamagata, M.; Uritani, M.; Suzuki, S. A large chondroitin sulfate proteoglycan (PG-M) synthesized before chondrogenesis in the limb bud of chick embryo. J. Biol. Chem. 1986, 261, 13517–13525. [Google Scholar] [CrossRef]
- Schwartz, N.; Hennig, A.; Krueger, R., Jr.; Krzystolik, M.; Li, H.; Mangoura, D. Developmental expression of S103L cross-reacting proteoglycans in embryonic chick. Prog. Clin. Biol. Res. 1993, 383, 505–514. [Google Scholar]
- Kiani, C.; Liwen, C.; Wu, Y.J.; Albert, J.Y.; Burton, B.Y. Structure and function of aggrecan. Cell Res. 2002, 12, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Domowicz, M.S.; Cortes, M.; Henry, J.G.; Schwartz, N.B. Aggrecan modulation of growth plate morphogenesis. Dev. Biol. 2009, 329, 242–257. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Li, S.; Shi, Z.; Yue, Y.; Sun, J.; Chen, J.; Fu, Q.; Hughes, C.E.; Caterson, B. Articular cartilage metabolism in patients with Kashin–Beck disease: An endemic osteoarthropathy in China. Osteoarthr. Cartil. 2008, 16, 680–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Lei, R.; Tiainen, M.; Wu, S.; Zhang, Q.; Pei, F.; Guo, X. Disordered glycometabolism involved in pathogenesis of Kashin–Beck disease, an endemic osteoarthritis in China. Exp. Cell Res. 2014, 326, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Gleghorn, L.; Ramesar, R.; Beighton, P.; Wallis, G. A mutation in the variable repeat region of the aggrecan gene (AGC1) causes a form of spondyloepiphyseal dysplasia associated with severe, premature osteoarthritis. Am. J. Hum. Genet. 2005, 77, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Tompson, S.W.; Merriman, B.; Funari, V.A.; Fresquet, M.; Lachman, R.S.; Rimoin, D.L.; Nelson, S.F.; Briggs, M.D.; Cohn, D.H.; Krakow, D. A recessive skeletal dysplasia, SEMD aggrecan type, results from a missense mutation affecting the C-type lectin domain of aggrecan. Am. J. Hum. Genet. 2009, 84, 72–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krueger, R.C.; Kurima, K.; Schwartz, N.B. Completion of the mouse aggrecan gene structure and identification of the defect in the cmd-Bc mouse as a near complete deletion of the murine aggrecan gene. Mamm. Genome 1999, 10, 1119–1125. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Kimata, K.; Line, S.; Strong, D.; Gao, L.-Y.; Kozak, C.A.; Yamada, Y. Mouse cartilage matrix deficiency (cmd) caused by a 7 bp deletion in the aggrecan gene. Nat. Genet. 1994, 7, 154–157. [Google Scholar] [CrossRef]
- Watanabe, H.; Nakata, K.; Kimata, K.; Nakanishi, I.; Yamada, Y. Dwarfism and age-associated spinal degeneration of heterozygote cmd mice defective in aggrecan. Proc. Natl. Acad. Sci. USA 1997, 94, 6943–6947. [Google Scholar] [CrossRef] [Green Version]
- Yoo, T.; Cho, H.; Yamada, Y. Hearing impairment in mice with the cmd/cmd (cartilage matrix deficiency) mutant gene. Ann. N. Y. Acad. Sci. 1991, 630, 265–267. [Google Scholar] [CrossRef]
- Primorac, D.; Stover, M.; Clark, S.; Rowe, D. Molecular basis of nanomelia, a heritable chondrodystrophy of chicken. Matrix Biol. 1994, 14, 297–305. [Google Scholar] [CrossRef]
- Stirpe, N.S.; Argraves, W.S.; Goetinck, P.F. Chondrocytes from the cartilage proteoglycan-deficient mutant, nanomelia, synthesize greatly reduced levels of the proteoglycan core protein transcript. Dev. Biol. 1987, 124, 77–81. [Google Scholar] [CrossRef]
- Vertel, B.M.; Grier, B.L.; Li, H.; Schwartz, N.B. The chondrodystrophy, nanomelia: Biosynthesis and processing of the defective aggrecan precursor. Biochem. J. 1994, 301, 211–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vertel, B.M.; Walters, L.M.; Grier, B.; Maine, N.; Goetinck, P.F. Nanomelic chondrocytes synthesize, but fail to translocate, a truncated aggrecan precursor. J. Cell Sci. 1993, 104, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Benito-Arenas, R.; Doncel-Pérez, E.; Fernández-Gutiérrez, M.; Garrido, L.; García-Junceda, E.; Revuelta, J.; Bastida, A.; Fernández-Mayoralas, A. A holistic approach to unravelling chondroitin sulfation: Correlations between surface charge, structure and binding to growth factors. Carbohydr. Polym. 2018, 202, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Miyachi, K.; Wakao, M.; Suda, Y. Syntheses of chondroitin sulfate tetrasaccharide structures containing 4, 6-disulfate patterns and analysis of their interaction with glycosaminoglycan-binding protein. Bioorganic Med. Chem. Lett. 2015, 25, 1552–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadanaka, S.; Ishida, M.; Ikegami, M.; Kitagawa, H. Chondroitin 4-O-sulfotransferase-1 modulates Wnt-3a signaling through control of E disaccharide expression of chondroitin sulfate. J. Biol. Chem. 2008, 283, 27333–27343. [Google Scholar] [CrossRef] [Green Version]
- Klüppel, M.; Wight, T.N.; Chan, C.; Hinek, A.; Wrana, J.L. Maintenance of chondroitin sulfation balance by chondroitin-4-sulfotransferase 1 is required for chondrocyte development and growth factor signaling during cartilage morphogenesis. Development 2005, 132, 3989–4003. [Google Scholar] [CrossRef] [Green Version]
- Fenwick, S.; Gregg, P.; Kumar, S.; Smith, J.; Rooney, P. Intrinsic control of vascularization in developing cartilage rudiments. Int. J. Exp. Pathol. 1997, 78, 187–196. [Google Scholar] [CrossRef]
- Carlevaro, M.F.; Cermelli, S.; Cancedda, R.; Cancedda, F.D. Vascular endothelial growth factor (VEGF) in cartilage neovascularization and chondrocyte differentiation: Auto-paracrine role during endochondral bone formation. J. Cell Sci. 2000, 113, 59–69. [Google Scholar] [CrossRef]
- Grodzinsky, A.J. Electromechanical and physicochemical properties of connective tissue. Crit. Rev. Biomed. Eng. 1983, 9, 133–199. [Google Scholar]
- Mongiat, M.; Taylor, K.; Otto, J.; Aho, S.; Uitto, J.; Whitelock, J.M.; Iozzo, R.V. The protein core of the proteoglycan perlecan binds specifically to fibroblast growth factor-7. J. Biol. Chem. 2000, 275, 7095–7100. [Google Scholar] [CrossRef] [Green Version]
- Muddasani, P.; Norman, J.C.; Ellman, M.; Van Wijnen, A.J.; Im, H.-J. Basic fibroblast growth factor activates the MAPK and NFκB pathways that converge on Elk-1 to control production of matrix metalloproteinase-13 by human adult articular chondrocytes. J. Biol. Chem. 2007, 282, 31409–31421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nummenmaa, E.; Hämäläinen, M.; Moilanen, T.; Vuolteenaho, K.; Moilanen, E. Effects of FGF-2 and FGF receptor antagonists on MMP enzymes, aggrecan, and type II collagen in primary human OA chondrocytes. Scand. J. Rheumatol. 2015, 44, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Vincent, T.; McLean, C.; Full, L.; Peston, D.; Saklatvala, J. FGF-2 is bound to perlecan in the pericellular matrix of articular cartilage, where it acts as a chondrocyte mechanotransducer. Osteoarthr. Cartil. 2007, 15, 752–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansouri, R.; Jouan, Y.; Hay, E.; Blin-Wakkach, C.; Frain, M.; Ostertag, A.; Le Henaff, C.; Marty, C.; Geoffroy, V.; Marie, P.J.; et al. Correction: Osteoblastic heparan sulfate glycosaminoglycans control bone remodeling by regulating Wnt signaling and the crosstalk between bone surface and marrow cells. Cell Death Dis. 2018, 9, 788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, T.; Izumikawa, T.; Tamura, J.-I.; Kitagawa, H. Chondroitin sulfate-E fine-tunes osteoblast differentiation via ERK1/2, Smad3 and Smad1/5/8 signaling by binding to N-cadherin and cadherin-11. Biochem. Biophys. Res. Commun. 2012, 420, 523–529. [Google Scholar] [CrossRef]
- Choi, Y.; Chung, H.; Jung, H.; Couchman, J.R.; Oh, E.-S. Syndecans as cell surface receptors: Unique structure equates with functional diversity. Matrix Biol. 2011, 30, 93–99. [Google Scholar] [CrossRef]
- Xian, X.; Gopal, S.; Couchman, J.R. Syndecans as receptors and organizers of the extracellular matrix. Cell Tissue Res. 2010, 339, 31–46. [Google Scholar] [CrossRef]
- David, G.; Bai, X.M.; Van der Schueren, B.; Marynen, P.; Cassiman, J.-J.; Van den Berghe, H. Spatial and temporal changes in the expression of fibroglycan (syndecan-2) during mouse embryonic development. Development 1993, 119, 841–854. [Google Scholar] [CrossRef]
- Gutierrez, J.; Osses, N.; Brandan, E. Changes in secreted and cell associated proteoglycan synthesis during conversion of myoblasts to osteoblasts in response to bone morphogenetic protein-2: Role of decorin in cell response to BMP-2. J. Cell. Physiol. 2006, 206, 58–67. [Google Scholar] [CrossRef]
- Teplyuk, N.M.; Haupt, L.M.; Ling, L.; Dombrowski, C.; Mun, F.K.; Nathan, S.S.; Lian, J.B.; Stein, J.L.; Stein, G.S.; Cool, S.M. The osteogenic transcription factor Runx2 regulates components of the fibroblast growth factor/proteoglycan signaling axis in osteoblasts. J. Cell. Biochem. 2009, 107, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Dieudonné, F.X.; Marion, A.; Marie, P.J.; Modrowski, D. Targeted inhibition of T-cell factor activity promotes syndecan-2 expression and sensitization to doxorubicin in osteosarcoma cells and bone tumors in mice. J. Bone Miner. Res. 2012, 27, 2118–2129. [Google Scholar] [CrossRef] [PubMed]
- Marion, A.; Dieudonné, F.X.; Patiño-Garcia, A.; Lecanda, F.; Marie, P.J.; Modrowski, D. Calpain-6 is an endothelin-1 signaling dependent protective factor in chemoresistant osteosarcoma. Int. J. Cancer 2012, 130, 2514–2525. [Google Scholar] [CrossRef] [PubMed]
- Orosco, A.; Fromigué, O.; Haÿ, E.; Marie, P.J.; Modrowski, D. Dual involvement of protein kinase C δ in apoptosis induced by syndecan-2 in osteoblasts. J. Cell. Biochem. 2006, 98, 838–850. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Li, Y.; Seghatoleslami, M.R.; Dealy, C.N.; Kosher, R.A. Heparan sulfate proteoglycans including syndecan-3 modulate BMP activity during limb cartilage differentiation. Matrix Biol. 2006, 25, 27–39. [Google Scholar] [CrossRef]
- Iozzo, R.V. The family of the small leucine-rich proteoglycans: Key regulators of matrix assembly and cellular growth. Crit. Rev. Biochem. Mol. Biol. 1997, 32, 141–174. [Google Scholar] [CrossRef]
- Berendsen, A.D.; Fisher, L.W.; Kilts, T.M.; Owens, R.T.; Robey, P.G.; Gutkind, J.S.; Young, M.F. Modulation of canonical Wnt signaling by the extracellular matrix component biglycan. Proc. Natl. Acad. Sci. USA 2011, 108, 17022–17027. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.-D.; Fisher, L.W.; Robey, P.G.; Young, M.F. The small leucine-rich proteoglycan biglycan modulates BMP-4-induced osteoblast differentiation. FASEB J. 2004, 18, 948–958. [Google Scholar] [CrossRef]
- Young, M.F.; Bi, Y.; Ameye, L.; Chen, X.-D. Biglycan knockout mice: New models for musculoskeletal diseases. Glycoconj. J. 2002, 19, 257–262. [Google Scholar] [CrossRef]
- Bi, Y.; Stuelten, C.H.; Kilts, T.; Wadhwa, S.; Iozzo, R.V.; Robey, P.G.; Chen, X.-D.; Young, M.F. Extracellular matrix proteoglycans control the fate of bone marrow stromal cells. J. Biol. Chem. 2005, 280, 30481–30489. [Google Scholar] [CrossRef] [Green Version]
- Eames, B.F.; Singer, A.; Smith, G.A.; Wood, Z.A.; Yan, Y.-L.; He, X.; Polizzi, S.J.; Catchen, J.M.; Rodriguez-Mari, A.; Linbo, T.; et al. UDP xylose synthase 1 is required for morphogenesis and histogenesis of the craniofacial skeleton. Dev. Biol. 2010, 341, 400–415. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.R.; Atwood, T.S.; Eames, B.F.; Eberhart, J.K.; Yan, Y.-L.; Postlethwait, J.H.; Johnson, E.A. RAD marker microarrays enable rapid mapping of zebrafish mutations. Genome Biol. 2007, 8, R105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, N. Regulation of chondroitin sulfate synthesis. Effect of beta-xylosides on synthesis of chondroitin sulfate proteoglycan, chondroitin sulfate chains, and core protein. J. Biol. Chem. 1977, 252, 6316–6321. [Google Scholar] [CrossRef]
- Koike, T.; Izumikawa, T.; Tamura, J.-I.; Kitagawa, H. FAM20B is a kinase that phosphorylates xylose in the glycosaminoglycan–protein linkage region. Biochem. J. 2009, 421, 157–162. [Google Scholar] [CrossRef] [Green Version]
- Tagliabracci, V.S.; Engel, J.L.; Wiley, S.E.; Xiao, J.; Gonzalez, D.J.; Appaiah, H.N.; Koller, A.; Nizet, V.; White, K.E.; Dixon, J.E. Dynamic regulation of FGF23 by Fam20C phosphorylation, GalNAc-T3 glycosylation, and furin proteolysis. Proc. Natl. Acad. Sci. USA 2014, 111, 5520–5525. [Google Scholar] [CrossRef] [Green Version]
- Iozzo, R.V. Series Introduction: Heparan sulfate proteoglycans: Intricate molecules with intriguing functions. J. Clin. Investig. 2001, 108, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Ruppert, R.; Hoffmann, E.; Sebald, W. Human bone morphogenetic protein 2 contains a heparin-binding site which modifies its biological activity. Eur. J. Biochem. 1996, 237, 295–302. [Google Scholar] [CrossRef]
- Olivares, G.H.; Carrasco, H.; Aroca, F.; Carvallo, L.; Segovia, F.; Larraín, J. Syndecan-1 regulates BMP signaling and dorso-ventral patterning of the ectoderm during early Xenopus development. Dev. Biol. 2009, 329, 338–349. [Google Scholar] [CrossRef] [Green Version]
- Paine-Saunders, S.; Viviano, B.L.; Zupicich, J.; Skarnes, W.C.; Saunders, S. glypican-3 controls cellular responses to Bmp4 in limb patterning and skeletal development. Dev. Biol. 2000, 225, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Grisaru, S.; Cano-Gauci, D.; Tee, J.; Filmus, J.; Rosenblum, N.D. Glypican-3 modulates BMP-and FGF-mediated effects during renal branching morphogenesis. Dev. Biol. 2001, 231, 31–46. [Google Scholar] [CrossRef] [Green Version]
- Kirkbride, K.C.; Townsend, T.A.; Bruinsma, M.W.; Barnett, J.V.; Blobe, G.C.; Kawai, Y.; Morinaga, H.; Kondo, H.; Miyoshi, N.; Nakamura, Y.; et al. Bone morphogenetic proteins signal through the transforming growth factor-β type III receptor. J. Biol. Chem. 2008, 283, 7628–7637. [Google Scholar] [CrossRef] [Green Version]
- DeCarlo, A.A.; Belousova, M.; Ellis, A.L.; Petersen, D.; Grenett, H.; Hardigan, P.; O’Grady, R.; Lord, M.; Whitelock, J.M. Perlecan domain 1 recombinant proteoglycan augments BMP-2 activity and osteogenesis. BMC Biotechnol. 2012, 12, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, R.; Kirn-Safran, C.; Farach-Carson, M.; Carson, D. Perlecan: An important component of the cartilage pericellular matrix. J. Musculoskelet. Neuronal Interact. 2002, 2, 511. [Google Scholar] [PubMed]
- Mochida, Y.; Parisuthiman, D.; Yamauchi, M. Biglycan is a positive modulator of BMP-2 induced osteoblast differentiation. In Tissue Engineering; Springer: Boston, MA, USA, 2006; pp. 101–113. [Google Scholar]
- Kawashima, K.; Ogawa, H.; Komura, S.; Ishihara, T.; Yamaguchi, Y.; Akiyama, H.; Matsumoto, K. Heparan sulfate deficiency leads to hypertrophic chondrocytes by increasing bone morphogenetic protein signaling. Osteoarthr. Cartil. 2020, 28, 1459–1470. [Google Scholar] [CrossRef] [PubMed]
- Inubushi, T.; Nozawa, S.; Matsumoto, K.; Irie, F.; Yamaguchi, Y. Aberrant perichondrial BMP signaling mediates multiple osteochondroma genesis in mice. JCI Insight 2017, 2, e90049. [Google Scholar] [CrossRef] [Green Version]
- Jiao, X.; Billings, P.C.; O’Connell, M.P.; Kaplan, F.S.; Shore, E.M.; Glaser, D.L. Heparan sulfate proteoglycans (HSPGs) modulate BMP2 osteogenic bioactivity in C2C12 cells. J. Biol. Chem. 2007, 282, 1080–1086. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, Y.; Matsumoto, K.; Irie, F.; Fukushi, J.-I.; Stallcup, W.B.; Yamaguchi, Y. Conditional ablation of the heparan sulfate-synthesizing enzyme Ext1 leads to dysregulation of bone morphogenic protein signaling and severe skeletal defects. J. Biol. Chem. 2010, 285, 19227–19234. [Google Scholar] [CrossRef] [Green Version]
- Fujise, M.; Takeo, S.; Kamimura, K.; Matsuo, T.; Aigaki, T.; Izumi, S.; Nakato, H. Dally regulates Dpp morphogen gradient formation in the Drosophila wing. Development 2003, 130, 1515–1522. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, T.; Kamimura, K.; Firkus, C.; Takeo, S.; Shimmi, O.; Nakato, H. Dally regulates Dpp morphogen gradient formation by stabilizing Dpp on the cell surface. Dev. Biol. 2008, 313, 408–419. [Google Scholar] [CrossRef] [Green Version]
- Ohkawara, B.; Iemura, S.-I.; ten Dijke, P.; Ueno, N. Action range of BMP is defined by its N-terminal basic amino acid core. Curr. Biol. 2002, 12, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Irie, A.; Habuchi, H.; Kimata, K.; Sanai, Y. Heparan sulfate is required for bone morphogenetic protein-7 signaling. Biochem. Biophys. Res. Commun. 2003, 308, 858–865. [Google Scholar] [CrossRef]
- Jasuja, R.; Allen, B.L.; Pappano, W.N.; Rapraeger, A.C.; Greenspan, D.S. Cell-surface heparan sulfate proteoglycans potentiate chordin antagonism of bone morphogenetic protein signaling and are necessary for cellular uptake of chordin. J. Biol. Chem. 2004, 279, 51289–51297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paine-Saunders, S.; Viviano, B.L.; Economides, A.N.; Saunders, S. Heparan sulfate proteoglycans retain Noggin at the cell surface: A potential mechanism for shaping bone morphogenetic protein gradients. J. Biol. Chem. 2002, 277, 2089–2096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pegge, J.; Tatsinkam, A.J.; Rider, C.C.; Bell, E. Heparan sulfate proteoglycans regulate BMP signalling during neural crest induction. Dev. Biol. 2020, 460, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Ai, X.; Do, A.-T.; Kusche-Gullberg, M.; Lindahl, U.; Lu, K.; Emerson, C.P. Substrate specificity and domain functions of extracellular heparan sulfate 6-O-endosulfatases, QSulf1 and QSulf2. J. Biol. Chem. 2006, 281, 4969–4976. [Google Scholar] [CrossRef] [Green Version]
- Freeman, S.D.; Moore, W.M.; Guiral, E.C.; Holme, A.D.; Turnbull, J.E.; Pownall, M.E. Extracellular regulation of developmental cell signaling by XtSulf1. Dev. Biol. 2008, 320, 436–445. [Google Scholar] [CrossRef] [Green Version]
- Lai, J.-P.; Chien, J.; Strome, S.E.; Staub, J.; Montoya, D.P.; Greene, E.L.; Smith, D.I.; Roberts, L.R.; Shridhar, V. HSulf-1 modulates HGF-mediated tumor cell invasion and signaling in head and neck squamous carcinoma. Oncogene 2004, 23, 1439–1447. [Google Scholar] [CrossRef] [Green Version]
- Meyers, J.R.; Planamento, J.; Ebrom, P.; Krulewitz, N.; Wade, E.; Pownall, M.E. Sulf1 modulates BMP signaling and is required for somite morphogenesis and development of the horizontal myoseptum. Dev. Biol. 2013, 378, 107–121. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.J.; Lee, J.Y.; Park, J.H.; Park, J.B.; Suh, J.S.; Choi, Y.S.; Lee, S.J.; Chung, C.-P.; Park, Y.J. The identification of a heparin binding domain peptide from bone morphogenetic protein-4 and its role on osteogenesis. Biomaterials 2010, 31, 7226–7238. [Google Scholar] [CrossRef]
- Billings, P.C.; Yang, E.; Mundy, C.; Pacifici, M. Domains with highest heparan sulfate–binding affinity reside at opposite ends in BMP2/4 versus BMP5/6/7: Implications for function. J. Biol. Chem. 2018, 293, 14371–14383. [Google Scholar] [CrossRef] [Green Version]
- Cappato, S.; Tonachini, L.; Giacopelli, F.; Tirone, M.; Galietta, L.J.; Sormani, M.; Giovenzana, A.; Spinelli, A.E.; Canciani, B.; Brunelli, S. High-throughput screening for modulators of ACVR1 transcription: Discovery of potential therapeutics for fibrodysplasia ossificans progressiva. Dis. Models Mech. 2016, 9, 685–696. [Google Scholar] [CrossRef] [Green Version]
- Jinnin, M.; Ihn, H.; Tamaki, K. Characterization of SIS3, a novel specific inhibitor of Smad3, and its effect on transforming growth factor-β1-induced extracellular matrix expression. Mol. Pharmacol. 2006, 69, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Mura, M.; Cappato, S.; Giacopelli, F.; Ravazzolo, R.; Bocciardi, R. The role of the 3′ UTR region in the regulation of the ACVR1/Alk-2 gene expression. PLoS ONE 2012, 7, e50958. [Google Scholar] [CrossRef] [PubMed]
- Olsen, O.E.; Sankar, M.; Elsaadi, S.; Hella, H.; Buene, G.; Darvekar, S.R.; Misund, K.; Katagiri, T.; Knaus, P.; Holien, T. BMPR2 inhibits activin and BMP signaling via wild-type ALK2. J. Cell Sci. 2018, 131, jcs213512. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Cai, J.; de Gorter, D.J.; Sanchez-Duffhues, G.; Kemaladewi, D.U.; Hoogaars, W.M.; Aartsma-Rus, A.; ’t Hoen, P.A.; ten Dijke, P. Antisense-oligonucleotide mediated exon skipping in activin-receptor-like kinase 2: Inhibiting the receptor that is overactive in fibrodysplasia ossificans progressiva. PLoS ONE 2013, 8, e69096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammerschmidt, M.; Serbedzija, G.N.; McMahon, A.P. Genetic analysis of dorsoventral pattern formation in the zebrafish: Requirement of a BMP-like ventralizing activity and its dorsal repressor. Genes Dev. 1996, 10, 2452–2461. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, B.; Seibel, W. Compound C/dorsomorphin: Its use and misuse as an AMPK inhibitor. In AMPK; Humana Press: New York, NY, USA, 2018; pp. 195–202. [Google Scholar]
- Paul, B.Y.; Hong, C.C.; Sachidanandan, C.; Babitt, J.L.; Deng, D.Y.; Hoyng, S.A.; Lin, H.Y.; Bloch, K.D.; Peterson, R.T. Dorsomorphin inhibits BMP signals required for embryogenesis and iron metabolism. Nat. Chem. Biol. 2008, 4, 33–41. [Google Scholar]
- Wrighton, K.H.; Lin, X.; Paul, B.Y.; Feng, X.-H. Transforming growth factor β can stimulate Smad1 phosphorylation independently of bone morphogenic protein receptors. J. Biol. Chem. 2009, 284, 9755–9763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, J.; Ho, J.N.; Lewis, J.A.; Karim, K.A.; Daniels, R.N.; Gentry, P.R.; Hopkins, C.R.; Lindsley, C.W.; Hong, C.C. In vivo structure–activity relationship study of dorsomorphin analogues identifies selective VEGF and BMP inhibitors. ACS Chem. Biol. 2010, 5, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Cuny, G.D.; Paul, B.Y.; Laha, J.K.; Xing, X.; Liu, J.-F.; Lai, C.S.; Deng, D.Y.; Sachidanandan, C.; Bloch, K.D.; Peterson, R.T. Structure–activity relationship study of bone morphogenetic protein (BMP) signaling inhibitors. Bioorganic Med. Chem. Lett. 2008, 18, 4388–4392. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.-C.; Cheng, C.-J.; Bilen, M.A.; Lu, J.-F.; Satcher, R.L.; Yu-Lee, L.-Y.; Gallick, G.E.; Maity, S.N.; Lin, S.-H. BMP4 promotes prostate tumor growth in bone through osteogenesis. Cancer Res. 2011, 71, 5194–5203. [Google Scholar] [CrossRef] [Green Version]
- Zilberberg, L.; ten Dijke, P.; Sakai, L.Y.; Rifkin, D.B. A rapid and sensitive bioassay to measure bone morphogenetic protein activity. BMC Cell Biol. 2007, 8, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, B.Y.; Deng, D.Y.; Lai, C.S.; Hong, C.C.; Cuny, G.D.; Bouxsein, M.L.; Hong, D.W.; McManus, P.M.; Katagiri, T.; Sachidanandan, C.; et al. BMP type I receptor inhibition reduces heterotopic ossification. Nat. Med. 2008, 14, 1363–1369. [Google Scholar]
- Kaplan, F.S.; Xu, M.; Seemann, P.; Connor, J.M.; Glaser, D.L.; Carroll, L.; Delai, P.; Fastnacht-Urban, E.; Forman, S.J.; Gillessen-Kaesbach, G.; et al. Classic and atypical fibrodysplasia ossificans progressiva (FOP) phenotypes are caused by mutations in the bone morphogenetic protein (BMP) type I receptor ACVR1. Hum. Mutat. 2009, 30, 379–390. [Google Scholar] [CrossRef] [Green Version]
- Pacifici, M.; Shore, E.M. Common mutations in ALK2/ACVR1, a multi-faceted receptor, have roles in distinct pediatric musculoskeletal and neural orphan disorders. Cytokine Growth Factor Rev. 2016, 27, 93–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pignolo, R.J.; Shore, E.M.; Kaplan, F.S. Fibrodysplasia ossificans progressiva: Clinical and genetic aspects. Orphanet J. Rare Dis. 2011, 6, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, J.; Traynor, R.; Sapkota, G.P. The specificities of small molecule inhibitors of the TGFß and BMP pathways. Cell. Signal. 2011, 23, 1831–1842. [Google Scholar] [CrossRef] [Green Version]
- Owens, P.; Pickup, M.W.; Novitskiy, S.V.; Giltnane, J.M.; Gorska, A.E.; Hopkins, C.R.; Hong, C.C.; Moses, H.L. Inhibition of BMP signalling suppresses metastasis in mammary cancer. Oncogene 2015, 34, 2437–2449. [Google Scholar] [CrossRef] [Green Version]
- Augeri, D.J.; Langenfeld, E.; Castle, M.; Gilleran, J.A.; Langenfeld, J. Inhibition of BMP and of TGFβ receptors downregulates expression of XIAP and TAK1 leading to lung cancer cell death. Mol. Cancer 2016, 15, 27. [Google Scholar] [CrossRef] [Green Version]
- Newman, J.H.; Augeri, D.J.; NeMoyer, R.; Malhotra, J.; Langenfeld, E.; Chesson, C.B.; Dobias, N.S.; Lee, M.J.; Tarabichi, S.; Jhawar, S.R.; et al. Novel bone morphogenetic protein receptor inhibitor JL5 suppresses tumor cell survival signaling and induces regression of human lung cancer. Oncogene 2018, 37, 3672–3685. [Google Scholar] [CrossRef]
- Boergermann, J.; Kopf, J.; Yu, P.; Knaus, P. Dorsomorphin and LDN-193189 inhibit BMP-mediated Smad, p38 and Akt signalling in C2C12 cells. Int. J. Biochem. Cell Biol. 2010, 42, 1802–1807. [Google Scholar] [CrossRef]
- Langenfeld, E.; Hong, C.C.; Lanke, G.; Langenfeld, J. Bone morphogenetic protein type I receptor antagonists decrease growth and induce cell death of lung cancer cell lines. PLoS ONE 2013, 8, e61256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.; Wang, X.-L.; Zettervall, S.L.; Cai, Y.; Guzman, R.J. DMH1, a Highly Selective Small Molecule BMP Inhibitor, Suppresses Arterial Medial Calcification. J. Vasc. Surg. 2017, 66, 586. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Overview of BMP signalling cascades. BMP homodimers and heterodimers signal through a heterotetrameric complex of serine/threonine kinase BMP type I and type II receptors. In the induced heterotetrameric complex, BMPR type II phosphorylates the GS-domain of BMPR type I to induce canonical and noncanonical BMP signalling pathways. In the canonical pathway, type I receptors phosphorylate Smads 1, 5, or 8 (R-Smad), which form a heteromeric complex with Smad 4, and then translocate to the nucleus. In the nucleus, this complex forms a complex with Runx2 to regulate osteogenic gene expression. The noncanonical signalling cascade, p38 mitogen-activated protein kinase (MAPK), is initiated by TAK1 phosphorylation, which recruits TAB1 and induces the MKK-P38 MAPK signalling pathway. The phosphorylated form of p38 phosphorylates and activates Runx2, Dlx5, and Sp7 transcription factors in the nucleus to initiate the transcription of osteogenic genes. Furthermore, phosphorylated Runx2 promotes the formation of the Smad–Runx2 complex. “Created with BioRender.com (Accessed on 31 January 2022)”.
Figure 1.
Overview of BMP signalling cascades. BMP homodimers and heterodimers signal through a heterotetrameric complex of serine/threonine kinase BMP type I and type II receptors. In the induced heterotetrameric complex, BMPR type II phosphorylates the GS-domain of BMPR type I to induce canonical and noncanonical BMP signalling pathways. In the canonical pathway, type I receptors phosphorylate Smads 1, 5, or 8 (R-Smad), which form a heteromeric complex with Smad 4, and then translocate to the nucleus. In the nucleus, this complex forms a complex with Runx2 to regulate osteogenic gene expression. The noncanonical signalling cascade, p38 mitogen-activated protein kinase (MAPK), is initiated by TAK1 phosphorylation, which recruits TAB1 and induces the MKK-P38 MAPK signalling pathway. The phosphorylated form of p38 phosphorylates and activates Runx2, Dlx5, and Sp7 transcription factors in the nucleus to initiate the transcription of osteogenic genes. Furthermore, phosphorylated Runx2 promotes the formation of the Smad–Runx2 complex. “Created with BioRender.com (Accessed on 31 January 2022)”.

Figure 2.
Roles of BMPs in skeletal development. BMP ligands are multifunctional growth factors that are important in a range of biological processes but are best recognised for inducing bone formation. MSCs give rise to chondrocytes, which go through a series of differentiation processes. BMPs govern various phases of chondrocyte differentiation by regulating the expression of Sox9 and Runx2. Sox9 promotes the proliferation and differentiation of MSCs into chondrocytes, while Runx2 induces chondrocyte hypertrophy. Moreover, BMPs promotes osteogenesis by enhancing Runx2 and Sp7 transcription factor activity. “Created with BioRender.com (Accessed on 31 January 2022)”.
Figure 2.
Roles of BMPs in skeletal development. BMP ligands are multifunctional growth factors that are important in a range of biological processes but are best recognised for inducing bone formation. MSCs give rise to chondrocytes, which go through a series of differentiation processes. BMPs govern various phases of chondrocyte differentiation by regulating the expression of Sox9 and Runx2. Sox9 promotes the proliferation and differentiation of MSCs into chondrocytes, while Runx2 induces chondrocyte hypertrophy. Moreover, BMPs promotes osteogenesis by enhancing Runx2 and Sp7 transcription factor activity. “Created with BioRender.com (Accessed on 31 January 2022)”.
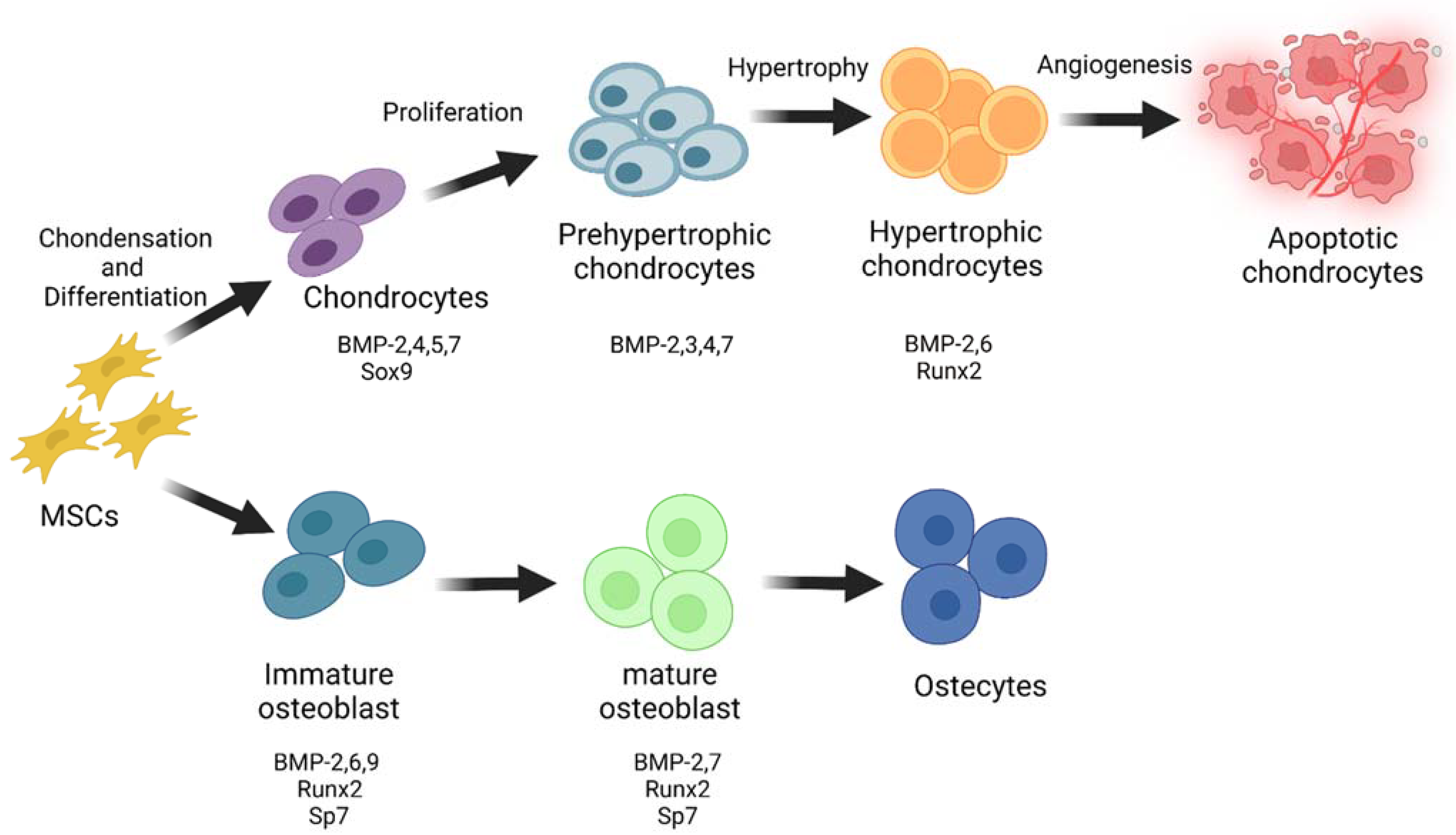
Figure 3.
Interactions between PGs and growth factors in the growth plate during endochondral and perichondral ossifications. Chondrocytes are imbedded in a PG-rich ECM so that growth factors can travel in it to regulate cartilage maturation and send a signal to the perichondrium layer to differentiate undifferentiated cells to the osteoblast. Aggrecan is the most abundant CSPG expressed in cartilage, which is required for creating an appropriate Ihh gradient. Furthermore, Wnts, FGFs, BMPs, and TGF-β are among the cell signalling pathways that can be influenced by CSPGs and HSPGs to regulate cartilage maturation and osteoblast differentiation. “Created with BioRender.com (Accessed on 31 January 2022)”.
Figure 3.
Interactions between PGs and growth factors in the growth plate during endochondral and perichondral ossifications. Chondrocytes are imbedded in a PG-rich ECM so that growth factors can travel in it to regulate cartilage maturation and send a signal to the perichondrium layer to differentiate undifferentiated cells to the osteoblast. Aggrecan is the most abundant CSPG expressed in cartilage, which is required for creating an appropriate Ihh gradient. Furthermore, Wnts, FGFs, BMPs, and TGF-β are among the cell signalling pathways that can be influenced by CSPGs and HSPGs to regulate cartilage maturation and osteoblast differentiation. “Created with BioRender.com (Accessed on 31 January 2022)”.
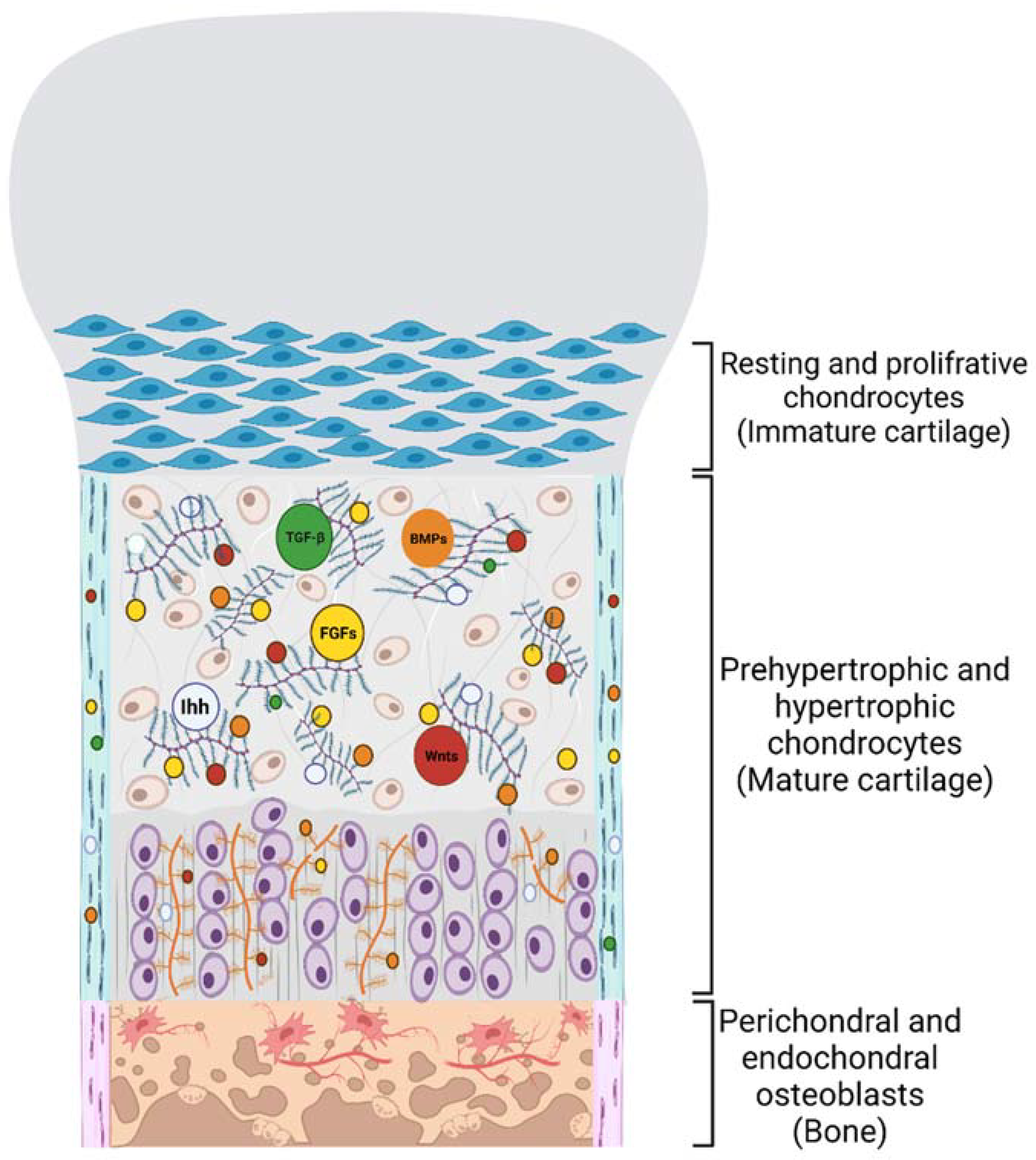

{kind=link}
{kind=link}
{kind=link}
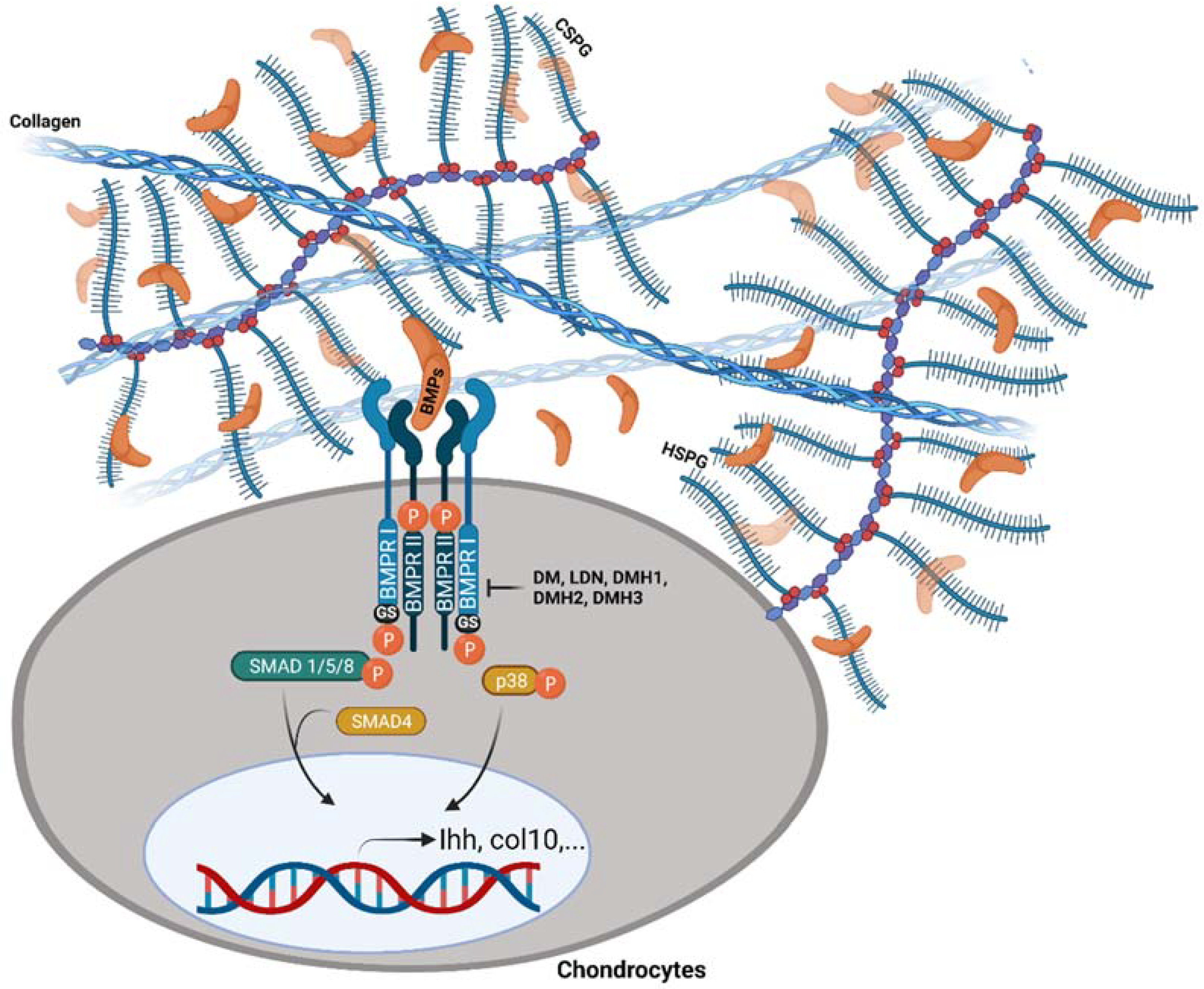{kind=link}
Table 1.
Small-molecule BMP inhibitors: Chemical Names and Structures.
| Compound | Chemical Name | Structure |
|---|---|---|
| Dorsomorphin | 6-[4-[2-(1-Piperidinyl)ethoxy]phenyl]-3-(4-pyridinyl)-pyrazolo[1,5-a]pyrimidine |  |
| LDN-193189 | 4-[6-[4-(1-Piperazinyl)phenyl]pyrazolo[1,5-a]pyrimidin-3-yl]-quinoline dihydrochloride |  |
| DMH1 | 4-[6-(4-Propan-2-yloxyphenyl)pyrazolo[1,5-a]pyrimidin-3-yl]quinoline |  |
| DMH2 | 4-(2-(4-(3-(quinolin-4-yl)pyrazolo[1,5-a]pyrimidin-6-yl)phenoxy)ethyl)morpholine |  |
| JL5 | 4-(3-(4-(3-(quinolin-4-yl)pyrazolo[1,5-a]pyrimidin-6-yl)phenyl)propyl)morpholine |  |
| DMH3 | N,N-dimethyl-3-(4-(3-(quinolin-4-yl)pyrazolo[1,5-a]pyrimidin-6-yl)phenoxy)propan-1-amine |  |
| DMH4 | 4-(2-(4-(3-phenylpyrazolo[1,5-a]pyrimidin-6-yl)phenoxy)ethyl)morpholine |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Koosha, E.; Eames, B.F. Two Modulators of Skeletal Development: BMPs and Proteoglycans. J. Dev. Biol. 2022, 10, 15. https://doi.org/10.3390/jdb10020015
AMA Style
Koosha E, Eames BF. Two Modulators of Skeletal Development: BMPs and Proteoglycans. Journal of Developmental Biology. 2022; 10(2):15. https://doi.org/10.3390/jdb10020015
Chicago/Turabian StyleKoosha, Elham, and B. Frank Eames. 2022. "Two Modulators of Skeletal Development: BMPs and Proteoglycans" Journal of Developmental Biology 10, no. 2: 15. https://doi.org/10.3390/jdb10020015
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.