
The Effects of Biochar on Indigenous Arbuscular Mycorrhizae Fungi from Agroenvironments

 , , , and
, , , and
Abstract
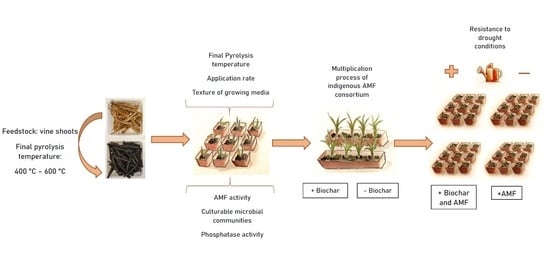
:
1. Introduction
2. Results
2.1. Biochar Properties
2.2. Soils Characterization
2.3. Experiment 1: Effects of Soil Texture, Addition Rates and Final Pyrolysis Temperatures on AMF Performance, Microbial Communities’ Variation and Phosphatase Activity
2.4. Experiment 2: Multiplication Process of AMF Consortium on Solid Substrate with Biochar
2.5. Experiment 3: Effects of Inoculum Application on Water-Stress Resistance and Development of Lettuce Plants
3. Discussion
4. Materials and Methods
4.1. Biochar Production, Characterization and Processing
4.2. Soils Characterization
4.3. Experimental Designs and Agronomic Tests Establishment
4.4. Substrate Analysis and Plant Measurements
4.5. Statistical Analysis of Results
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hüseyin, B.; Hüdaverdi, G. Drought Stress Due To Climate Change. In Proceedings of the 8th Atmospheric Sciences Symposium (ATMOS 2017), Istanbul, Turkey, 1–4 November 2017. [Google Scholar]
- Lehmann, J.; Joseph, S. Biochar for Environmental Management: An Introduction. In Biochar for Environmental Management, Science and Technology; Lehmann, J., Joseph, S., Eds.; Earthscan: London, UK, 2009; pp. 1–12. [Google Scholar]
- Paz-Ferreiro, J.; Méndez, A.; Gascó, G.; Guo, M.; He, Z.; Uchimiya, S.M. Application of Biochar for Soil Biological Improvement. In Agricultural and Environmental Applications of Biochar: Advances and Barriers; Guo, M., He, Z., Uchimiya, S.M., Eds.; Soil Science Society of America, Inc.: Madison, MA, USA, 2016; pp. 145–174. [Google Scholar]
- Arif, M.; Ilyas, M.; Riaz, M.; Ali, K.; Shah, K.; Ul Haq, I.; Fahad, S. Biochar improves phosphorus use efficiency of organic-inorganic fertilizers, maize-wheat productivity and soil quality in a low fertility alkaline soil. Field Crop. Res. 2017, 214, 25–37. [Google Scholar] [CrossRef]
- Song, D.; Xi, X.; Zheng, Q.; Liang, G.; Zhou, W.; Wang, X. Soil nutrient and microbial activity responses to two years after maize straw biochar application in a calcareous soil. Ecotoxicol. Environ. Saf. 2019, 180, 348–356. [Google Scholar] [CrossRef]
- Yadav, V.; Jain, S.; Mishra, P.; Khare, P.; Shukla, A.K.; Karak, T.; Singh, A.K. Amelioration in nutrient mineralization and microbial activities of sandy loam soil by short term field aged biochar. Appl. Soil Ecol. 2019, 138, 144–155. [Google Scholar] [CrossRef]
- Igalavithana, A.D.; Ok, Y.S.; Usman, A.R.A.; Al-wabel, M.I.; Oleszczuk, P.; Lee, S.S. The Effects of Biochar Amendment on Soil Fertility Could Biochar Be Used as a Fertilizer. In Agricultural and Environmental Applications of Biochar: Advances and Barriers; Guo, M., He, Z., Uchimiya, S.M., Eds.; Soil Science Society of America, Inc.: Madison, MA, USA, 2016; pp. 123–144. [Google Scholar]
- De Melo Carvalho, M.T.; De Holanda Nunes Maia, A.; Madari, B.E.; Bastiaans, L.; Van Oort, P.A.J.; Heinemann, A.B.; Da Silva, M.A.S.; Petter, F.A.; Marimon, B.H.; Meinke, H. Biochar increases plant-available water in a sandy loam soil under an aerobic rice crop system. Solid Earth 2014, 5, 939–952. [Google Scholar] [CrossRef] [Green Version]
- Lei, O.; Zhang, R. Effects of biochars derived from different feedstocks and pyrolysis temperatures on soil physical and hydraulic properties. J. Soils Sediments 2013, 13, 1561–1572. [Google Scholar] [CrossRef]
- Zhang, Q.; Song, Y.; Wu, Z.; Yan, X.; Gunina, A.; Kuzyakov, Y.; Xiong, Z. Effects of six-year biochar amendment on soil aggregation, crop growth, and nitrogen and phosphorus use efficiencies in a rice-wheat rotation. J. Clean. Prod. 2020, 242, 118435. [Google Scholar] [CrossRef]
- Xing, D.; Magdouli, S.; Zhang, J.; Koubaa, A. Microbial remediation for the removal of inorganic contaminants from treated wood: Recent trends and challenges. Chemosphere 2020, 258, 127429. [Google Scholar] [CrossRef] [PubMed]
- Chica, A.; Artola, A.; Rosal, A.; Solé-Mauri, F.; Fernandez, F.; García, J.; Dios, M.; Díaz, M.; Ramón, G.; Font, X. De Residuo a Recurso, El camino hacia la Sostenibilidad. In III. Recursos Orgánicos. Aspectos Agronómicos y Mediambientales. España. Capítulo 8: Enmiendas Orgánicas y de Nueva Generación: Biochar y Otras Biomoléculas; Red Española de Compostaje, Ed.; Mundi-Prensa: Madrid, Spain, 2015; p. 290. [Google Scholar]
- Jaizme-Vega, M.C. Los microorganismos rizosféricos: Bioindicadores de sostenibilidad en suelos de tomate en las Islas Canarias. AE. Revista Agroecológica de Divulgación 2015, 20, 18–19. [Google Scholar]
- Alizadeh, V.; Shokri, V.; Soltani, A.; Yousefi, M.A. Effects of Climate Change and Drought-Stress on Plant Physiology. Int. J. Adv. Biol. Biomed. Res. 2014, 2, 468–472. [Google Scholar]
- Barea, J.M.; Palenzuela, J.; Cornejo, P.; Sánchez-Castro, I.; Navarro-Fernández, C.; Lopéz-García, A.; Estrada, B.; Azcón, R.; Ferrol, N.; Azcón-Aguilar, C. Ecological and functional roles of mycorrhizas in semi-arid ecosystems of Southeast Spain. J. Arid Environ. 2011, 75, 1292–1301. [Google Scholar] [CrossRef]
- Ruíz-lozano, J.M.; Perálvarez, C.; Aroca, R. The application of a treated sugar beet waste residue to soil modifies the responses of mycorrhizal and non mycorrhizal lettuce plants to drought stress. Plant Soil 2011, 153–166. [Google Scholar] [CrossRef]
- Jaafar, N.M. Biochar as a Habitat for Arbuscular Mycorrhizal Fungi. In Mycorrhizal Fungi: Use in Sustainable Agriculture and Land Restoration. Soil Biology; Solaiman, Z., Abbott, L., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 41. [Google Scholar] [CrossRef]
- Jaizme-Vega, M.C. Microorganismos funcionales del suelo. Su papel en el manejo ecológico de los secanos. In Agricultura Ecológica en Secano. Soluciones Sostenibles en Ambientes Mediterráneos; Ministerio de Medio Ambiente y Medio Rural y Marino, Ed.; Mundi-Prensa: Madrid, Spain, 2011; pp. 417–439. [Google Scholar]
- Warnock, D.D.; Mummey, D.L.; McBride, B.; Major, J.; Lehmann, J.; Rillig, M.C. Influences of non-herbaceous biochar on arbuscular mycorrhizal fungal abundances in roots and soils: Results from growth-chamber and field experiments. Appl. Soil Ecol. 2010, 46, 450–456. [Google Scholar] [CrossRef]
- Amendola, C.; Montagnoli, A.; Terzaghi, M.; Trupiano, D.; Oliva, F.; Baronti, S.; Miglietta, F.; Chiatante, D.; Scippa, G.S. Short-term effects of biochar on grapevine fine root dynamics and arbuscular mycorrhizae production. Agric. Ecosyst. Environ. 2017, 239, 236–245. [Google Scholar] [CrossRef]
- Cobb, A.B.; Wilson, G.W.T.; Goad, C.L.; Grusak, M.A. Influence of alternative soil amendments on mycorrhizal fungi and cowpea production. Heliyon 2018, 4, e00704. [Google Scholar] [CrossRef] [Green Version]
- LeCroy, C.; Masiello, C.A.; Rudgers, J.A.; Hockaday, W.C.; Silberg, J.J. Nitrogen, biochar, and mycorrhizae: Alteration of the symbiosis and oxidation of the char surface. Soil Biol. Biochem. 2013, 58, 248–254. [Google Scholar] [CrossRef] [Green Version]
- Vanek, S.J.; Lehmann, J. Phosphorus availability to beans via interactions between mycorrhizas and biochar. Plant Soil 2015, 395, 105–123. [Google Scholar] [CrossRef]
- Ameloot, N.; Sleutel, S.; Das, K.C.; Kanagaratnam, J.; de Neve, S. Biochar amendment to soils with contrasting organic matter level: Effects on N mineralization and biological soil properties. GCB Bioenergy 2015, 7, 135–144. [Google Scholar] [CrossRef]
- Solaiman, Z.M.; Abbott, L.K.; Murphy, D.V. Biochar phosphorus concentration dictates mycorrhizal colonisation, plant growth and soil phosphorus cycling. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hammer, E.C.; Balogh-Brunstad, Z.; Jakobsen, I.; Olsson, P.A.; Stipp, S.L.S.; Rillig, M.C. A mycorrhizal fungus grows on biochar and captures phosphorus from its surfaces. Soil Biol. Biochem. 2014, 77, 252–260. [Google Scholar] [CrossRef]
- Medynska-Juraszek, A.; Cwielag-Piasecka, I. Biochar as a growing media component. In Biochar as a Renewable-Based Material: With Applications in Agriculture, the Environment and Energy; Manyà, J.J., Gascó, G., Eds.; World Scientific Publishing Europe Ltd.: London, UK, 2021; pp. 85–104. [Google Scholar]
- Sashidhar, P.; Kochar, M.; Singh, B.; Gupta, M.; Cahill, D.; Adholeya, A.; Dubey, M. Biochar for delivery of agri-inputs: Current status and future perspectives. Sci. Total Environ. 2020, 703, 134892. [Google Scholar] [CrossRef]
- Medina Peñafiel, A. Estudio de la Interacción Entre Inoculantes Microbianos y Residuos Agroindustriales Biotransformados Para su uso en Estrategias de Revegetación y Bioremediación. Ph.D. Thesis, Universidad de Granada, Granada, Spain, 2006. [Google Scholar]
- Videgain-Marco, M.; Marco-Montori, P.; Martí-Dalmau, C.; Jaizme-Vega, M.D.C.; Manyà-Cervelló, J.J.; García-Ramos, F.J. Effects of Biochar Application in a Sorghum Crop under Greenhouse Conditions: Growth Parameters and Physicochemical Fertility. Agronomy 2020, 10, 104. [Google Scholar] [CrossRef] [Green Version]
- Jaizme-vega, M.C.; Rodríguez-romero, A.S. Integración de microorganismos benéficos (Hongos micorrícicos y bacterias rizosféricas) en agrosistemas de las Islas Canarias. Agroecología 2008, 3, 33–40. [Google Scholar]
- Qayyum, M.F.; Haider, G.; Iqbal, M.; Hameed, S.; Ahmad, N.; ur Rehman, M.Z.; Majeed, A.; Rizwan, M.; Ali, S. Effect of alkaline and chemically engineered biochar on soil properties and phosphorus bioavailability in maize. Chemosphere 2021, 266. [Google Scholar] [CrossRef] [PubMed]
- Warnock, D.D.; Lehmann, J.; Kuyper, T.W.; Rillig, M.C. Mycorrhizal responses to biochar in soil—Concepts and mechanisms. Plant Soil 2007, 300, 9–20. [Google Scholar] [CrossRef]
- González, A.J.H. Posibilidades de la Producción de Inóculo de Micorrizas Vesiculo-Arbusculares Sobre Sustratos Canarios de Origen Volcánico; Trabajo fin de carrera Universidad de La Laguna: Tenerife, Spain, 1993; p. 124. [Google Scholar]
- Etesami, H.; Maheshwari, D.K. Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 156, 225–246. [Google Scholar] [CrossRef] [PubMed]
- Pérez-De-Luque, A.; Tille, S.; Johnson, I.; Pascual-Pardo, D.; Ton, J.; Cameron, D.D. The interactive effects of arbuscular mycorrhiza and plant growth-promoting rhizobacteria synergistically enhance host plant defences against pathogen. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Ordoñez, Y. Interacción sinérgica entre hongos formadores de micorrizas arbusculares—Pseudomonas fluorescens y su relación en la nutrición vegetal de fósforo. In Trabajo de Grado, Magíster en Ciencias; Facultad de Ciencias, Maestría en microbiología; Universidad Nacional de Colombia: Bogotá, Colombia, 2009. [Google Scholar]
- Nadeem, S.M.; Imran, M.; Naveed, M.; Khan, M.Y.; Ahmad, M.; Zahir, Z.A.; Crowley, D.E. Synergistic use of biochar, compost and plant growth-promoting rhizobacteria for enhancing cucumber growth under water deficit conditions. J. Sci. Food Agric. 2017, 97, 5139–5145. [Google Scholar] [CrossRef]
- Olander, L.P.; Vitousek, P.M. Regulation of soil phosphatase and chitinase activity by N and P availability. Biogeochemistry 2000, 49, 175–190. [Google Scholar] [CrossRef]
- Treseder, K.K.; Vitousek, P.M. Effects of soil nutrient availability on investment in acquisition of N and P in Hawaiian rain forests. Ecology 2001, 82, 946–954. [Google Scholar] [CrossRef]
- Dick, W.A.; Cheng, L.; Wang, P. Soil acid and alkaline phosphatase activity as pH adjustment indicators. Soil Biol. Biochem. 2000, 32, 1915–1919. [Google Scholar] [CrossRef]
- Paz-Ferreiro, J.; Gascó, G.; Gutiérrez, B.; Méndez, A. Soil biochemical activities and the geometric mean of enzyme activities after application of sewage sludge and sewage sludge biochar to soil. Biol. Fertil. Soils 2012, 48, 511–517. [Google Scholar] [CrossRef]
- Masto, R.E.; Kumar, S.; Rout, T.K.; Sarkar, P.; George, J.; Ram, L.C. Biochar from water hyacinth (Eichornia crassipes) and its impact on soil biological activity. Catena 2013, 111, 64–71. [Google Scholar] [CrossRef]
- Khadem, A.; Raiesi, F. Response of soil alkaline phosphatase to biochar amendments: Changes in kinetic and thermodynamic characteristics. Geoderma 2019, 337, 44–54. [Google Scholar] [CrossRef]
- Jin, Y.; Liang, X.; He, M.; Liu, Y.; Tian, G.; Shi, J. Manure biochar influence upon soil properties, phosphorus distribution and phosphatase activities: A microcosm incubation study. Chemosphere 2016, 142, 128–135. [Google Scholar] [CrossRef]
- Tanure, M.M.C.; da Costa, L.M.; Huiz, H.A.; Fernandes, R.B.A.; Cecon, P.R.; Pereira Junior, J.D.; da Luz, J.M.R. Soil water retention, physiological characteristics, and growth of maize plants in response to biochar application to soil. Soil Tillage Res. 2019, 192, 164–173. [Google Scholar] [CrossRef]
- Paneque, M.; De la Rosa, J.M.; Franco-Navarro, J.D.; Colmenero-Flores, J.M.; Knicker, H. Effect of biochar amendment on morphology, productivity and water relations of sunflower plants under non-irrigation conditions. Catena 2016, 147, 280–287. [Google Scholar] [CrossRef] [Green Version]
- Aller, D.; Rathke, S.; Laird, D.; Cruse, R.; Hatfield, J. Impacts of fresh and aged biochars on plant available water and water use efficiency. Geoderma 2017, 307, 114–121. [Google Scholar] [CrossRef]
- Sara, M.; Rouissi, T.; Brar, S.K.; Blais, J.F. Life Cycle Analysis of Potential Substrates of Sustainable Biorefinery; Elsevier Incorporation: Amsterdam, The Netherlands, 2016; ISBN 9780128029800. [Google Scholar]
- Bashan, Y.; de-Bashan, L.E.; Prabhu, S.R.; Hernandez, J.P. Advances in plant growth-promoting bacterial inoculant technology: Formulations and practical perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Hale, L.; Luth, M.; Crowley, D. Biochar characteristics relate to its utility as an alternative soil inoculum carrier to peat and vermiculite. Soil Biol. Biochem. 2015, 81, 228–235. [Google Scholar] [CrossRef]
- Ghazi, A. Potential for Biochar as an Alternate Carrier to Peat Moss for the Preparation of Rhizobia Bio Inoculum. Microbiol. Res. J. Int. 2017, 18, 1–9. [Google Scholar] [CrossRef]
- Liang, C.; Gascó, G.; Fu, S.; Méndez, A.; Paz-Ferreiro, J. Biochar from pruning residues as a soil amendment: Effects of pyrolysis temperature and particle size. Soil Tillage Res. 2016, 164, 3–10. [Google Scholar] [CrossRef]
- Greco, G.; Videgain, M.; Di Stasi, C.; González, B.; Manyà, J.J. Evolution of the mass-loss rate during atmospheric and pressurized slow pyrolysis of wheat straw in a bench-scale reactor. J. Anal. Appl. Pyrolysis 2018, 136, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Porter, W.M. The “most probable number” method for enumerating infective propagules of vesicular arbuscular mycorrhizal fungi in soil. Aust. J. Soil Res. 1979, 17, 515–519. [Google Scholar] [CrossRef]
- Sieverding, E. Vesicular-Arbuscular Mycorrhiza Management in Tropical Agrosystems; GTZ, D., Ed.; TZ-Verlagsgesellschaft: Eschborn, Germany, 1991. [Google Scholar]
- De Prager, S. Metodologías Básicas Para el Trabajo con Micorriza Arbuscular y Hongos Formadores de Micorriza Arbuscular; Universidad Nacional de Colombia, Ed.; Universidad Nacional de Colombia: Palmira, Colombia, 2016; p. 139. ISBN 9789588095608. [Google Scholar]
- Phillips, N.C.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment to infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular—Arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- Gerdemann, J.W.; Nicholson, T.H. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanthing. Trans. Br. Mycol. Soc. 1963, 46, 235–244. [Google Scholar] [CrossRef]
- Oehl, F.; Sieverding, E.; Palenzuela, J.; Ineichen, K.; Alves da Silva, G. Advances in Glomeromycota taxonomy and classification. IMA Fungus 2011, 2, 191–199. [Google Scholar] [CrossRef]
- Bawazir, A.M.A.; Shivanna, G.B.; Shantaram, M. Impact of Different Media for Growth and Production of Different Soluble Pigments in Actinomycetes Isolated from Soils of Hadhramout, Yemen. Eur. J. Biomed. 2018, 5, 615–619. [Google Scholar]
- Yao, Q.; Gao, J.L.; Zhu, H.H.; Long, L.K.; Xing, Q.X.; Chen, J.Z. Evaluation of the potential of trap plants to detect arbuscular mycorrhizal fungi using polymerase chain reaction-denaturing gradient gel electrophoresis analysis. Soil Sci. Plant Nutr. 2010, 56, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Jaizme-vega, M.C. Las Micorrizas, Una Estrategia Agroecológica Para Optimizar la Calidad de los Cultivos; Universidad de la Laguna, Instituto Canario de Investigaciones Agrarias, Phytoma España S.L., Eds.; Phytoma España S.L.: Valencia—San Cristóbal de la Laguna (Tenerife), Spain, 2019; p. 112. ISBN 978-84-946691-5-6. [Google Scholar]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Determination 1 | Unit | S1 2 | S2 |
|---|---|---|---|
| AMF potential (MPN) | Number of infective mycorrhizal propagules 100 | 39.6 ± 9.3 | 30.9 ± 13.1 |
| Number of AMF spores | Number of AMF spores 100 g soil−1 | 420 ± 18 | 465 ± 23 |
| Identified genera | % spores | 20 Gigaspora spp. 3 Scutellospora spp. 77 Glomus spp. | 6 Acaulospora spp. 94 Glomus spp. |
| Culturable microbial communities | |||
| Mesophilic aerobic microorganisms | log cfu g−1 | 6.59 ± 0.90 | 7.08 ± 0.67 |
| Actinomycetes | log cfu g−1 | 6.08 ± 0.45 | 6.45 ± 0.27 |
| Pseudomonas genus | log cfu g−1 | 5.74 ± 0.13 | 5.96 ± 0.34 |
| Mycobiota | log cfu g−1 | 4.75 ± 0.65 | 4.71 ± 0.43 |
| Variable Measured | Sandy-Loam Substrate | Clay-Loam Substrate | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| D0 | B400 | B600 | D0 | B400 | B600 | ||||||
| D1 | D2 | D1 | D2 | D1 | D2 | D1 | D2 | ||||
| Root AMF colonization % | D120 | 10.0 a (1.0) | 13.3 a (5,7) | 12.5 a (1.0) | 10.0 a (1.5) | 10.0 a (1.5) | 10.5 a (2.0) | 13.3 a (6.7) | 10.0 a (0.0) | 11.6 a (4.0) | 13.3 a (5.7) |
| D210 | 11.7 a (2.8) | 18.3 a (5.7) | 16.7 a (2.9) | 11.7 a (2.8) | 13.3 a (2.9) | 21.7 a (5.7) | 21.7 a (5.7) | 13.3 a (28.9) | 13.3 a (2.8) | 15.0 a (0.0) | |
| D330 | 8.3 b (2.9) | 13.3 ab (2.9) | 20.0 a (0.0) | 10.0 b (5.0) | 11.7 ab (2.9) | 26.7 ab (2.9) | 26.7 ab (7.3) | 31.7 a (2.9) | 26.7 ab (2.9) | 18.3 b (2.9) | |
| D390 | 5.7 bc (1.1) | 13.3 ab (2.9) | 18.3 a (2.9) | 5.0 c (0.0) | 10.0 bc (0.0) | 25.0 a (0.0) | 23.3 a (2.9) | 31.6 a (2.9) | 25.0 a (0.0) | 6.7 b (5.8) | |
| Number of AMF spores 100 g soil−1 | D390 | 548 c (103) | 830 bc (198) | 1511 a (170) | 1380 ab (275) | 1095 ab (230) | 1668 b (200) | 2058 b (232) | 1930 b (287) | 1978 b (95) | 5245 a (1399) |
| Factor | Root AMF Colonization | AMF Spores | Phosphatase Activity | Microbial Communities | ||||
|---|---|---|---|---|---|---|---|---|
| D210 | D330 | D390 | AcdP | AlkP | MAM | PS | ||
| Growing substrate texture (S) | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.040 | ||
| Biochar temperature (B) | 0.001 | <0.0001 | 0.001 | <0.0001 | <0.0001 | |||
| Application rate (D) | <0.0001 | <0.0001 | ||||||
| S B | 0.006 | 0.012 | ||||||
| S D | 0.002 | 0.006 | <0.0001 | |||||
| B D | 0.048 | 0.007 | <0.0001 | 0.005 | 0.001 | 0.001 | ||
| S B D | <0.0001 | 0.003 | 0.001 | 0.011 | ||||
| Treatment | AMF Root Colonization (%) | Number of AMF Spores 100 g Soil−1 | Infective Mycorrhizal Propagules 100 |
|---|---|---|---|
| T0 | 15.5 ± 2.1 | 514.6 ± 59.3 | 72.4 ± 32.0 |
| T1 | 32.8 *** ± 5.2 | 866.9 *** ± 125.0 | 161.5 ** ± 80.3 |
| Factors | Root AMF Coloniz. (%) D62 | Number of AMF Spores | Dry Biomass | SPAD Index | n | ||||
|---|---|---|---|---|---|---|---|---|---|
| Irrigation | Inoculum | ADB | RDB | D30 | D50 | D62 | |||
| WID 1 | −AMF | 1.0 ± 3.0 c | 2.0 ± 4.3 c | 2.70 ± 0.35 ab | 1.28 ± 0.20 b | 27.4 ± 3.2 | 36.2 ± 2.1 a | 37.2 ± 2.5 a | 10 |
| +AMF | 13.0 ± 3.5 b | 51.8 ± 14.3 b | 3.45 ± 0.50 ab | 0.93 ± 0.23 b | 27.5 ± 1.2 | 36.9 ± 2.0 a | 37.24 ± 1.8 a | 9 | |
| +B +AMF | 24.5 ± 7.2 a | 144.1 ±35.6 ab | 3.71 ± 0.48 a | 1.26 ± 0.21 b | 29.0 ± 1.0 | 35.7 ± 2.3 a | 35.9 ± 2.6 a | 9 | |
| WID 2 | −AMF | 2.0 ± 4.2 b | 1.9 ± 4.2 c | 1.25 ± 0.45 b | 28.5 ± 3.0 | 21.6 ± 4.2 c | 0 | ||
| +AMF | 19.0 ± 10.0 a | 178.6 ± 17.9 a | 2.49 ± 0.21 c | 1.39 ± 0.39 b | 27.6 ± 3.5 | 28.1 ± 3.8 b | 16.5 ± 1.1 c | 5 | |
| +B +AMF | 24.0 ± 6.0 a | 201.3 ± 73.0 a | 2.69 ± 0.25 bc | 1.92 ± 0.60 a | 27.1 ± 4.1 | 33.7 ± 1.4 a | 17.2 ± 1.4 b | 8 | |
| Experiment | Factors | Treatments | Replicates |
|---|---|---|---|
| 1 | 1. Growing media texture | S1—substrate 1—sandy-loam growing media S2—substrate—2—clay-loam growing media | Five replicates/treatment: 50 experimental units |
| 2. Final pyrolysis temperature | B1—biochar 1—400 °C B2—biochar 2—600 °C | ||
| 3. Biochar application rate | D0—Control—without biochar | ||
| D1—1.5 wt. % | |||
| D2—3 wt. % | |||
| 2 | 1. Biochar application rate | T0—Control—without biochar | Three replicates/treatment: 6 experimental units |
| T1—1.5 wt. % B1 | |||
| 3 | 1. Inoculum composition | −AMF—Control—without inoculum | Ten replicates/treatment: 60 experimental units |
| +AMF—+ inoculum obtained from T0 | |||
| +B+AMF—+ inoculum obtained from T1 | |||
| 2. Water irrigation dose | WID1—70–80% container capacity | ||
| WID2—10% container capacity |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Videgain-Marco, M.; Marco-Montori, P.; Martí-Dalmau, C.; Jaizme-Vega, M.d.C.; Manyà-Cervelló, J.J.; García-Ramos, F.J. The Effects of Biochar on Indigenous Arbuscular Mycorrhizae Fungi from Agroenvironments. Plants 2021, 10, 950. https://doi.org/10.3390/plants10050950
Videgain-Marco M, Marco-Montori P, Martí-Dalmau C, Jaizme-Vega MdC, Manyà-Cervelló JJ, García-Ramos FJ. The Effects of Biochar on Indigenous Arbuscular Mycorrhizae Fungi from Agroenvironments. Plants. 2021; 10(5):950. https://doi.org/10.3390/plants10050950
Chicago/Turabian StyleVidegain-Marco, María, Pedro Marco-Montori, Clara Martí-Dalmau, María del Carmen Jaizme-Vega, Joan Josep Manyà-Cervelló, and Francisco Javier García-Ramos. 2021. "The Effects of Biochar on Indigenous Arbuscular Mycorrhizae Fungi from Agroenvironments" Plants 10, no. 5: 950. https://doi.org/10.3390/plants10050950