
Drosophila Corazonin Neurons as a Hub for Regulating Growth, Stress Responses, Ethanol-Related Behaviors, Copulation Persistence and Sexually Dimorphic Reward Pathways



{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Developmental Processes Involving Crz-Neuron Function Are Required for the Regulation of Diverse Physiological Mechanisms
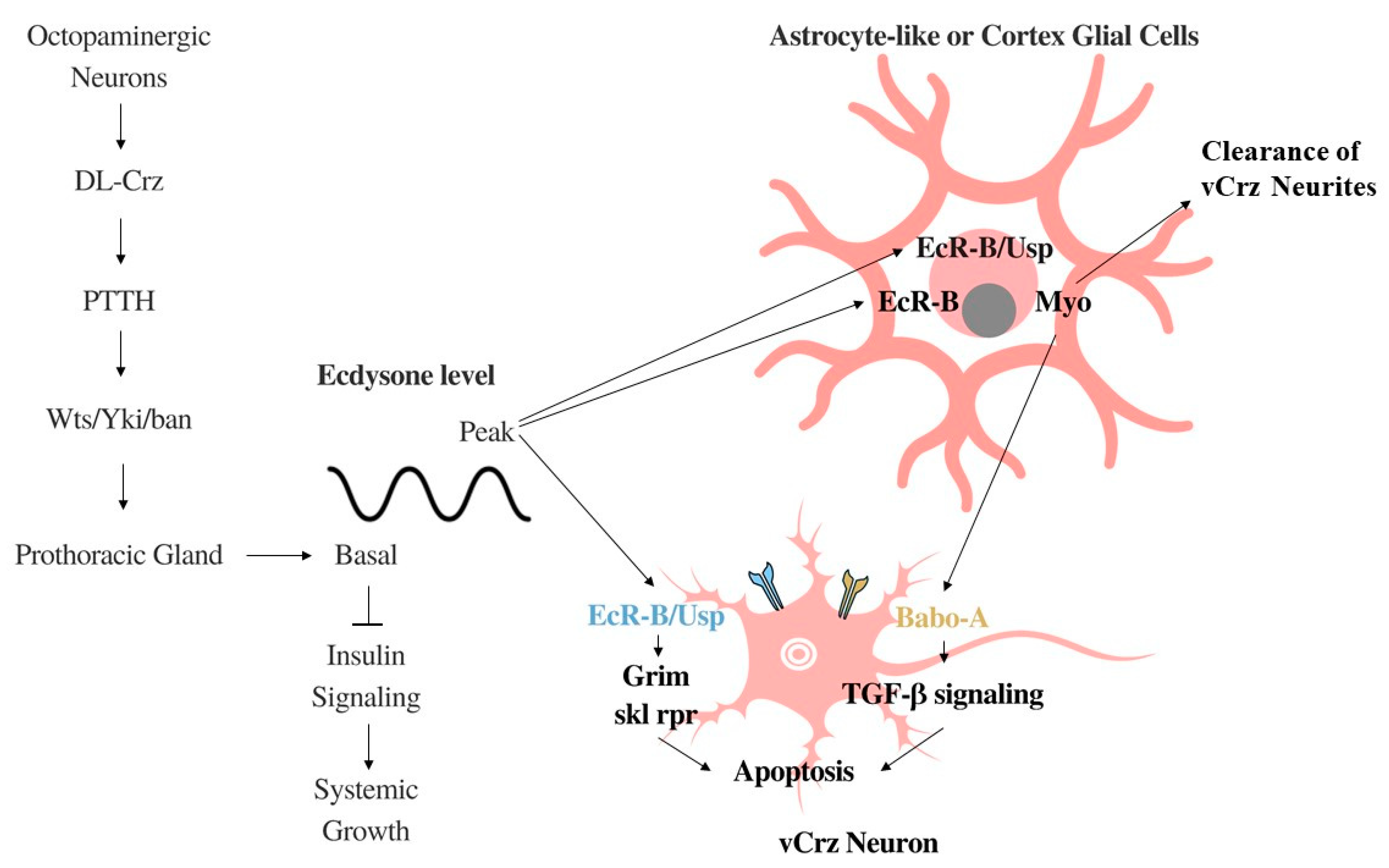
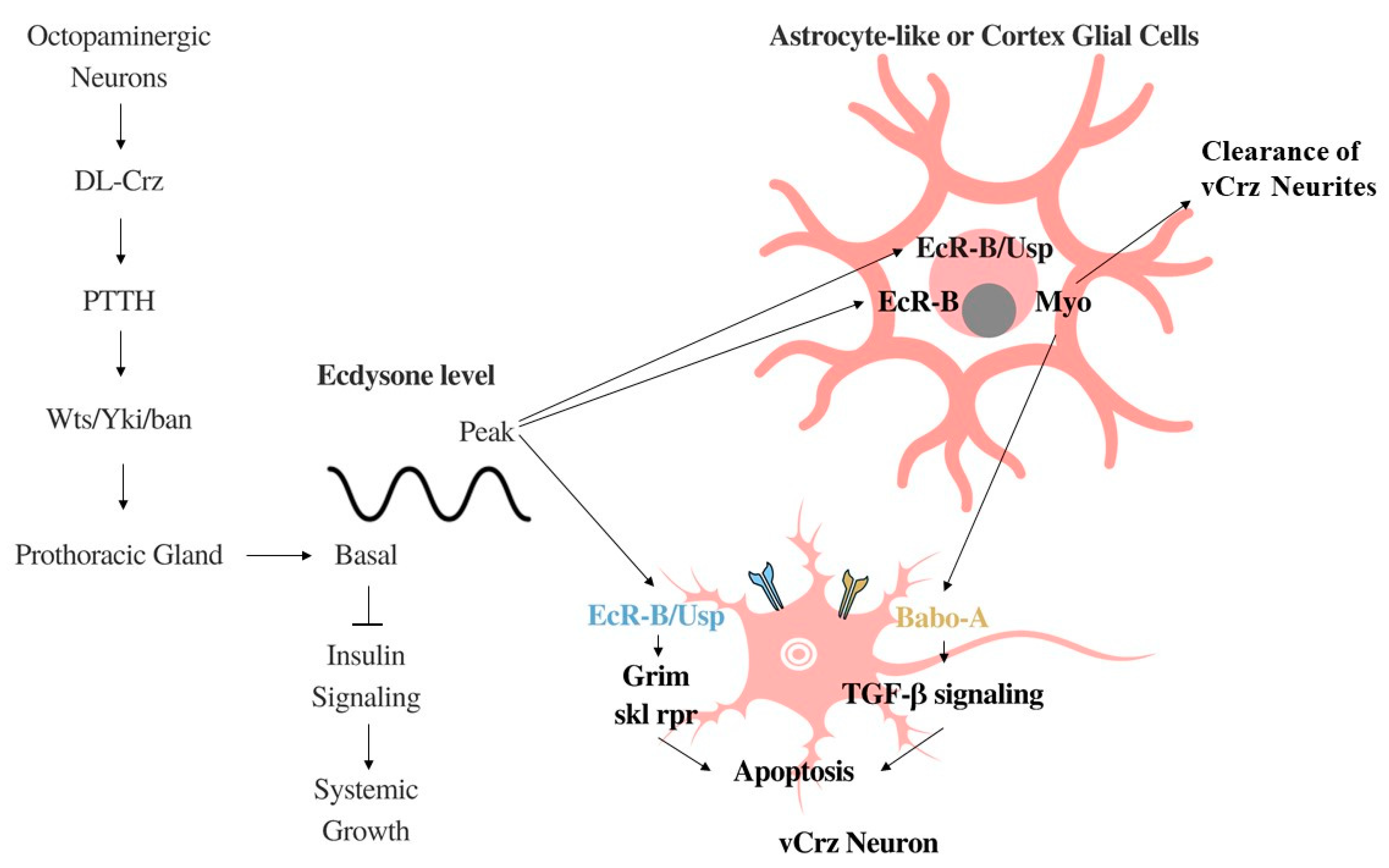
2.1. Corazonin Regulates Systemic Growth, Feeding Behavior, Stress, and Homeostasis
2.2. Systemic Growth Inhibition by DL-Crz Modulation of Ecdysone
2.3. Ecdysone-Driven Apoptosis of vCrz Neurons during Metamorphosis
3. Role of Crz Neurons in Ethanol-Related Behaviors
3.1. Developmental Transcription Regulation and Neuronal Activity in Crz Neurons Modulate Sensitivity to Ethanol Sedation
3.2. Ethanol Recovery and Consumption Are Regulated by Crz Neuron Function
4. Crz Neuron Activity Modulates Mating-Related Features Such as Ejaculation, Copulation Duration, and Reward-Behaviors
4.1. Crz Functions as a Hub Regulating Ejaculation within the VNC
4.2. Different Degrees of Conservation in Mechanisms and Molecules Regulating Ejaculation That Relate to Crz-Dependent Signaling
4.3. Crz-Independent Function of abgCrz Neurons
4.4. Crz Function as a Timer to Regulate Copulation Duration and Animal Persistence
4.5. Activation of Crz Neurons Is Pleasurable
5. Future Directions
Possible Use of Fictive Mating to Study Sex Addiction and Feeding Disorders in Drosophila
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Veenstra, J.A. Isolation and structure of corazonin, a cardioactive peptide from the American cockroach. FEBS Lett. 1989, 250, 231–234. [Google Scholar] [CrossRef] [Green Version]
- Jékely, G. Global view of the evolution and diversity of metazoan neuropeptide signaling. Proc. Natl. Acad. Sci. USA 2013, 110, 8702–8707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boerjan, B.; Verleyen, P.; Huybrechts, J.; Schoofs, L.; De Loof, A. In search for a common denominator for the diverse functions of arthropod corazonin: A role in the physiology of stress? Gen. Comp. Endocrinol. 2010, 166, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Zandawala, M.; Tian, S.; Elphick, M.R. The evolution and nomenclature of GnRH-type and corazonin-type neuropeptide signaling systems. Gen. Comp. Endocrinol. 2018, 264, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Semmens, D.C.; Elphick, M.R. The evolution of neuropeptide signalling: Insights from echinoderms. Briefings Funct. Genom. 2017, 16, 288–298. [Google Scholar] [CrossRef] [Green Version]
- Tayler, T.D.; Pacheco, D.A.; Hergarden, A.C.; Murthy, M.; Anderson, D.J. A neuropeptide circuit that coordinates sperm transfer and copulation duration in Drosophila. Proc. Natl. Acad. Sci. USA 2012, 109, 20697–20702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClure, K.D.; Heberlein, U. A Small Group of Neurosecretory Cells Expressing the Transcriptional Regulator apontic and the Neuropeptide corazonin Mediate Ethanol Sedation in Drosophila. J. Neurosci. 2013, 33, 4044–4054. [Google Scholar] [CrossRef] [PubMed]
- Zer-Krispil, S.; Zak, H.; Shao, L.; Ben-Shaanan, S.; Tordjman, L.; Bentzur, A.; Shmueli, A.; Shohat-Ophir, G. Ejaculation Induced by the Activation of Crz Neurons Is Rewarding to Drosophila Males. Curr. Biol. 2018, 28, 1445–1452.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornquist, S.C.; Langer, K.; Zhang, S.X.; Rogulja, D.; Crickmore, M.A. CaMKII Measures the Passage of Time to Coordinate Behavior and Motivational State. Neuron 2020, 105, 334–345.e9. [Google Scholar] [CrossRef]
- Hartenstein, V. The neuroendocrine system of invertebrates: A developmental and evolutionary perspective. J. Endocrinol. 2006, 190, 555–570. [Google Scholar] [CrossRef] [Green Version]
- Nässel, D.R.; Zandawala, M. Hormonal axes in Drosophila: Regulation of hormone release and multiplicity of actions. Cell Tissue Res. 2020, 382, 233–266. [Google Scholar] [CrossRef]
- Lubawy, J.; Urbański, A.; Colinet, H.; Pflüger, H.-J.; Marciniak, P. Role of the Insect Neuroendocrine System in the Response to Cold Stress. Front. Physiol. 2020, 11, 376. [Google Scholar] [CrossRef]
- Hückesfeld, S.; Schlegel, P.; Miroschnikow, A.; Schoofs, A.; Zinke, I.; Haubrich, A.N.; Schneider-Mizell, C.M.; Truman, J.W.; Fetter, R.D.; Cardona, A.; et al. Unveiling the sensory and interneuronal pathways of the neuroendocrine connectome in Drosophila. eLife 2021, 10, 65745. [Google Scholar] [CrossRef]
- Pandey, U.B.; Nichols, C. Human Disease Models in Drosophila melanogaster and the Role of the Fly in Therapeutic Drug Discovery. Pharmacol. Rev. 2011, 63, 411–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.; Kim, K.-M.; Kikuno, K.; Wang, Z.; Choi, Y.-J.; Park, J.H. Developmental regulation and functions of the expression of the neuropeptide corazonin in Drosophila melanogaster. Cell Tissue Res. 2007, 331, 659–673. [Google Scholar] [CrossRef]
- Sha, K.; Choi, S.-H.; Im, J.; Lee, G.G.; Loeffler, F.; Park, J.H. Regulation of Ethanol-Related Behavior and Ethanol Metabolism by the Corazonin Neurons and Corazonin Receptor in Drosophila melanogaster. PLoS ONE 2014, 9, e87062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imura, E.; Shimada-Niwa, Y.; Nishimura, T.; Hückesfeld, S.; Schlegel, P.; Ohhara, Y.; Kondo, S.; Tanimoto, H.; Cardona, A.; Pankratz, M.J.; et al. The Corazonin-PTTH Neuronal Axis Controls Systemic Body Growth by Regulating Basal Ecdysteroid Biosynthesis in Drosophila melanogaster. Curr. Biol. 2020, 30, 2156–2165.e5. [Google Scholar] [CrossRef] [PubMed]
- Zandawala, M.; Nguyen, T.; Segura, M.B.; Johard, H.A.D.; Amcoff, M.; Wegener, C.; Paluzzi, J.-P.; Nässel, D.R. A neuroendocrine pathway modulating osmotic stress in Drosophila. PLoS Genet. 2021, 17, e1009425. [Google Scholar] [CrossRef] [PubMed]
- Veenstra, J.A. Does corazonin signal nutritional stress in insects? Insect Biochem. Mol. Biol. 2009, 39, 755–762. [Google Scholar] [CrossRef]
- Kapan, N.; Lushchak, O.V.; Luo, J.; Nässel, D.R. Identified peptidergic neurons in the Drosophila brain regulate insulin-producing cells, stress responses and metabolism by coexpressed short neuropeptide F and corazonin. Cell. Mol. Life Sci. 2012, 69, 4051–4066. [Google Scholar] [CrossRef] [PubMed]
- Nässel, D.R.; Enell, L.E.; Santos, J.G.; Wegener, C.; Johard, H.A. A large population of diverse neurons in the Drosophilacentral nervous system expresses short neuropeptide F, suggesting multiple distributed peptide functions. BMC Neurosci. 2008, 9, 90. [Google Scholar] [CrossRef]
- Megha; Wegener, C.; Hasan, G. ER-Ca2+ sensor STIM regulates neuropeptides required for development under nutrient restriction in Drosophila. PLoS ONE 2019, 14, e0219719. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, T.; Amrein, H. Diverse roles for the Drosophila fructose sensor Gr43a. Fly (Austin). Landes Biosci. 2014, 8, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Mishra, D.; Miyamoto, T.; Rezenom, Y.H.; Broussard, A.; Yavuz, A.; Slone, J.; Russell, D.H.; Amrein, H. The Molecular Basis of Sugar Sensing in Drosophila Larvae. Curr. Biol. 2013, 23, 1466–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, E.C.; Shafer, O.T.; Trigg, J.S.; Park, J.; Schooley, D.A.; Dow, J.; Taghert, P.H. A novel diuretic hormone receptor in Drosophila: Evidence for conservation of CGRP signaling. J. Exp. Biol. 2005, 208, 1239–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubrak, O.I.; Lushchak, O.V.; Zandawala, M.; Nässel, D.R. Systemic corazonin signalling modulates stress responses and metabolism in Drosophila. Open Biol. 2016, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.-S.; You, K.-H.; Choo, J.-K.; Han, Y.-M.; Yu, K. Drosophila Short Neuropeptide F Regulates Food Intake and Body Size. J. Biol. Chem. 2004, 279, 50781–50789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rulifson, E.J.; Kim, S.K.; Nusse, R. Ablation of insulin-producing neurons in flies: Growth and diabetic phenotypes. Science 2002, 296, 1118–1120. [Google Scholar] [CrossRef] [PubMed]
- Brogiolo, W.; Stocker, H.; Ikeya, T.; Rintelen, F.; Fernandez, R.; Hafen, E. An evolutionarily conserved function of the Drosophila insulin receptor and insulin-like peptides in growth control. Curr. Biol. 2001, 11, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Hergarden, A.C.; Tayler, T.D.; Anderson, D.J. Allatostatin-A neurons inhibit feeding behavior in adult Drosophila. Proc. Natl. Acad. Sci. USA 2012, 109, 3967–3972. [Google Scholar] [CrossRef] [Green Version]
- Tatar, M.; Post, S.; Yu, K. Nutrient control of Drosophila longevity. Trends Endocrinol. Metab. 2014, 25, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Tatar, M. Aging Regulated Through a Stability Model of Insulin/Insulin Growth Factor Receptor Function. Front. Endocrinol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Bretz, C.A.; Hawksworth, S.A.; Hirsh, J.; Johnson, E.C. Corazonin Neurons Function in Sexually Dimorphic Circuitry That Shape Behavioral Responses to Stress in Drosophila. PLoS ONE 2010, 5, e9141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, R.; Palmer, M.; Koski, H.; Curtis-Joseph, N.; Tatar, M. Aging modulated by the Drosophila insulin receptor through distinct structure-defined mechanisms. Genetics 2021, 217. [Google Scholar] [CrossRef] [PubMed]
- Root, C.M.; Ko, K.I.; Jafari, A.; Wang, J.W. Presynaptic Facilitation by Neuropeptide Signaling Mediates Odor-Driven Food Search. Cell 2011, 145, 133–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, W.T.; Gonzalez, C.R.; Fernandez, C.; Ramasamy, L.; Tabachnik, T.; Du, R.R.; Felsen, P.D.; Maire, M.R.; Perona, P.; Anderson, D.J. Behavioral Responses to a Repetitive Visual Threat Stimulus Express a Persistent State of Defensive Arousal in Drosophila. Curr. Biol. 2015, 25, 1401–1415. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, L.I.; Warren, J.T. A Molecular Genetic Approach to the Biosynthesis of the Insect Steroid Molting Hormone. Vitam. Horm. 2005, 73, 31–57. [Google Scholar] [CrossRef]
- Yamanaka, N.; Rewitz, K.F.; O’Connor, M.B. Ecdysone Control of Developmental Transitions: Lessons fromDrosophilaResearch. Annu. Rev. Èntomol. 2013, 58, 497–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombani, J.; Bianchini, L.; Layalle, S.; Pondeville, E.; Dauphin-Villemant, C.; Antoniewski, C.; Carré, C.; Noselli, S.; Léopold, P. Antagonistic actions of ecdysone and insulins determine final size in Drosophila. Science 2005, 310, 667–670. [Google Scholar] [CrossRef] [PubMed]
- McBrayer, Z.; Ono, H.; Shimell, M.; Parvy, J.-P.; Beckstead, R.B.; Warren, J.T.; Thummel, C.S.; Dauphin-Villemant, C.; Gilbert, L.I.; O’Connor, M.B. Prothoracicotropic Hormone Regulates Developmental Timing and Body Size in Drosophila. Dev. Cell 2007, 13, 857–871. [Google Scholar] [CrossRef] [Green Version]
- Boulan, L.; Martín, D.; Milan, M. bantam miRNA Promotes Systemic Growth by Connecting Insulin Signaling and Ecdysone Production. Curr. Biol. 2013, 23, 473–478. [Google Scholar] [CrossRef] [Green Version]
- Boulan, L.; Milan, M.; Léopold, P. The Systemic Control of Growth. Cold Spring Harb. Perspect. Biol. 2015, 7, a019117. [Google Scholar] [CrossRef]
- Moeller, M.E.; Nagy, S.; Gerlach, S.U.; Soegaard, K.C.; Danielsen, E.T.; Texada, M.J.; Rewitz, K.F. Warts Signaling Controls Organ and Body Growth through Regulation of Ecdysone. Curr. Biol. 2017, 27, 1652–1659.e4. [Google Scholar] [CrossRef] [Green Version]
- Seidman, D.S.; Laor, A.; Gale, R.; Stevenson, D.K.; Danon, Y.L. A Longitudinal Study of Birth Weight and Being Overweight in Late Adolescence. Arch. Pediatr. Adolesc. Med. 1991, 145, 779–781. [Google Scholar] [CrossRef]
- Ravelli, G.-P.; Stein, Z.A.; Susser, M.W. Obesity in Young Men after Famine Exposure in Utero and Early Infancy. N. Engl. J. Med. 1976, 295, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Strauss, R.S.; Dietz, W.H. Effects of intrauterine growth retardation in premature infants on early childhood growth. J. Pediatr. 1997, 130, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Dietz, W.H. Periods of Risk in Childhood for the Development of Adult Obesity—What Do We Need to Learn? J. Nutr. 1997, 127, 1884S–1886S. [Google Scholar] [CrossRef] [PubMed]
- Cowan, W.M.; Fawcett, J.; O’Leary, D.D.; Stanfield, B.B. Regressive events in neurogenesis. Science 1984, 225, 1258–1265. [Google Scholar] [CrossRef] [PubMed]
- Piochon, C.; Kano, M.; Hansel, C.P.C. LTD-like molecular pathways in developmental synaptic pruning. Nat. Neurosci. 2016, 19, 1299–1310. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.; Truman, J. Remodeling dendrites during insect metamorphosis. J. Neurobiol. 2005, 64, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Vonhoff, F.; Keshishian, H. Activity-Dependent Synaptic Refinement: New Insights from Drosophila. Front. Syst. Neurosci. 2017, 11. [Google Scholar] [CrossRef] [Green Version]
- Schuldiner, O.; Yaron, A. Mechanisms of developmental neurite pruning. Cell. Mol. Life Sci. 2014, 72, 101–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Low, L.K.; Cheng, H.-J. Axon pruning: An essential step underlying the developmental plasticity of neuronal connections. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1531–1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.-J.; Lee, G.; Park, J. Programmed cell death mechanisms of identifiable peptidergic neurons in Drosophila melanogaster. Development 2006, 133, 2223–2232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Lee, G.; Vuong, R.; Park, J.H. Two-factor specification of apoptosis: TGF-β signaling acts cooperatively with ecdysone signaling to induce cell- and stage-specific apoptosis of larval neurons during metamorphosis in Drosophila melanogaster. Apoptosis 2019, 24, 972–989. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Urban, J.; Technau, G.M. Distribution, classification, and development ofDrosophila glial cells in the late embryonic and early larval ventral nerve cord. Dev. Genes Evol. 1995, 204, 284–307. [Google Scholar] [CrossRef]
- Lee, G.; Sehgal, R.; Wang, Z.; Park, J.H. Ultraspiracle-independent anti-apoptotic function of ecdysone receptors is required for the survival of larval peptidergic neurons via suppression of grim expression in Drosophila melanogaster. Apoptosis 2019, 24, 256–268. [Google Scholar] [CrossRef]
- Lee, G.; Sehgal, R.; Wang, Z.; Nair, S.; Kikuno, K.; Chen, C.-H.; Hay, B.; Park, J.H. Essential role of grim-led programmed cell death for the establishment of corazonin-producing peptidergic nervous system during embryogenesis and metamorphosis in Drosophila melanogaster. Biol. Open 2013, 2, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Tasdemir-Yilmaz, O.E.; Freeman, M.R. Astrocytes engage unique molecular programs to engulf pruned neuronal debris from distinct subsets of neurons. Genes Dev. 2013, 28, 20–33. [Google Scholar] [CrossRef] [Green Version]
- Diamanti-Kandarakis, E.; Bourguignon, J.-P.; Giudice, L.C.; Hauser, R.; Prins, G.S.; Soto, A.M.; Zoeller, R.T.; Gore, A.C. Endocrine-Disrupting Chemicals: An Endocrine Society Scientific Statement. Endocr. Rev. 2009, 30, 293–342. [Google Scholar] [CrossRef]
- Kajta, M.; Wójtowicz, A.K. Impact of endocrine-disrupting chemicals on neural development and the onset of neurological disorders. Pharmacol. Rep. 2013, 65, 1632–1639. [Google Scholar] [CrossRef]
- Van der Linde, K.; Lyons, L.C. Circadian modulation of acute alcohol sensitivity but not acute tolerance in Drosophila. Chronobiol. Int. 2011, 28, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Shohat-Ophir, G.; Kaun, K.R.; Azanchi, R.; Mohammed, H.; Heberlein, U. Sexual Deprivation Increases Ethanol Intake in Drosophila. Science 2012, 335, 1351–1355. [Google Scholar] [CrossRef] [Green Version]
- Nässel, D.R.; Wegener, C. A comparative review of short and long neuropeptide F signaling in invertebrates: Any similarities to vertebrate neuropeptide Y signaling? Peptides 2011, 32, 1335–1355. [Google Scholar] [CrossRef] [PubMed]
- Kacsoh, B.Z.; Lynch, Z.; Mortimer, N.; Schlenke, T.A. Fruit Flies Medicate Offspring After Seeing Parasites. Science 2013, 339, 947–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadanandappa, M.K.; Sathyanarayana, S.H.; Kondo, S.; Bosco, G. Neuropeptide F signaling regulates parasitoid-specific germline development and egg-laying in Drosophila. PLoS Genet. 2021, 17, e1009456. [Google Scholar] [CrossRef]
- Wen, T.; Parrish, C.A.; Xu, D.; Wu, Q.; Shen, P. Drosophila neuropeptide F and its receptor, NPFR1, define a signaling pathway that acutely modulates alcohol sensitivity. Proc. Natl. Acad. Sci. USA 2005, 102, 2141–2146. [Google Scholar] [CrossRef] [Green Version]
- Thiele, T.E.; Marsh, D.J.; Marie, L.S.; Bernstein, I.L.; Palmiter, R.D. Ethanol consumption and resistance are inversely related to neuropeptide Y levels. Nat. Cell Biol. 1998, 396, 366–369. [Google Scholar] [CrossRef]
- Davies, A.G.; Pierce-Shimomura, J.T.; Kim, H.; VanHoven, M.; Thiele, T.; Bonci, A.; Bargmann, C.; McIntire, S.L. A Central Role of the BK Potassium Channel in Behavioral Responses to Ethanol in C. elegans. Cell 2003, 115, 655–666. [Google Scholar] [CrossRef] [Green Version]
- Sapiro, A.L.; Shmueli, A.; Henry, G.L.; Li, Q.; Shalit, T.; Yaron, O.; Shohat-Ophir, G. Illuminating spatial A-to-I RNA editing signatures within the Drosophila brain. Proc. Natl. Acad. Sci. USA 2019, 116, 2318–2327. [Google Scholar] [CrossRef] [Green Version]
- Ryvkin, J.; Bentzur, A.; Shmueli, A.; Tannenbaum, M.; Shallom, O.; Dokarker, S.; Benichou, J.I.C.; Levi, M.; Shohat-Ophir, G. Transcriptome Analysis of NPFR Neurons Reveals a Connection Between Proteome Diversity and Social Behavior. Front. Behav. Neurosci. 2021, 15. [Google Scholar] [CrossRef] [PubMed]
- Ferveur, J.-F.; Greenspan, R.J. Courtship behavior of brain mosaics in Drosophila. J. Neurogenet. 1998, 12, 205–226. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.C. Control of male reproductive behavior by the central nervous system of Drosophila: Dissection of a courtship pathway by genetic mosaics. Genetics 1979, 92, 437–457. [Google Scholar] [CrossRef]
- Pavlou, H.J.; Lin, A.C.; Neville, M.C.; Nojima, T.; Diao, F.; Chen, B.E.; White, B.H.; Goodwin, S.F. Neural circuitry coordinating male copulation. eLife 2016, 5, e20713. [Google Scholar] [CrossRef]
- Jois, S.; Chan, Y.B.; Fernández, M.P.; Leung, A.K.-W. Characterization of the Sexually Dimorphic fruitless Neurons That Regulate Copulation Duration. Front. Physiol. 2018, 9, 780. [Google Scholar] [CrossRef] [Green Version]
- Hou, Q.-L.; Chen, E.-H.; Jiang, H.-B.; Yu, S.-F.; Yang, P.-J.; Liu, X.-Q.; Park, Y.; Wang, J.-J.; Smagghe, G. Corazonin Signaling Is Required in the Male for Sperm Transfer in the Oriental Fruit Fly Bactrocera dorsalis. Front. Physiol. 2018, 9, 660. [Google Scholar] [CrossRef] [PubMed]
- Gillman, N.; Gillman, M. Premature Ejaculation: Aetiology and Treatment Strategies. Med Sci. 2019, 7, 102. [Google Scholar] [CrossRef] [Green Version]
- Giuliano, F.; Clément, P. Serotonin and Premature Ejaculation: From Physiology to Patient Management. Eur. Urol. 2006, 50, 454–466. [Google Scholar] [CrossRef]
- McMahon, C.G. Emerging and investigational drugs for premature ejaculation. Transl. Androl. Urol. 2016, 5, 487–501. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Zhang, X.; Jiang, T.; Huang, Y.; Cheng, P.; Tang, D.; Gao, J.; Du, J. Association Between Lifelong Premature Ejaculation and Polymorphism of Tryptophan Hydroxylase 2 Gene in the Han Population. Sex. Med. 2020, 8, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Beaulieu, J.-M.; Sotnikova, T.D.; Gainetdinov, R.; Caron, M.G. Tryptophan Hydroxylase-2 Controls Brain Serotonin Synthesis. Science 2004, 305, 217. [Google Scholar] [CrossRef] [PubMed]
- Ozbek, E.; Tasci, A.I.; Tugcu, V.; Ilbey, Y.O.; Simsek, A.; Ozcan, L.; Polat, E.C.; Koksal, V. Possible association of the 5-HTTLPR serotonin transporter promoter gene polymorphism with premature ejaculation in a Turkish population. Asian J. Androl. 2009, 11, 351–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, N.; Huang, Y.; Zhao, H.; Li, G. Association between the serotonin transporter linked polymorphic region and lifelong premature ejaculation: An updated meta-analysis of case-control studies. Medicine 2020, 99, e22169. [Google Scholar] [CrossRef]
- Chan, J.S.; Snoeren, E.; Cuppen, E.; Waldinger, M.D.; Olivier, B.; Oosting, R.S. The Serotonin Transporter Plays an Important Role in Male Sexual Behavior: A Study in Serotonin Transporter Knockout Rats. J. Sex. Med. 2011, 8, 97–108. [Google Scholar] [CrossRef]
- Geng, H.; Peng, D.; Huang, Y.; Tang, D.; Gao, J.; Zhang, Y.; Zhang, X. Changes in sexual performance and biochemical characterisation of functional neural regions: A study in serotonin transporter knockout male rats. Andrologia 2019, 51, e13291. [Google Scholar] [CrossRef] [PubMed]
- Acebes, A.; Grosjean, Y.; Everaerts, C.; Ferveur, J.-F. Cholinergic Control of Synchronized Seminal Emissions in Drosophila. Curr. Biol. 2004, 14, 704–710. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Rodriguez, D.; Franssen, D.; Bakker, J.; Lomniczi, A.; Parent, A.-S. Cellular and molecular features of EDC exposure: Consequences for the GnRH network. Nat. Rev. Endocrinol. 2021, 17, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Kolho, K.-L.; Huhtaniemi, I. Neonatal treatment of male rats with a gonadotropin-releasing hormone antagonist impairs ejaculation and fertility. Physiol. Behav. 1989, 46, 373–377. [Google Scholar] [CrossRef]
- Myers, B.; Baum, M. Facilitation of copulatory performance in male rats by naloxone: Effects of hypophysectomy, 17 α-estradiol, and luteinizing hormone releasing hormone. Pharmacol. Biochem. Behav. 1980, 12, 365–370. [Google Scholar] [CrossRef]
- Tsai, H.-W.; Tsai, Y.-F.; Tai, M.-Y.; Yeh, K.-Y. Regional and subtype-specific loss of GnRH neurons is associated with diminished mating behavior in middle-aged male rats. Behav. Brain Res. 2014, 258, 112–118. [Google Scholar] [CrossRef]
- Azizi, V.; Oryan, S.; Khazali, H.; Hosseini, A. Central injection of neuropeptide Y modulates sexual behavior in male rats: Interaction with GnRH and kisspeptin/neurokinin B/dynorphin. Int. J. Neurosci. 2020, 1–9. [Google Scholar] [CrossRef]
- Kondo, S.; Ueda, R. Highly Improved Gene Targeting by Germline-Specific Cas9 Expression in Drosophila. Genetics 2013, 195, 715–721. [Google Scholar] [CrossRef] [Green Version]
- Brunet Avalos, C.; Maier, G.L.; Bruggmann, R.; Sprecher, S.G. Single cell transcriptome atlas of the Drosophila larval brain. Elife 2019, 8, e50354. [Google Scholar] [CrossRef]
- Vargas, V.M.; Torres, D.; Corona, F.; Vergara, M.; Gómez, L.E.; Delgado-Lezama, R.; Cueva-Rolón, R. Cholinergic facilitation of erection and ejaculation in spinal cord-transected rats. Int. J. Impot. Res. 2004, 16, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Sanders, J.; Biron, D. The importance of waiting. eLife 2014, 3, e03754. [Google Scholar] [CrossRef] [PubMed]
- Chapelle, P.-A.; Blanquart, F.; Puech, A.J.; Held, J.-P. Treatment of anejaculation in the total paraplegic by subcutaneous injection of physostigmine. Spinal Cord 1983, 21, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Guttmann, L.; Walsh, J.J. Prostigmin assessment test of fertility in spinal man. Paraplegia 1971, 9, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Flavell, S.W.; Pokala, N.; Macosko, E.Z.; Albrecht, D.R.; Larsch, J.; Bargmann, C.I. Serotonin and the Neuropeptide PDF Initiate and Extend Opposing Behavioral States in C. elegans. Cell 2013, 154, 1023–1035. [Google Scholar] [CrossRef] [Green Version]
- Lottem, E.; Banerjee, D.; Vertechi, P.; Sarra, D.; Lohuis, M.O.; Mainen, Z.F. Activation of serotonin neurons promotes active persistence in a probabilistic foraging task. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bailey, M.R.; Goldman, O.; Bello, E.P.; Chohan, M.O.; Jeong, N.; Winiger, V.; Chun, E.; Schipani, E.; Kalmbach, A.; Cheer, J.; et al. An Interaction between Serotonin Receptor Signaling and Dopamine Enhances Goal-Directed Vigor and Persistence in Mice. J. Neurosci. 2018, 38, 2149–2162. [Google Scholar] [CrossRef] [PubMed]
- Sayin, S.; De Backer, J.-F.; Siju, K.; Wosniack, M.E.; Lewis, L.P.; Frisch, L.-M.; Gansen, B.; Schlegel, P.; Edmondson-Stait, A.; Sharifi, N.; et al. A Neural Circuit Arbitrates between Persistence and Withdrawal in Hungry Drosophila. Neuron 2019, 104, 544–558.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, Y.; Kennedy, A.; Chiu, H. (Vivian); Mohammad, F.; Claridge-Chang, A.; Anderson, D.J. Neurons that Function within an Integrator to Promote a Persistent Behavioral State in Drosophila. Neuron 2020, 105, 322–333.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crickmore, M.A.; Vosshall, L.B. Opposing Dopaminergic and GABAergic Neurons Control the Duration and Persistence of Copulation in Drosophila. Cell 2013, 155, 881–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paredes, R.G. Evaluating the Neurobiology of Sexual Reward. ILAR J. 2009, 50, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Tenk, C.M.; Wilson, H.; Zhang, Q.; Pitchers, K.K.; Coolen, L.M. Sexual reward in male rats: Effects of sexual experience on conditioned place preferences associated with ejaculation and intromissions. Horm. Behav. 2009, 55, 93–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaus, J.G.; Scardochio, T.; Parada, M.; Gerson, C.; Quintana, G.R.; Coria-Avila, G.A. Do rats have orgasms? Socioaffective Neurosci. Psychol. 2016, 6, 31883. [Google Scholar] [CrossRef] [Green Version]
- Shao, L.; Saver, M.; Chung, P.; Ren, Q.; Lee, T.; Kent, C.F.; Heberlein, U. Dissection of theDrosophilaneuropeptide F circuit using a high-throughput two-choice assay. Proc. Natl. Acad. Sci. USA 2017, 114, E8091–E8099. [Google Scholar] [CrossRef] [Green Version]
- Koliada, A.; Gavrilyuk, K.; Burdylyuk, N.; Strilbytska, O.; Storey, K.B.; Kuharskii, V.; Lushchak, O.; Vaiserman, A. Mating status affects Drosophila lifespan, metabolism and antioxidant system. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 246, 110716. [Google Scholar] [CrossRef]
- Wu, Q.; Yu, G.; Cheng, X.; Gao, Y.; Fan, X.; Yang, D.; Xie, M.; Wang, T.; Piper, M.D.W.; Yang, M. Sexual dimorphism in the nutritional requirement for adult lifespan inDrosophila melanogaster. Aging Cell 2020, 19, e13120. [Google Scholar] [CrossRef] [Green Version]
- Kuo, S.-Y.; Wu, C.-L.; Hsieh, M.-Y.; Lin, C.-T.; Wen, R.-K.; Chen, L.-C.; Chen, Y.-H.; Yu, Y.-W.; Wang, H.-D.; Su, Y.-J.; et al. PPL2ab neurons restore sexual responses in aged Drosophila males through dopamine. Nat. Commun. 2015, 6, 7490. [Google Scholar] [CrossRef] [Green Version]
- Koppik, M.; Ruhmann, H.; Fricke, C. The effect of mating history on male reproductive ageing in Drosophila melanogaster. J. Insect Physiol. 2018, 111, 16–24. [Google Scholar] [CrossRef]
- Grover, D.; Katsuki, T.; Li, J.; Dawkins, T.J.; Greenspan, R.J. Imaging brain activity during complex social behaviors in Drosophila with Flyception2. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraus, S.W.; Voon, V.; Potenza, M.N. Should compulsive sexual behavior be considered an addiction? Addiction 2016, 111, 2097–2106. [Google Scholar] [CrossRef] [PubMed]
- Krueger, R.B. Diagnosis of hypersexual or compulsive sexual behavior can be made using ICD-10 and DSM-5 despite rejection of this diagnosis by the American Psychiatric Association. Addiction 2016, 111, 2110–2111. [Google Scholar] [CrossRef]
- Demos, K.E.; Heatherton, T.F.; Kelley, W.M. Individual Differences in Nucleus Accumbens Activity to Food and Sexual Images Predict Weight Gain and Sexual Behavior. J. Neurosci. 2012, 32, 5549–5552. [Google Scholar] [CrossRef] [PubMed]
- LeGates, T.A.; Kvarta, M.; Tooley, J.R.; Francis, T.C.; Lobo, M.K.; Creed, M.; Thompson, S.M. Reward behaviour is regulated by the strength of hippocampus–nucleus accumbens synapses. Nat. Cell Biol. 2018, 564, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Alekseyenko, O.V.; Lee, C.; Kravitz, E.A. Targeted Manipulation of Serotonergic Neurotransmission Affects the Escalation of Aggression in Adult Male Drosophila melanogaster. PLoS ONE 2010, 5, e10806. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Chiu, H. (Vivian); Pfeiffer, B.D.; Wong, A.M.; Hoopfer, E.D.; Rubin, G.M.; Anderson, D.J. A Circuit Node that Integrates Convergent Input from Neuromodulatory and Social Behavior-Promoting Neurons to Control Aggression in Drosophila. Neuron 2017, 95, 1112–1128.e7. [Google Scholar] [CrossRef]
- Kravitz, E.A.; Fernandez, M.D.L.P. Aggression in Drosophila. Behav. Neurosci. 2015, 129, 549–563. [Google Scholar] [CrossRef]
- Gospocic, J.; Shields, E.J.; Glastad, K.M.; Lin, Y.; Penick, C.; Yan, H.; Mikheyev, A.; Linksvayer, T.; Garcia, B.A.; Berger, S.; et al. The Neuropeptide Corazonin Controls Social Behavior and Caste Identity in Ants. Cell 2017, 170, 748–759.e12. [Google Scholar] [CrossRef] [PubMed]
- Burke, C.; Huetteroth, W.; Owald, D.; Perisse, E.; Krashes, M.J.; Das, G.; Gohl, D.; Silies, M.; Certel, S.; Waddell, S. Layered reward signalling through octopamine and dopamine in Drosophila. Nat. Cell Biol. 2012, 492, 433–437. [Google Scholar] [CrossRef] [Green Version]
- Hammer, M. An identified neuron mediates the unconditioned stimulus in associative olfactory learning in honeybees. Nat. Cell Biol. 1993, 366, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Hammer, M.; Menzel, R. Multiple sites of associative odour learning as revealed by local brain microinjections of octopamine in honeybees. Learn. Mem. 1998, 5, 146–156. [Google Scholar] [PubMed]
- Schwaerzel, M.; Monastirioti, M.; Scholz, H.; Friggi-Grelin, F.; Birman, S.; Heisenberg, M. Dopamine and Octopamine Differentiate between Aversive and Appetitive Olfactory Memories inDrosophila. J. Neurosci. 2003, 23, 10495–10502. [Google Scholar] [CrossRef] [Green Version]
- Aleyasin, H.; Flanigan, M.E.; Russo, S.J. Neurocircuitry of aggression and aggression seeking behavior: Nose poking into brain circuitry controlling aggression. Curr. Opin. Neurobiol. 2018, 49, 184–191. [Google Scholar] [CrossRef]
- Golden, S.A.; Jin, M.; Shaham, Y. Animal Models of (or for) Aggression Reward, Addiction, and Relapse: Behavior and Circuits. J. Neurosci. 2019, 39, 3996–4008. [Google Scholar] [CrossRef] [Green Version]
- Kaun, K.; Devineni, A.V.; Heberlein, U. Drosophila melanogaster as a model to study drug addiction. Qual. Life Res. 2012, 131, 959–975. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, A.S.; Rothenfluh, A. I Believe I Can Fly!: Use of Drosophila as a Model Organism in Neuropsychopharmacology Research. Neuropsychopharmacology 2015, 41, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- Bath, E.; Thomson, J.; Perry, J.C. Anxiety-like behaviour is regulated independently from sex, mating status and the sex peptide receptor in Drosophila melanogaster. Anim. Behav. 2020, 166, 1–7. [Google Scholar] [CrossRef]
- Mohammad, F.; Aryal, S.; Ho, J.; Stewart, J.; Norman, N.A.; Tan, T.L.; Eisaka, A.; Claridge-Chang, A. Ancient Anxiety Pathways Influence Drosophila Defense Behaviors. Curr. Biol. 2016, 26, 981–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaumberg, K.; Zerwas, S.; Goodman, E.; Yilmaz, Z.; Bulik, C.M.; Micali, N. Anxiety disorder symptoms at age 10 predict eating disorder symptoms and diagnoses in adolescence. J. Child Psychol. Psychiatry 2019, 60, 686–696. [Google Scholar] [CrossRef] [Green Version]
- Raney, T.; Thornton, L.M.; Berrettini, W.; Brandt, H.; Crawford, S.; Fichter, M.M.; Halmi, K.A.; Johnson, C.; Kaplan, A.S.; LaVia, M.; et al. Influence of overanxious disorder of childhood on the expression of anorexia nervosa. Int. J. Eat. Disord. 2008, 41, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Cuesto, G.; Everaerts, C.; León, L.G.; Acebes, A. Molecular bases of anorexia nervosa, bulimia nervosa and binge eating disorder: Shedding light on the darkness. J. Neurogenetics 2017, 31, 266–287. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, D.S.; Horton, R.Y.; Aamodt, E.J. Role of the evolutionarily conserved starvation response in anorexia nervosa. Mol. Psychiatry 2010, 16, 595–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, H.J.; Initiative, A.N.G.; Yilmaz, Z.; Thornton, L.M.; Hübel, C.; Coleman, J.R.I.; Gaspar, H.A.; Bryois, J.; Hinney, A.; Leppä, V.; et al. Genome-wide association study identifies eight risk loci and implicates metabo-psychiatric origins for anorexia nervosa. Nat. Genet. 2019, 51, 1207–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagou, V.; Meta-Analyses of Glucose and Insulin-related traits Consortium (MAGIC); Mägi, R.; Hottenga, J.-J.; Grallert, H.; Perry, J.R.B.; Bouatia-Naji, N.; Marullo, L.; Rybin, D.; Jansen, R.; et al. Sex-dimorphic genetic effects and novel loci for fasting glucose and insulin variability. Nat. Commun. 2021, 12, 1–18. [Google Scholar] [CrossRef]
- Williams, M.J.; Akram, M.; Barkauskaite, D.; Patil, S.; Kotsidou, E.; Kheder, S.; Vitale, G.; Filaferro, M.; Blemings, S.W.; Maestri, G.; et al. CCAP regulates feeding behavior via the NPF pathway in Drosophila adults. Proc. Natl. Acad. Sci. USA 2020, 117, 7401–7408. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.R. Drosophila melanogaster as a model for nutrient regulation of ovarian function. Reproduction 2019, REP-18-0593.R3. [Google Scholar] [CrossRef]
- Ables, E.T.; Armstrong, A.R. Nutritional, hormonal, and metabolic drivers of development. Dev. Biol. 2021, 476, 171–172. [Google Scholar] [CrossRef]
- Anderson, D.J. Circuit modules linking internal states and social behaviour in flies and mice. Nat. Rev. Neurosci. 2016, 17, 692–704. [Google Scholar] [CrossRef]
- Pribram, K.H. Chapter 7: Comparative Neurology and the Evolution of Behavior. In Behavior and Evolution; Roe, A., Simpson, G.G., Eds.; Yale University Press: London, UK, 1958; pp. 14–164. [Google Scholar]
- Pribram, K.H. A Review of Theory in Physiological Psychology. Annu. Rev. Psychol. 1960, 11, 1–40. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, Z.; Tondravi, M.; Oliver, R.; Vonhoff, F.J. Drosophila Corazonin Neurons as a Hub for Regulating Growth, Stress Responses, Ethanol-Related Behaviors, Copulation Persistence and Sexually Dimorphic Reward Pathways. J. Dev. Biol. 2021, 9, 26. https://doi.org/10.3390/jdb9030026
Khan Z, Tondravi M, Oliver R, Vonhoff FJ. Drosophila Corazonin Neurons as a Hub for Regulating Growth, Stress Responses, Ethanol-Related Behaviors, Copulation Persistence and Sexually Dimorphic Reward Pathways. Journal of Developmental Biology. 2021; 9(3):26. https://doi.org/10.3390/jdb9030026
Chicago/Turabian StyleKhan, Ziam, Maya Tondravi, Ryan Oliver, and Fernando J. Vonhoff. 2021. "Drosophila Corazonin Neurons as a Hub for Regulating Growth, Stress Responses, Ethanol-Related Behaviors, Copulation Persistence and Sexually Dimorphic Reward Pathways" Journal of Developmental Biology 9, no. 3: 26. https://doi.org/10.3390/jdb9030026