
A Potential Role for MMPs during the Formation of Non-Neurogenic Placodes


Abstract
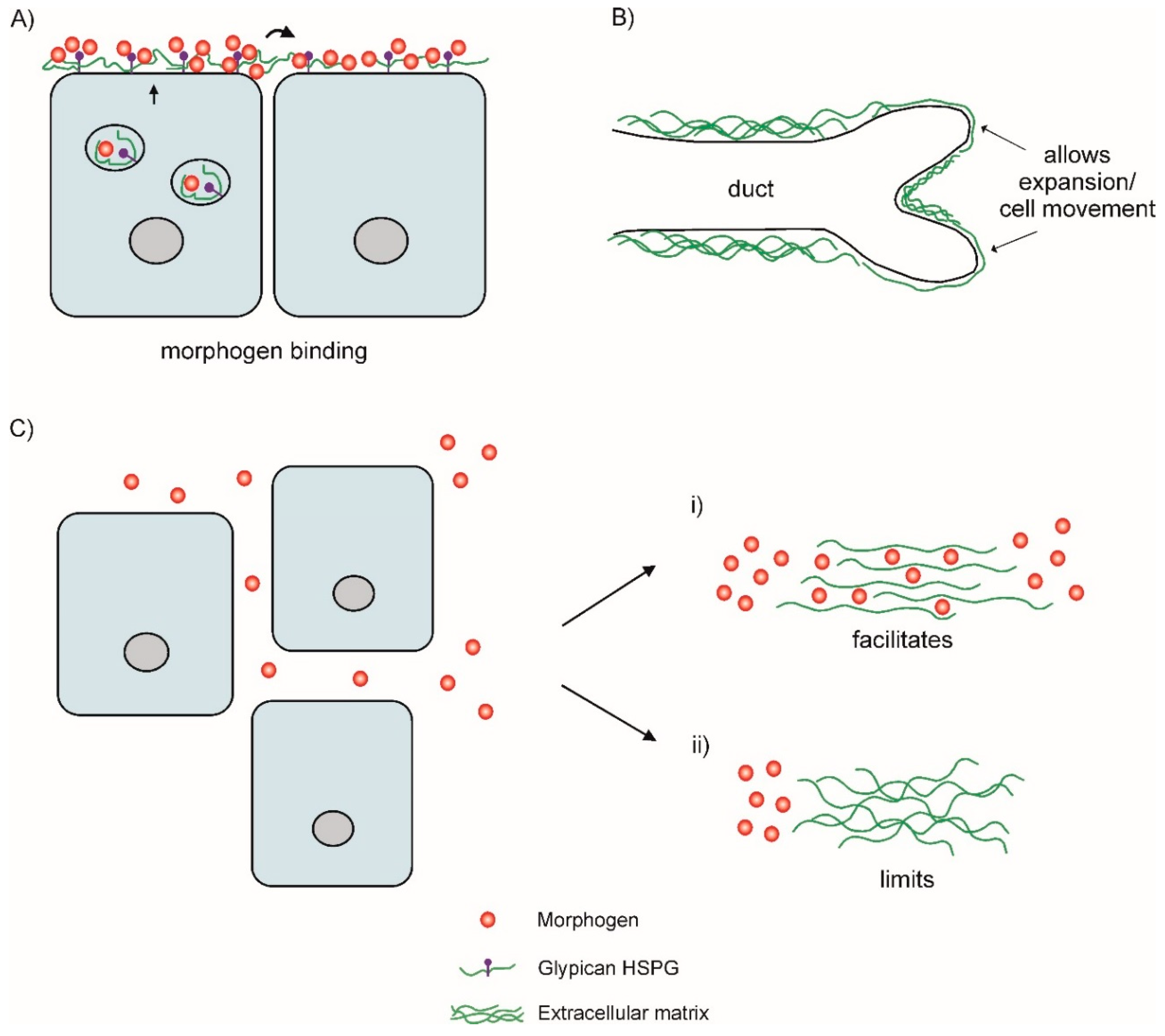
:1. MMPs in the ECM
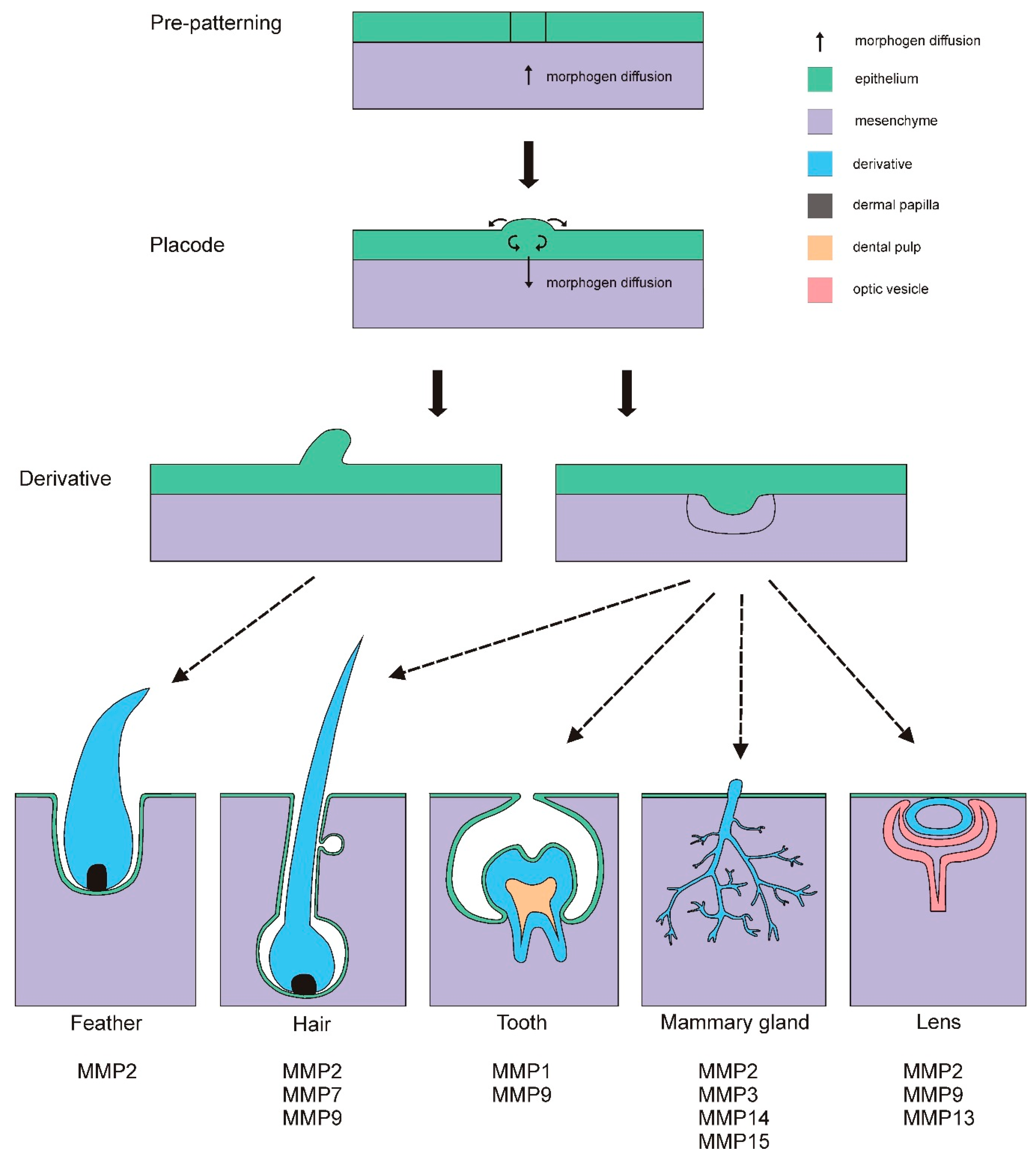
2. Placodes
2.1. Feather and Hair Development
2.2. Mammary Gland Development
2.3. Tooth Development
2.4. Lens Development
2.5. Other Placode Systems
3. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mott, J.D.; Werb, Z. Regulation of matrix biology by matrix metalloproteinases. Curr. Opin. Cell Biol. 2004, 16, 558–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz-Munoz, W.; Khokha, R. The role of tissue inhibitors of metalloproteinases in tumorigenesis and metastasis. Crit. Rev. Clin. Lab. Sci. 2008, 45, 291–338. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O. The extracellular matrix: Not just pretty fibrils. Science 2009, 326, 1216–1219. [Google Scholar] [CrossRef] [PubMed]
- Angerer, L.; Hussain, S.; Wei, Z.; Livingston, B.T. Sea urchin metalloproteases: A genomic survey of the BMP-1/tolloid-like, MMP and ADAM families. Dev. Biol. 2006, 300, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Page-McCaw, A. Remodeling the model organism: Matrix metalloproteinase functions in invertebrates. Semin. Cell Dev. Biol. 2008, 19, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Jackson, B.C.; Nebert, D.W.; Vasiliou, V. Update of human and mouse matrix metalloproteinase families. Hum. Genom. 2010, 4, 194–201. [Google Scholar] [CrossRef]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyatt, R.A.; Keow, J.Y.; Harris, N.D.; Haché, C.A.; Li, D.H.; Crawford, B.D. The zebrafish embryo: A powerful model system for investigating matrix remodeling. Zebrafish 2009, 6, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Das, B.; Mathew, S.; Shi, Y.B. Genome-wide identification of Xenopus matrix metalloproteinases: Conservation and unique duplications in amphibians. BMC Genom. 2009, 10, 81. [Google Scholar] [CrossRef] [PubMed]
- Small, C.D.; Crawford, B.D. Matrix metalloproteinases in neural development: A phylogenetically diverse perspective. Neural Regen. Res. 2016, 11, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozario, T.; DeSimone, D.W. The extracellular matrix in development and morphogenesis: A dynamic view. Dev. Biol. 2010, 341, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Lander, A.D.; Nie, Q.; Wan, F.Y. Do morphogen gradients arise by diffusion? Dev. Cell 2002, 2, 785–796. [Google Scholar] [CrossRef]
- Lin, X. Functions of heparin sulfate proteoglycans in cell signaling during development. Development 2004, 131, 6009–6021. [Google Scholar] [CrossRef] [PubMed]
- Tabata, T.; Takei, Y. Morphogens, their identification and regulation. Development 2004, 131, 703–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, A.J.; Scott, M.P. Incredible journey: How do developmental signals travel through tissue? Genes Dev. 2004, 18, 2985–2997. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Lin, X. Shaping morphogen gradients by proteoglycans. Cold Spring Harb. Perspect. Biol. 2009, 1, a002493. [Google Scholar] [CrossRef] [PubMed]
- Plouhinec, J.L.; Zakin, L.; Moriyama, Y.; De Robertis, E.M. Chordin forms a self-organizing morphogen gradient in the extracellular space between ectoderm and mesoderm in the Xenopus embryo. Proc. Natl. Acad. Sci. USA 2013, 110, 20372–20379. [Google Scholar] [CrossRef] [PubMed]
- Daley, W.P.; Yamada, K.M. ECM-modulated cellular dynamics as a driving force for tissue morphogenesis. Curr. Opin. Genet. Dev. 2013, 23, 408–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harada, M.; Murakami, H.; Okawa, A.; Okimoto, N.; Hiraoka, S.; Nakahara, T.; Akasaka, R.; Shiraishi, Y.; Futatsugi, N.; Mizutani-Koseki, Y.; et al. FGF9 monomer-dimer equilibrium regulates extracellular matrix affinity and tissue diffusion. Nat. Genet. 2009, 41, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, T.; Mohamed, O.A.; Taketo, M.M.; Dufort, D.; Groves, A.K. Wnt signals mediate a fate decision between otic placode and epidermis. Development 2006, 133, 865–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turque, N.; Buttice, G.; Beuscart, A.; Stehelin, D.; Crepieux, P.; Desbiens, X. Hydrocortisone modulates the expression of c-ets-1 and 72 kDa type IV collagenase in chicken dermis during early feather morphogenesis. Int. J. Dev. Biol. 1997, 41, 103–109. [Google Scholar] [PubMed]
- Widelitz, R.B. Wnt signaling in skin organogenesis. Organogenesis 2008, 4, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Sharov, A.A.; Schroeder, M.; Sharova, T.Y.; Mardaryev, A.N.; Peters, E.M.; Tobin, D.J.; Botchkarev, V.A. Matrix metalloproteinase-9 is involved in the regulation of hair canal formation. J. Investig. Dermatol. 2011, 131, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Botchkarev, V.A.; Sharov, A.A. BMP signaling in the control of skin development and hair follicle growth. Differentiation 2004, 72, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.X.; Tuan, T.L.; Wu, P.; Widelitz, R.B.; Chuong, C.M. From buds to follicles: Matrix metalloproteinases in developmental tissue remodeling during feather morphogenesis. Differentiation 2011, 81, 307–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, T.X.; Chuong, C.M. Mechanism of skin morphogenesis. I. Analyses with antibodies to adhesion molecules tenascin, N-CAM, and integrin. Dev. Biol. 1992, 150, 82–98. [Google Scholar] [CrossRef]
- Dhouailly, D.; Sawyer, R.H. Avian scale development. XI. Initial appearance of the dermal defect in scaleless skin. Dev. Biol. 1984, 105, 343–350. [Google Scholar] [CrossRef]
- Mauger, A.; Demarchez, M.; Georges, D.; Herbage, D.; Grimaud, J.A.; Druguet, M.; Hartmann, D.J.; Sengel, P. Répartition du collagène, de la fibronectine et de la laminine au cours de la morphogenèse de la peau et des phanères chez l’embryon de Poulet. CR Acad. Sci. Paris 1982, 294, 475–480. [Google Scholar]
- Mauger, A.; Demarchez, M.; Herbage, D.; Grimaud, J.A.; Druguet, M.; Hartmann, D.J.; Foidart, J.M.; Sengel, P. Immunofluorescent localization of collagen types I, III, IV, fibronectin and laminin during morphogenesis of scales and scaleless skin in the chick embryo. Wilhelm Roux’s Arch. Dev. Biol. 1983, 192, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Cowin, P.; Wysolmerski, J. Molecular mechanisms guiding embryonic mammary gland development. Cold Spring Harb. Perspect. Biol. 2010, 2, a003251. [Google Scholar] [CrossRef] [PubMed]
- Inman, J.L.; Robertson, C.; Mott, J.D.; Bissell, M.J. Mammary gland development: Cell fate specification, stem cells and the microenviroment. Development 2015, 142, 1028–1042. [Google Scholar] [CrossRef] [PubMed]
- Mori, H.; Gjorevski, N.; Inman, J.L.; Bissell, M.J.; Nelson, C.M. Self-organization of engineered epithelial tubules by differential cellular motility. Proc. Natl. Acad. Sci. USA 2009, 106, 14890–14895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, H.; Lo, A.T.; Inman, J.L.; Alcaraz, J.; Ghajar, C.M.; Mott, J.D.; Nelson, C.M.; Chen, C.S.; Zhang, H.; Bascom, J.L.; et al. Transmembrane/cytoplasmic, rather than catalytic, domains of Mmp14 signal to MAPK activation and mammary branching morphogenesis via binding to integrin ß1. Development 2013, 140, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, T.Y.; Rowe, R.G.; Saunders, T.L.; Weiss, S.J. Functional roles of MMP14 and MMP15 in early postnatal mammary gland development. Development 2016, 143, 3956–3968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hens, J.; Dann, P.; Hiremath, M.; Pan, T.C.; Chodosh, L.; Wysolmerski, J. Analysis of gene expression in PTHrP-/- mammary buds supports a role for BMP signaling and MMP2 in the initiation of ductal morphogenesis. Dev. Dyn. 2009, 238, 2713–2724. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, B.S.; Sternlicht, M.D.; Lund, L.R.; Alexander, C.M.; Mott, J.; Bissell, M.J.; Soloway, P.; Itohara, S.; Werb, Z. Site-specific inductive and inhibitory activities of MMP-2 and MMP-3 orchestrate mammary gland branching morphogenesis. J. Cell Biol. 2003, 162, 1123–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sternlicht, M.D.; Lochter, A.; Sympson, C.J.; Huey, B.; Rougier, J.P.; Gray, J.W.; Pinkel, D.; Bissell, M.J.; Werb, Z. The stromal proteinase MMP3/stromelysin-1 promotes mammary carcinogenesis. Cell 1999, 98, 137–146. [Google Scholar] [CrossRef]
- Radisky, D.C.; Levy, D.D.; Littlepage, L.E.; Liu, H.; Nelson, C.M.; Fata, J.E.; Leake, D.; Godden, E.L.; Albertson, D.G.; Nieto, M.A.; et al. Rac1b and reactive oxygen species mediate MMP-3-induced EMT and genomic instability. Nature 2005, 436, 123–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessenbrock, K.; Dijkgraaf, G.J.; Lawson, D.A.; Littlepage, L.E.; Shahi, P.; Pieper, U.; Werb, Z. A role for matrix metalloproteinases in regulating mammary stem cell function via the Wnt signaling pathway. Cell Stem Cell 2013, 13, 300–313. [Google Scholar] [CrossRef] [PubMed]
- Randall, L.E.; Hall, R.C. Temperospatial expression of matrix metalloproteinases 1, 2, 3, and 9 during early tooth development. Connect. Tissue Res. 2002, 43, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Sahlberg, C.; Reponen, P.; Tryggvason, K.; Thesleff, I. Timp-1, -2, and -3 show coexpression with gelatinases A and B during mouse tooth morphogenesis. Eur. J. Oral Sci. 1999, 107, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Sahlberg, C.; Reponen, P.; Tryggvason, K.; Thesleff, I. Association between the expression of murine 72 kDa type IV collagenase by odontoblasts and basement membrane degradation during mouse tooth development. Arch. Oral Biol. 1992, 37, 1021–1030. [Google Scholar] [CrossRef]
- Heikinheimo, K.; Salo, T. Expression of basement membrane type IV collagen and type IV collagenases (MMP-2 and MMP-9) in human fetal teeth. J. Dent. Res. 1995, 74, 1226–1234. [Google Scholar] [CrossRef] [PubMed]
- Kallenbach, E. Electron microscopy of the differentiating rat incisor ameloblast. J. Ultrastruct. Res. 1971, 35, 508–531. [Google Scholar] [CrossRef]
- Feng, J.; McDaniel, J.S.; Chuang, H.H.; Huang, O.; Rakian, A.; Xu, X.; Steffensen, B.; Donly, K.J.; MacDougall, M.; Chen, S. Binding of amelogenin to MMP-9 and their co-expression in developing mouse teeth. J. Mol. Histol. 2012, 43, 473–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DenBesten, P.K.; Heffernan, L.M. Enamel proteases in secretory and maturation enamel of rats ingesting 0 and 100 PPM fluoride in drinking water. Adv. Dent. Res. 1989, 3, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.; Septier, D.; Embery, G.; Goldberg, M. Stromelysin-1 (MMP-3) in forming enamel and predentine in rat incisor-coordinated distribution with proteoglycans suggests a functional role. Histochem. J. 1999, 31, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Papagerakis, P.; Yamakoshi, Y.; Hu, J.C.C.; Bartlett, J.D.; Simmer, J.P. Functions of KLK4 and MMP-20 in dental enamel formation. Biol. Chem. 2008, 389, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Bahuguna, R. Role of matrix metalloproteinases in dental caries, pulp and periapical inflammation: An overview. J. Oral Biol. Craniofac. Res. 2015, 5, 212–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, B.E.; Tian, Y.; Garlisch, A.K.; Carinato, M.E.; Elkins, M.B.; Wolfe, A.D.; Schaefer, J.J.; Perry, K.J.; Henry, J.J. Molecular profiling: Gene expression reveals discrete phases of lens induction and development in Xenopus laevis. Mol. Vis. 2004, 10, 186–198. [Google Scholar] [PubMed]
- Huang, J.; Rajagopal, R.; Liu, Y.; Dattilo, L.K.; Shaham, O.; Ashery-Padan, R.; Beebe, D.C. The mechanism of lens placode formation: A case of matrix-mediated morphogenesis. Dev. Biol. 2011, 355, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Jampel, H.D.; Roche, N.; Stark, M.J.; Roberts, A.B. Transforming growth factor-beta in human aqueous humor. Curr. Eye Res. 1990, 9, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hales, A.M.; Chamberlain, C.G.; McAvoy, J.W. Induction of cataract-like changes in rat lens epithelial explants by transforming growth factor beta. Investig. Ophthalmol. Vis. Sci. 1994, 35, 388–401. [Google Scholar]
- Richiert, D.M.; Ireland, M.E. Matrix metalloproteinase secretion is stimulated by TGF-beta in cultured lens epithelial cells. Curr. Eye Res. 1999, 19, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Tamiya, S.; Wormstone, I.M.; Marcantonio, J.M.; Gavrilovic, J.; Duncan, G. Induction of matrix metalloproteinases 2 and 9 following stress to the lens. Exp. Eye Res. 2000, 71, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Pino, G. The Requirement of Matrix Metalloproteinase 2 and 9 in Transforming Growth Factor Beta Induced Epithelial Mesenchymal Transition of Lens Epithelial Cells. Ph.D. Thesis, McMaster University, Hamilton, ON, Canada, 2011. [Google Scholar]
- Nathu, Z.; Dwivedi, D.J.; Reddan, J.R.; Sheardown, H.; Margetts, P.J.; West-Mays, J.A. Temporal changes in MMP mRNA expression in the lens epithelium during anterior subcapsular cataract formation. Exp. Eye Res. 2009, 88, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, R.W.; Zwaan, J. The matrix of the optic vesicle-presumptive lens interface during induction of the lens in the chicken embryo. J. Embryol. Exp. Morphol. 1975, 33, 1023–1049. [Google Scholar] [PubMed]
- Knosp, W.M.; Knox, S.M.; Hoffman, M.P. Salivary gland organogenesis. Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Cantemir, V. Role of Membrane-Type Matrix Metalloproteinases and Tissue Inhibitor of Metalloproteinase-2 in Neural Crest Cell Migration. Ph.D. Thesis, Creighton University, Omaha, NE, USA, 2005. [Google Scholar]
- Patterson, R.A.; Cavanaugh, A.M.; Cantemir, V.; Brauer, P.R.; Reedy, M.V. MT2-MMP expression during early avian morphogenesis. Anat. Rec. 2013, 296, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Visconti, R.P.; Hilfer, S.R. Perturbation of extracellular matrix prevents association of the otic primordium with the posterior rhombencephalon and inhibits subsequent invagination. Dev. Dyn. 2002, 223, 48–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Ortho, M.P.; Will, H.; Atkinson, S.; Butler, G.; Messent, A.; Gavrilovic, J.; Smith, B.; Timpl, R.; Zardi, L.; Murphy, G. Membrane-type matrix metalloproteinases 1 and 2 exhibit broad-spectrum proteolytic capacities comparable to many matrix metalloproteinases. Eur. J. Biochem. 1997, 250, 751–757. [Google Scholar] [CrossRef] [PubMed]
- Northcutt, R.G. Distribution and innervation of lateral line organs in the axolotl. J. Comp. Neurol. 1992, 325, 95–123. [Google Scholar] [CrossRef] [PubMed]
- Raible, D.W.; Kruse, G.J. Organization of the lateral line system in embryonic zebrafish. J. Comp. Neurol. 2000, 421, 189–198. [Google Scholar] [CrossRef]
- Ledent, V. Postembryonic development of the posterial lateral line in zebrafish. Development 2002, 129, 597–604. [Google Scholar] [PubMed]
- Streit, A. Early development of the cranial sensory nervous system: From a common field to individual placodes. Dev. Biol. 2004, 276, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Schlosser, G. Induction and specification of cranial placodes. Dev. Biol. 2006, 294, 303–351. [Google Scholar] [CrossRef] [PubMed]
- Ma, E.Y.; Raible, D.W. Signaling pathways regulating zebrafish lateral line development. Curr. Biol. 2009, 19, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Vass, S.; Heck, M.M. Perturbation of invadolysin disrupts cell migration in zebrafish (Danio rerio). Exp. Cell Res. 2013, 19, 1198–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, B.D.; Po, M.D.; Saranyan, P.V.; Forsberg, D.; Schulz, R.; Pilgrim, D.B. Mmp25ß facilitates elongation of sensory neurons during zebrafish development. Genesis 2014, 52, 833–848. [Google Scholar] [CrossRef] [PubMed]
- Piccinini, A.M.; Midwood, K.S. Illustrating the interplay between the extracellular matrix and microRNAs. Int. J. Exp. Pathol. 2014, 95, 158–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brabletz, T.; Jung, A.; Dag, S.; Hlubek, F.; Kirchner, T. ß-Catenin regulates the expression of the matrix metalloproteinases-7 in human colorectal cancer. Am. J. Pathol. 1999, 155, 1033–1038. [Google Scholar] [CrossRef]
- Crawford, H.C.; Fingleton, B.M.; Rudolph-Owen, L.A.; Goss, K.J.; Rubinfeld, B.; Polakis, P.; Matrisian, L.M. The metalloproteinase matrilysin is a target of beta-catenin transactivation in intestinal tumors. Oncogene 1999, 18, 2883–2891. [Google Scholar] [CrossRef] [PubMed]
- Garg, P.; Sarma, D.; Jeppsson, S.; Patel, N.R.; Gewirtz, A.T.; Merlin, D.; Sitaraman, S.V. Matrix metalloproteinase-9 functions as a tumor suppressor in colitis-associated cancer. Cancer Res. 2010, 70, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Sonderegger, S.; Haslinger, P.; Sabri, A.; Leisser, C.; Otten, J.V.; Fiala, C.; Knöfler, M. Wingless (Wnt)-3A induces trophoblast migration and matrix metalloproteinase-2 secretion through canonical Wnt signaling and protein kinase B/AKT activation. Endocrinology 2010, 151, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Noramly, S.; Freeman, A.; Morgan, B.A. ß-catenin signaling can initiate feather bud development. Development 1999, 126, 3509–3521. [Google Scholar] [PubMed]
- Andl, T.; Reddy, S.T.; Gaddapara, T.; Millar, S.E. WNT signals are required for the initiation of hair follicle development. Dev. Cell 2002, 2, 643–653. [Google Scholar] [CrossRef]
- Chu, E.Y.; Hens, J.; Andl, T.; Kairo, A.; Yamaguchi, T.P.; Brisken, C.; Glick, A.; Wysolmerski, J.J.; Millar, S.E. Canonical WNT signaling promotes mammary placode development and is essential for initiation of mammary gland morphogenesis. Development 2004, 131, 4819–4829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Thirumangalathu, S.; Gallant, N.M.; Yang, S.H.; Stoick-Cooper, C.L.; Reddy, S.T.; Andl, T.; Taketo, M.M.; Dlugosz, A.A.; Moon, R.T.; et al. Wnt-beta-catenin signaling initiates taste papilla development. Nat. Genet. 2007, 39, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Chu, E.Y.; Watt, B.; Zhang, Y.; Gallant, N.M.; Andl, T.; Yang, S.H.; Lu, M.M.; Piccolo, S.; Schmidt-Ullrich, R.; et al. Wnt/beta-catenin signaling directs multiple stages of tooth morphogenesis. Dev. Biol. 2008, 313, 212–224. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| MMP | Substrate | Placodal Systems |
|---|---|---|
| 1 | Collagens (I, II, III, VII, VIII, X and XI), entactin, proteoglycans, ovostatin, MMP2, MMP9, pro-MMP9, fibronectin, vitronectin, laminin, tenascin, aggrecan, link protein, myelin basic protein, versican | Tooth, feather, otic, lens |
| 2 | Collagens (I, II, III, IV, V, VII, X and XI), gelatin, elastin, fibronectin, vitronectin, laminin, entactin, tenascin, SPARC, aggrecan, link protein, galectin-3, versican, decorin, myelin basic protein | Hair, feather, mammary gland, tooth, lens, otic |
| 3 | Collagens (III, IV, V, VII, IX, X and XI), gelatin, aggrecan, laminin, elastin, casein, osteonectin, fibronectin, ovostatin, entactin, plasminogen, pro-MMP9, vitronectin, tenascin, SPARC, link protein, decorin, myelin basic protein, perlecan, versican, fibulin | Mammary gland, tooth, feather, otic, lens |
| 7 | Collagens (I and IV, gelatin, fibronectin, laminin, elastin, transferrin, casein, vitronectin, SPARC, aggrecan, decorin, versican, fibulin, myelin basic protein | Hair, feather, lens |
| 8 | Collagens (I, II and III), fibronectin, proteoglycans, aggrecan | feather, lens |
| 9 | Collagens (IV, V, VII, X and XIV), gelatin, elastin, fibronectin, vitronectin, laminin, SPARC, aggrecan, link protein, galectin-3, versican, decorin, myelin basic protein | Hair, feather, tooth, lens, otic |
| 10 | Collagens (III, IV, V), gelatin, casein, elastin, fibronectin, aggrecan, link protein | feather, lens |
| 11 | Collagen IV, laminin, elastin, fibronectin, casein, proteoglycans | Feather, otic, lens |
| 12 | Collagen IV, elastin, gelatin, casein, fibronectin, vitronectin, laminin, entactin, fibrinogen | Feather, otic, lens |
| 13 | Collagens (I, II, III, IV, VI, IX, X and XIV), large tenascin-C, plasminogen, aggrecan, fibronectin, SPARC, gelatin, perlecan MMP9 | Lens, feather |
| 14 | Collagens (I, II and III), gelatin, fibronectin, laminin, vitronectin, entactin, pro-MMP2, aggrecan, perlecan, | Mammary gland, salivary gland, feather, otic, lens |
| 15 | Fibronectin, gelatin, vitronectin, entactin, laminin, pro-MMP2, tenascin, perlecan | Mammary gland, salivary gland, otic, feather, lens |
| 16 | Collagen III, gelatin, casein, fibronectin, pro-MMP2, laminin, vitronectin | Feather, otic, lens |
| 17 | Gelatin, fibrinogen, pro-MMP2 | |
| 18 | Collagen I | |
| 19 | Collagens (I and IV), gelatin, fibronectin, laminin, entactin, large tenascin-C, fibronectin | Feather, otic, lens |
| 20 | Amelogenin, aggrecan | |
| 21 | Gelatin, casein | |
| 23 (A, B) | Gelatin | |
| 24 | Fibronectin, pro-MMP2, proteoglycans, gelatin | Feather, lens |
| 25 | Collagen IV, gelatin, fibronectin, pro-MMP2, pro-MMPp | Feather, lens |
| 26 | Collagen IV, fibrinogen, fibronectin, gelatin, pro-MMP9, vitronectin | feather |
| 27 | Gelatin, casein | |
| 28 | Casein |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drake, P.M.; Franz-Odendaal, T.A. A Potential Role for MMPs during the Formation of Non-Neurogenic Placodes. J. Dev. Biol. 2018, 6, 20. https://doi.org/10.3390/jdb6030020
Drake PM, Franz-Odendaal TA. A Potential Role for MMPs during the Formation of Non-Neurogenic Placodes. Journal of Developmental Biology. 2018; 6(3):20. https://doi.org/10.3390/jdb6030020
Chicago/Turabian StyleDrake, Paige M., and Tamara A. Franz-Odendaal. 2018. "A Potential Role for MMPs during the Formation of Non-Neurogenic Placodes" Journal of Developmental Biology 6, no. 3: 20. https://doi.org/10.3390/jdb6030020
APA StyleDrake, P. M., & Franz-Odendaal, T. A. (2018). A Potential Role for MMPs during the Formation of Non-Neurogenic Placodes. Journal of Developmental Biology, 6(3), 20. https://doi.org/10.3390/jdb6030020