
The Roles of GRETCHEN HAGEN3 (GH3)-Dependent Auxin Conjugation in the Regulation of Plant Development and Stress Adaptation

1
College of Life Science and Technology, Gansu Agricultural University, Lanzhou 730070, China
2
State Key Laboratory of Soil and Sustainable Agriculture, Institute of Soil Science, Chinese Academy of Sciences, Nanjing 210008, China
3
University of Chinese Academy of Sciences, Beijing 100049, China
*
Authors to whom correspondence should be addressed.
†
These authors contribute equally to this work.
Plants 2023, 12(24), 4111; https://doi.org/10.3390/plants12244111
Submission received: 8 November 2023
/
Revised: 5 December 2023
/
Accepted: 7 December 2023
/
Published: 8 December 2023
(This article belongs to the Special Issue Advances in Plant Auxin Biology)
Abstract
:The precise control of free auxin (indole-3-acetic acid, IAA) gradient, which is orchestrated by biosynthesis, conjugation, degradation, hydrolyzation, and transport, is critical for all aspects of plant growth and development. Of these, the GRETCHEN HAGEN 3 (GH3) acyl acid amido synthetase family, pivotal in conjugating IAA with amino acids, has garnered significant interest. Recent advances in understanding GH3-dependent IAA conjugation have positioned GH3 functional elucidation as a hot topic of research. This review aims to consolidate and discuss recent findings on (i) the enzymatic mechanisms driving GH3 activity, (ii) the influence of chemical inhibitor on GH3 function, and (iii) the transcriptional regulation of GH3 and its impact on plant development and stress response. Additionally, we explore the distinct biological functions attributed to IAA-amino acid conjugates.

1. Introduction
Auxins, a group of phytohormones, are integral to the regulation of plant development and stress responses [1,2]. Previous research has identified three naturally occurring auxins: indole-3-acetic acid (IAA), phenylacetic acid (PAA), and 4-chloro-indole-3-acetic acid (4-Cl-IAA), with IAA being the most prevalent and significant in plants [1]. The meticulous modulation of IAA levels, governed by biosynthesis, transport, and inactivation, is essential for normal plant growth, development, and adaptation to both biotic and abiotic environmental stresses [1,3,4].
Four primary pathways have been documented for IAA inactivation in plants: (i) IAA CARBOXYL METHYLTRANSFERASE1 (IAMT1) converts IAA to methyl IAA; (ii) UDP-glucosyltransferase (UGTs) generate ester-linked IAA conjugates; (iii) GRETCHEN HAGEN 3 (GH3) acyl amido synthetases facilitate the formation of amide-linked IAA conjugates; and (iv) IAA oxidation carried out by DIOXYGENASE FOR AUXIN OXIDATION (DAO) [5,6,7,8,9,10]. Methyl IAA and ester-linked IAA, both subject to reconversion into IAA by specific hydrolases, are predominantly regarded as forms of IAA storage [11]. Contrastingly, the reversibility of amide-linked conjugates varies based on the amino acid involved. Most, such as IAA-alanine (IAA-Ala), IAA-phenylalanine (IAA-Phe), and IAA-leucine (IAA-Leu), revert to free IAA, while others like IAA-glutamate (IAA-Glu) and IAA-aspartate (IAA-Asp) are directly degraded [5,11,12,13]. However, recent findings suggest that IAA-Glu and IAA-Asp are also hydrolyzed to free IAA by IAA-LEU-RESISTANT1 (ILR1), and that DAO-dependent oxidation also occurs by oxidizing IAA-Asp and IAA-Glu to oxIAA-Asp and oxIAA-Glu, which in turn are hydrolyzed to oxIAA in the presence of the hydrolase ILR1, rather than by direct IAA oxidation to oxIAA. This suggests that GH3-dependent IAA conjugation may be a key node in IAA storage and IAA oxidative degradation [6,14].
The first GH3 gene was identified from Glycine max as a rapid early auxin-responsive gene. Subsequent research has established the widespread distribution of the GH3 gene family across the plant kingdom, encompassing species from Arabidopsis thaliana to Oryza sativa, Zea mays, Triticum aestivum, and even non-vascular plants like Physcomitrella patens and Marchantia polymorpha [8,11,15,16,17,18,19]. Based on sequence homology and substrate specificity, the GH3 family in Arabidopsis is categorized into three distinct groups: Group I, II, and III. Group I GH3 genes are known to encode enzymes that synthesize amides from jasmonic acid (JA) or salicylic acid (SA). Group II GH3 enzymes function as IAA-amido synthetases, and Group III has been shown to catalyze the conjugation between amino acids and 4-substituted benzoates or indole-3-butyric acid (IBA) [8,20]. Emerging evidence underscores the role of GH3-mediated IAA conjugation not only in modulating free IAA availability but also in its potential as a signaling molecule or inhibitor, impacting plant growth and development [21,22,23,24]. This review will delve into the biochemical mechanisms of GH3-mediated IAA conjugation and its transcriptional regulation.
2. The Catalytic Mechanisms and Substrate Specificity of GH3 Acyl Acid Amido Synthetase Enzyme
Chen et al. firstly utilized a combination of initial velocity and product inhibition analyses, alongside mass spectrometry, to delineate the kinetic and chemical mechanisms governing OsGH3.8 activity [25]. They discovered that the conjugation of IAA with Asp operates via a ‘Bi Uni Uni Bi Ping Pong’ mechanism, as depicted in Figure 1A. The process initiates with the binding of IAA and ATP, in the presence of Mg2+, to the unoccupied enzyme. This interaction results in the formation of an adenylated IAA intermediate (IAA-AMP) and the concurrent release of pyrophosphate (PPi). Following this, Asp attaches to the enzyme•IAA•AMP complex, leading to the displacement of AMP and the establishment of an amide linkage between IAA and Asp. The final reaction products, IAA-Asp and AMP, are then released from the OsGH3.8 enzyme’s active site [25].
Structural analyses of the GH3 enzyme in Arabidopsis, grape, and rice have illuminated that both monocotyledons and dicotyledons employ a similar mechanism for AMP and IAA binding [26]. The GH3 enzyme exhibits distinct acyl acid binding preferences, with specific residues within its active site conferring selectivity for particular substrates [26]. In OsGH3.8, the amino acids Arg130 and Leu137 play a crucial role in substrate specificity. The mutation of Arg130 to Leu (Arg130-Leu) shifts the enzyme’s substrate preference from IAA to benzoate/SA, while an Arg130-Thr substitution favors JA over IAA. Similarly, Leu137-Ser mutation leads to a benzoate/SA preference, and the replacement of Leu137 with Arg/Ile induces a preference for JA [25,26]. GH3 proteins also exhibit amino acid specificity; for instance, the carboxylate group of Asp is a determinant for the active site’s specificity in OsGH3.8 [25,26]. Ser341 participates in adenylate formation by forming hydrogen bonds with phosphate groups [26]. Moreover, Mg2+ is essential for the enzyme’s maximum activity, aiding in AMP orientation, with Glu342 being critical for Mg2+ coordination [25,26,27]. Additionally, seven residues identified through the sequence comparison of acyl-substrate binding sites (Arg130, Leu137, Valine 174 (Val174), Leu175, Methionine 337 (Met337), Alanine 339 (Ala339), and Tyrosine 344 (Tyr344)) are thought to be involved in the substrate-specific selection of IAA [26].
It has been observed that the residues involved in the AMP binding site of acyl adenylate cleavage enzymes exhibit a high degree of conservation across the GH3 protein family. In contrast, the residues that interact with acyl substrates show variability, accommodating the binding of diverse substrates [25,26,28,29]. This variation in amino acid residues leads to the formation of different binding pockets, each tailored for specific substrates [29]. This may account for the fact that auxin function at all stages of plant growth and development, a process that offers great flexibility in regulating auxin action is necessary. The existence of a complex auxin conjugation system, as evidenced by these variations in GH3 proteins, is likely a strategic evolutionary development to facilitate this flexibility [28].
3. The Modulation of IAA Homeostasis by Small Chemical Molecules via the Inhibition of GH3 Enzyme Activity
The functional redundancy of class II GH3 enzymes in plants presents a challenge to traditional genetic approaches when exploring their biological roles [30]. To overcome this obstacle, small molecule inhibitors have emerged as a powerful alternative. These inhibitors can be applied to any plant tissue at any developmental stage, given in appropriate concentrations. Their utility lies in the ability to bypass gene redundancy and the potential detrimental effects of lethal mutations often associated with simultaneous multi-gene mutations [31]. To date, three potent inhibitors have been identified that modulate GH3-dependent IAA conjugation: adenosine-59-[2-(1H-indol-3-yl) ethyl] phosphate (AIEP), kakeimide (KKI), and N-[4-[[6-(1H-pyrazol-1-yl)-3-pyridazinyl]amino] phenyl]-3-(trifluoromethyl) benzamide (nalacin), as represented in Figure 1B [30,31,32].
3.1. AIEP, the First Chemical Inhibitor of Auxin Conjugation
The GH3-dependent IAA conjugation initiates when the unbound GH3 enzyme interacts with ATP and IAA, leading to the formation of IAA•AMP. Böttcher et al. engineered and synthesized a stable analogue of IAA•AMP, named AIEP. This molecule competes with ATP and IAA for the binding sites on the GH3 enzyme at the onset of catalysis [30]. The competitive inhibitory effect of AIEP on ATP and IAA binding was validated through substrate velocity experiments involving VvGH3.1 and VvGH3.6 from grape. However, the study did not extend to phenotypic examinations to assess the broader biological impacts of this inhibition (Figure 1).
3.2. KKI, a Specific Inhibitor of IAA-Conjugating GH3 Enzymes
In the quest to identify inhibitors of IAA-conjugating GH3 enzymes, researchers leveraged Arabidopsis AtGH3.6 overexpression plants as a biological assay system. They screened a synthetic chemical library comprising 10,000 compounds for agents capable of reverting the altered root hair growth phenotype of AtGH3.6-overexpressed lines. This led to the initial identification of compound ‘1’, followed by the synthesis of 25 derivatives of ‘1’. Among these, kakeimide (KKI) emerged as a highly potent inhibitor. (Figure 1B). KKI functions by directly interacting with the IAA binding site within the GH3•ATP complex, forming a stable GH3•ATP•KKI ternary complex that impedes the synthesis of IAA-amino acid conjugates [32]. Validation experiments demonstrated KKI’s effectiveness in targeting the IAA binding sites of various GH3 enzymes, notably VvGH3.1, AtGH3.5, and OsGH3.8, while sparing the IBA binding site of AtGH3.15. This specificity, coupled with KKI’s lack of interference in jasmonic acid (JA) homeostasis, underscores its role as a specific inhibitor of IAA-conjugating GH3 enzymes [32].
3.3. Nalacin, a Potent Inhibitor Targeting Group II GH3 Enzymes
Nalacin was identified from a chemical screen by observing the auxin-related root phenotypes in the Arabidopsis wild-type Col-0 (Figure 1B). Subsequent studies have shown that nalacin competitively inhibits substrate acceptance by AtGH3.6 and AtGH3.11 through trifluoromethyl phenyl occupancy of the IAA binding site of AtGH3s, suggesting that nalacin also functions in the first step of the ‘Bi Uni Uni Bi Ping Pong’ reaction of GH3 enzymes. Unlike KKI, which selectively inhibits only class II GH3 enzymes, nalacin also impedes the formation of JA amino acid conjugates mediated by AtGH3.11, albeit through a distinct binding mode [31]. Consequently, there is potential for further chemical modifications of nalacin to enhance its selective inhibition of different GH3 enzyme members.
4. The Transcriptional Control of GH3 Enzymes in Plant Growth, Development, and Stress Adaptation
Research into GH3-dependent IAA conjugation has revealed its critical role in the modulation of plant growth, development, and stress responses across a variety of species, with extensive studies conducted particularly in maize, wheat, rice, and Arabidopsis. The forthcoming sections will focus on the advances in understanding GH3-dependent IAA conjugation in Arabidopsis and rice. Additionally, progress in other species, reflecting the broader impact and relevance of GH3 enzymes in plant biology, has been systematically compiled in Table 1.
4.1. GH3-Dependent IAA Conjugation Is Involved in Regulating Multiple Developmental Processes
In Arabidopsis, eight Group II GH3 genes are involved in catalyzing IAA conjugation to amino acids. Due to redundant gene functions, mutations in single genes result in only subtle phenotypic changes and modified sensitivity to exogenous IAA [54]. In contrast, mutants with overexpressed GH3 genes, obtained through activation tag insertion, provide a more discernible phenotype for study. For instance, AtGH3.2 and AtGH3.6 were identified through the screening of their overexpressed mutants, ydk1-D and dfl1-D, respectively [54,55]. Interestingly, however, despite all overexpressing genes being closely related to GH3 Group II family members, they still showed inconsistent phenotypes. Under various light conditions, the dfl1-D mutant displayed shortened hypocotyls exclusively in light, while the ydk1-D mutant showed this phenotype under both light and dark conditions. Additionally, the ydk1-D mutant had a shorter primary root but did not exhibit significant difference in susceptibility to auxin-mediated root growth inhibition. In contrast, the dfl1-D mutant was resistant to IAA-mediated root growth inhibition and did not present a short-root phenotype compared with the wild type [54,55]. Furthermore, several Group II GH3 genes are transcriptionally induced by IAA, whereas AtGH3.9 is repressed in response to exogenous IAA application [56]. Altogether, these data indicate that, while Group II GH3 members share some commonalities and function in a similar pathway by regulating free IAA conjugation, they each play distinct roles in plant development, which may be due to differences in their tissue specificity and/or hormonal regulation (e.g., IAA, JA, etc.).
As early auxin response genes, Group II GH3 genes play a significant role downstream of auxin response factors (ARFs), which are key elements in the auxin signaling pathway [57]. Research has elucidated the involvement of ARF6, AtARF7, AtARF8, and AtARF17 in the transcriptional regulation of several AtGH3 genes. AtARF7 and AtARF8 are known to positively regulate the transcription of AtGH3.2, AtGH3.5, and AtGH3.6, influencing hypocotyl elongation under different light conditions [54,58]. In contrast, AtARF17 has a negative regulatory role on AtGH3.5 and AtGH3.6 transcription, which is essential for proper plant development. The microRNA AtmiR160 directly targets AtARF17 mRNA, which is crucial for normal leaf and root growth [59]. In terms of adventitious root development, AtARF6 and AtARF8 act as positive regulators, while AtARF17 functions as a negative regulator. They co-regulate the transcription of AtGH3.3, AtGH3.5, and AtGH3.6, impacting JA conjugation with amino acids, but not IAA conjugation [60]. Additionally, AtARF7 actives another auxin-induced transcription factor (TF), AtMYB77, which subsequently upregulates AtGH3.2 and AtGH3.3 to control root development [61]. AtMYB30, another MYB TF, directly binds to the promoters of AtGH3.2 and AtGH3.3, repressing their transcription to foster root elongation [62]. WRINKLED1 (WRI1), yet another TF, binds to the AtGH3.3 promoter. Although AtGH3.3 transcription increases in the Atwri1-1 mutant, AtWRI1 does not appear to repress AtGH3.3 directly, suggesting it may function as a transcriptional co-factor [63,64]. Subsequent research identified that AtTCP20, a TF that interacts with AtWRI1, binds to the AtGH3.3 promoter, and activates its transcription [63]. Electrophoretic mobility shift assays have shown that the addition of AtWRI1 decreases the binding activity of AtTCP20 to the AtGH3.3 promoter in a dose-dependent manner, indicating that AtWRI1 regulates AtGH3.3 transcription by antagonizing AtTCP20’s binding [63]. Several basic leucine zipper (bZIP) transcription factors, including AtbZIP2, 11, 44, 53, and 63, also directly bind to the AtGH3.3 promoter and enhance its transcription [65]. Furthermore, cytokinin influences the root meristem by maintaining IAA concentration, with AtARR1 directly activating AtGH3.17 transcription [66]. There are also indications of TFs AtHLS1, AtURO, and AtSTY1 being involved in modulating GH3-dependent IAA conjugation in plant developmental processes, although direct regulatory evidence is yet to be established (Figure 2) [67,68,69].
In rice, the Group II GH3 family, which includes OsGH3.1, OsGH3.2, OsGH3.5, OsGH3.8, and OsGH3.13, plays a significant role in various developmental aspects such as shoot height, leaf angle, floret fertility, and tiller number. These effects are primarily mediated through the modulation of IAA conjugation [70,71,72,73,74]. Recent findings highlight several TFs that directly activate the transcription of OsGH3 genes, leading to decreased levels of free IAA and consequent impacts on rice morphology. For example, OsbZIP49 has been shown to upregulate OsGH3.2 and OsGH3.13 by binding to TGACG motifs in their promoters. This upregulation results in reduced free IAA concentrations, affecting cell elongation mediated by expansins and thus impacting shoot gravitropism [75]. Furthermore, OsGH3.2 is regulated by OsARF8, a downstream target of OsmiR167, indicating a critical role for the OsmiR167-OsARF8-OsGH3.2 pathway in cellular auxin homeostasis, particularly in response to exogenous auxin [76]. Another ARF, OsARF19, has been found to reduce free IAA levels by activating OsGH3.5, thereby influencing leaf angulation. Intriguingly, the transcription of OsARF19 itself is induced by IAA and brassinolide (BR), suggesting the OsARF19-OsGH3.5 module’s involvement in integrating IAA-BR signals [73]. Additionally, OsSPL7, a target of OsmiR156f, directly activates OsGH3.8, affecting tiller number and shoot height [70]. The involvement of OsMADS1 and OsMADS6 in binding to the OsGH3.8 promoter and regulating floret fertility has also been documented [77]. Collectively, these findings demonstrate that Group II GH3 genes, together with their upstream TFs, form a complex regulatory network that integrates light, miRNA, and hormonal signals to control plant growth and development (Figure 3) [78].
4.2. The Integration of Hormonal Signals in GH3-Dependent IAA Conjugation’s Responses to Abiotic Stresses
4.2.1. Drought Stress
In Arabidopsis, AtGH3.5 has been observed to respond rapidly to drought conditions, with the wes1-D mutant, which overexpresses AtGH3.5, exhibiting enhanced drought resistance [79]. A subsequent study revealed that AtMYB96 modulates the expression of several AtGH3 genes, including AtGH3.3, AtGH3.5, and AtGH3.6, under drought stress through an abscisic acid (ABA)-dependent pathway [80]. This finding underscores the importance of ABA signaling in modulating GH3 gene expression during drought response. Additionally, the gh3oct mutant, with knockouts of all Group II GH3 genes (GH3.1,2,3,4,5,6,9,17), exhibits increased drought tolerance, further highlighting the role of these genes in drought response mechanisms [81]. In rice, the activation of OsGH3.13 has been linked to a reduction in free IAA levels, leading to a structural adaptation in the leaves, such as thicker blades, which enhance drought tolerance by minimizing water loss [74]. However, the response to drought stress in rice is complex, as evidenced by the contrasting effects observed with OsGH3.2. While OsGH3.2 is also upregulated in response to drought, its overexpression leads to decreased drought tolerance. This discrepancy is attributed to the inhibition of carotenoid biosynthesis by overexpressed OsGH3.2, which consequently reduces ABA synthesis. This is in stark contrast to the increased ABA levels seen in lines overexpressing OsGH3.13 [72]. These findings collectively indicate that, while GH3 genes are integral to stress responses through IAA homeostasis regulation, the distinct spatial–temporal expression patterns and secondary growth effects can result in varying stress sensitivities [72,74].
4.2.2. Temperature (Heat/Cold/Freezing) Stress
In Arabidopsis, the transcription of AtGH3.5 is notably responsive to temperature extremes, showing increased levels under both low (4 °C) and high (37 °C) temperature conditions. The wes1-D mutant, characterized by the overexpression of AtGH3.5, shows increased survival after exposure to freezing temperatures (−7 °C). This suggests a broad regulatory role for AtGH3.5 across a spectrum of temperature stresses [82]. In rice, the overexpression of OsGH3.2 leads to a reduction in free IAA levels, thereby activating cold-responsive genes and enhancing the plant’s ability to scavenge reactive oxygen species (ROS). Consequently, this confers increased resistance to cold stress [72].
4.2.3. Salt and Osmotic Stress
All root-expressed Group II GH3 genes in Arabidopsis are upregulated following treatment with NaCl at concentrations of 75 mM and 150 mM. The Atgh3oct mutant, with combined knockouts of all Group II GH3 genes, exhibits greater resilience to NaCl stress compared to the wild type [81]. This enhanced tolerance also extends to sorbitol and mannitol exposure, suggesting that Group II GH3s may confer broad osmotic stress resistance, inclusive of salinity stress. Further investigation reveals that NaCl treatment increases AtACS2 transcription, leading to the accumulation of the ethylene precursor ACC, which in turn downregulates AtGH3.5 and AtGH3.9 transcription, maintaining free IAA levels and primary root growth [83]. Additionally, both osmotic and salt stresses are known to stimulate the accumulation of ABA, which reduces free IAA content, inhibiting lateral root (LR) development. At the same time, these stresses activate AtWRKY46, a transcription factor that suppresses AtGH3.1 transcription, thus maintaining free IAA levels and LR development. Additionally, ABA inhibits AtWRKY46 transcription. This antagonistic interaction between ABA and AtWRKY46 fine-tunes free IAA levels and LR development, allowing for improved adaptation to osmotic and salt stresses [84].
4.2.4. Ammonium (NH4+) Stress
NH4+ serves as a vital nitrogen source for plants, but when available in excess, it can be detrimental to growth [85,86]. Prior research indicates that high NH4+ levels lead to a reduction in free IAA [87,88,89]. In Arabidopsis, elevated NH4+ conditions trigger the induction of nearly all Group II GH3 genes, which in turn accelerates the conjugation of free IAA to amino acids [87]. Concurrently, high NH4+ levels also enhance the transcription of AtWRKY46, a transcription factor that binds to the promoters of AtGH3.1 and AtGH3.6, repressing their transcription. This response serves to maintain free IAA levels and support primary root growth under high NH4+stress conditions. The overexpression of AtWRKY46 improves NH4+ tolerance, suggesting its critical role in modulating primary root development during NH4+ stress [90].
4.2.5. Pathogen Stress
In response to pathogen attacks, plants activate a comprehensive defense strategy: (1) they initiate a hypersensitive response leading to rapid programmed cell death at the infection site alongside other defense responses; (2) they activate systemic-acquired resistance (SAR) in distal tissues; and (3) they activate basal immunity to limit pathogen growth [91]. GH3-dependent IAA conjugation is intricately involved in these plant defense mechanisms. In Arabidopsis, the pathogens B. cinerea and P. syringae pv tomato (Pst) DC3000 significantly upregulate AtGH3.2 and AtGH3.3 transcription. Loss-of-function mutations in AtGH3.2 or AtGH3.4 enhance resistance to both pathogens, suggesting that AtGH3.2, AtGH3.3, and AtGH3.4 may negatively influence the plant’s response to B. cinerea and Pst DC3000 [92]. The overexpression of mutant gh3.5-1D shows a compromised hypersensitive response but retains normal SAR and basal immunity, whereas the Atgh3.5 mutant exhibits a defective SAR response yet maintains a typical hypersensitive response and basal immunity. In contrast, the dfl1-D mutant displays altered hypersensitive and basal immune responses [91].
In rice, IAA has been linked to increased susceptibility to various pathogens, partly due to IAA-induced expansins that relax the cell wall, a plant’s primary defense barrier [93]. Overexpressing certain Group II GH3 genes, such as OsGH3.1, OsGH3.2, and OsGH3.8, bolsters resistance against bacterial and fungal pathogens. This resistance is attributed to the suppression of pathogen-induced IAA accumulation, leading to the downregulation of expansin expression and subsequent stabilization of the cell wall [71,93,94].
The varied effects of Group II GH3 genes on pathogen stress between rice and Arabidopsis could be attributed to multiple factors. In rice, OsGH3s appear to primarily contribute to forming a robust cell wall barrier, which acts as the first line of defense against pathogen entry, without further invoking a hypersensitive response or SAR [39,94]. In contrast, the expression of AtGH3 in Arabidopsis is highly specific to tissue type and developmental stage, and unspecific overexpression could lead to unintended consequences, such as disruptions in the levels of IAA and salicylic acid (SA), which are critical for the plant’s defense response [91,92]. Additionally, the GH3-catalyzed IAA-Asp conjugate is hypothesized to act as a susceptibility signal [23]. While GH3 activity reduces free IAA levels, it simultaneously results in the accumulation of IAA-Asp. The specific distribution and balance of IAA and IAA-Asp within the plant may significantly influence the outcome of GH3’s role in pathogen response [92].
5. Atypical Roles of Group II GH3 and IAA-Amino Acids
The exploration of Group II GH3 enzymes as IAA-acyl acid amino synthetases in 2002 marked a significant advancement in our understanding of these enzymes [11]. In addition to the determination of their three-dimensional structures and the key amino acid sites for enzyme activity, research has expanded their known functions to include the conjugation of other auxins like PAA and IBA with amino acids, IAA conjugation with proteins, and the metabolism of auxinic herbicides [95]. Furthermore, AtGH3.15, previously an undefined member of Group III, is now recognized as an IBA-conjugating acyl acid amido synthetase. IBA serves as a precursor to IAA, which is converted through the peroxisomal β-oxidation pathway [96]. Intriguingly, AtGH3.13, AtGH3.14, and AtGH3.16—other members of Group III—possess acyl acid binding sites similar to AtGH3.15, suggesting their possible involvement in IAA homeostasis [20,95]. Here, we will discuss the atypical roles of Group II GH3 and IAA-amino acids.
5.1. The Roles of Group II GH3 beyond the Catalyzation of IAA-Amino Acid Conjugate Formation
The capacity of Group II GH3 enzymes extends beyond the synthesis of IAA-amino acid conjugates. Research using recombinant GH3 IAA-amino acid synthetase from pea has revealed the enzyme’s ability to conjugate IAA not only to aspartate but also to proteins in immature seeds. The proposition that IAA conjugation to proteins may serve a regulatory function acting as a prosthetic group and influencing protein activity via posttranslational modifications is a compelling avenue for further exploration [97,98]. Besides proteins, Group II GH3 enzymes also facilitate the conjugation of PAA to amino acids. PAA, another natural but less active auxin than IAA, exhibits unique distribution and transport characteristics, implying a role in sustaining the auxin equilibrium necessary for plant cellular processes [99,100,101]. Notably, PAA can stimulate GH3-dependent IAA conjugation, while high IAA levels can suppress PAA biosynthesis, underscoring the importance of Group II GH3 enzymes in balancing IAA/PAA ratios [99]. Moreover, this enzyme group, along with AtGH3.17, has been implicated in the detoxification of the auxinic herbicide 2,4-DB, suggesting their potential application in herbicide resistance strategies [102].
5.2. The Specialized Functions of IAA-Amino Acid Conjugates beyond Their Role as Auxin Stock
Previous studies found that the exogenous addition of IAA-aa and IAA both rapidly increased content-free IAA levels and exhibited similar high growth factor phenotypes, suggesting that IAA-aa is a storage form of IAA [103]. Subsequent studies found that these IAA-aa, IAA-Leu, and IAA-Ala could be reversibly converted to free IAA by the action of the hydrolases, IAA-LEUCINE RESIS TANT1 (ILR1), ILR1-LIKE proteins (ILLs), and IAA-ALANINE RESISTANT3 (IAR3) [5,104,105]. IAA-Glu and IAA-Asp, once considered only as degradation intermediates, are now recognized as reversible storage forms [14]. Beyond storage, IAA-aa have been identified as possessing unique biological functions. IAA-Trp, for instance, acts as a ‘super inactivator’ by not only consuming free IAA for its synthesis but also antagonizing the activity of residual IAA, with IAA-Trp significantly mitigating root inhibition effects caused by IAA [24]. IAA-Asp has been reported to have more diverse roles: (1) correcting the temperature sensitivity of henbane (Hyoscyamus muticus) XIlB2 (temperature-sensitive variant) cells [22]; (2) IAA-Asp directly and specifically enhance the pea (Pisum sativum) responses to abiotic stress by increasing the antioxidant enzyme activity and then reducing the H2O2 concentration [23]; (3) IAA-Asp as a ripening signal in grapes (Vitis vinifera) can be perceived at a certain stage of fruit development; however, the mechanism of sensing remains unknown [21]; and (4) IAA-Asp promotes pathogen development in plants by regulating the transcription of virulence genes [92]. These insights suggest that IAA-amino acids are not just byproducts of GH3 activity but are biologically active molecules with specific roles.
6. Concluding Remarks
Our review underscores that Group II GH3 enzymes and IAA-amino acids are integral components of plant biology, with roles extending beyond the simple conversion of free IAA to its stored form. The functional diversity of these enzymes and their products in plant growth, development, and stress adaptation is a testament to the complexity of plant hormonal regulation. Each GH3 gene exhibits specific spatial and temporal expression patterns, contributing to a nuanced regulatory network. These findings challenge the traditional view of IAA-amino acids as mere storage forms and highlight the need to consider their biological activity in understanding plant physiology and development.
Author Contributions
P.L., T.-T.L. and D.-W.D. drafted and wrote this review; D.-W.D., Q.M. and W.-M.S. edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (Grant No. 32030099) and Special Project on people’s livelihood of Gansu Province (Grant No. 21CX6NA082).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Di, D.W.; Zhang, C.; Luo, P.; An, C.W.; Guo, G.Q. The biosynthesis of auxin: How many paths truly lead to IAA? Plant Growth Regul. 2016, 78, 275–285. [Google Scholar] [CrossRef]
- Fiedler, L.; Friml, J. Rapid auxin signaling: Unknowns old and new. Curr. Opin. Plant Biol. 2023, 75, 102443. [Google Scholar] [CrossRef] [PubMed]
- Jing, H.; Wilkinson, E.G.; Sageman-Furnas, K.; Strader, L.C. Auxin and abiotic stress responses. J. Exp. Bot. 2023, 74, 7000–7014. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Di, D.W. Precise regulation of the TAA1/TAR-YUCCA auxin biosynthesis pathway in plants. Int. J. Mol. Sci. 2023, 24, 8514. [Google Scholar] [CrossRef] [PubMed]
- LeClere, S.; Tellez, R.; Rampey, R.A.; Matsuda, S.P.T.; Bartel, B. Characterization of a family of IAA-amino acid conjugate hydrolases from Arabidopsis. J. Biol. Chem. 2002, 277, 20446–20452. [Google Scholar] [CrossRef]
- Mellor, N.; Band, L.R.; Pencík, A.; Novák, O.; Rashed, A.; Holman, T.; Wilson, M.H.; Voss, U.; Bishopp, A.; King, J.R.; et al. Dynamic regulation of auxin oxidase and conjugating enzymes and modulates auxin homeostasis. Proc. Natl. Acad. Sci. USA 2016, 113, 11022–11027. [Google Scholar] [CrossRef]
- Porco, S.; Pencík, A.; Rashed, A.; Voss, U.; Casanova-Sáez, R.; Bishopp, A.; Golebiowska, A.; Bhosale, R.; Swarup, R.; Swarup, K.; et al. Dioxygenase-encoding gene controls IAA oxidation and homeostasis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2016, 113, 11016–11021. [Google Scholar] [CrossRef]
- Staswick, P.E.; Serban, B.; Rowe, M.; Tiryaki, I.; Maldonado, M.T.; Maldonado, M.C.; Suza, W. Characterization of an Arabidopsis enzyme family that conjugates amino acids to indole-3-acetic acid. Plant Cell 2005, 17, 616–627. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, R.; Ma, C.J.; Vlot, A.C.; Klessig, D.F.; Pichersky, E. Inactive methyl indole-3-acetic acid ester can be hydrolyzed and activated by several esterases belonging to the AMES esterase family of Arabidopsis. Plant Physiol. 2008, 147, 1034–1045. [Google Scholar] [CrossRef]
- Zhang, J.; Lin, J.E.; Harris, C.; Pereira, F.C.M.; Wu, F.; Blakeslee, J.J.; Peer, W.A. DAO1 catalyzes temporal and tissue-specific oxidative inactivation of auxin in. Proc. Natl. Acad. Sci. USA 2016, 113, 11010–11015. [Google Scholar] [CrossRef] [PubMed]
- Casanova-Saez, R.; Mateo-Bonmati, E.; Ljung, K. Auxin Metabolism in Plants. CSH Perspect. Biol. 2021, 13, a039867. [Google Scholar] [CrossRef] [PubMed]
- Ostin, A.; Kowalyczk, M.; Bhalerao, R.P.; Sandberg, G. Metabolism of indole-3-acetic acid in Arabidopsis. Plant Physiol. 1998, 118, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Rampey, R.A.; LeClere, S.; Kowalczyk, M.; Ljung, K.; Sandberg, G.; Bartel, B. A family of auxin-conjugate hydrolases that contributes to free indole-3-acetic acid levels during Arabidopsis germination. Plant Physiol. 2004, 135, 978–988. [Google Scholar] [CrossRef]
- Hayashi, K.I.; Arai, K.; Aoi, Y.; Tanaka, Y.; Hira, H.; Guo, R.; Hu, Y.; Ge, C.; Zhao, Y.; Kasahara, H.; et al. The main oxidative inactivation pathway of the plant hormone auxin. Nat. Commun. 2021, 12, 6752. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Yue, R.; Tao, S.; Yang, Y.; Zhang, L.; Xu, M.; Wang, H.; Shen, C. Genome-wide identification, expression analysis of auxin-responsive GH3 family genes in maize (Zea mays L.) under abiotic stresses. J. Integr. Plant Biol. 2015, 57, 783–795. [Google Scholar] [CrossRef]
- Hagen, G.; Kleinschmidt, A.; Guilfoyle, T. Auxin-regulated gene expression in intact soybean hypocotyl and excised hypocotyl sections. Planta 1984, 162, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Kaur, N.; Tyagi, A.K.; Khurana, J.P. The auxin-responsive GH3 gene family in rice (Oryza sativa). Funct. Integr. Genom. 2006, 6, 36–46. [Google Scholar] [CrossRef]
- Jiang, W.; Yin, J.; Zhang, H.; He, Y.; Shuai, S.; Chen, S.; Cao, S.; Li, W.; Ma, D.; Chen, H. Genome-wide identification, characterization analysis and expression profiling of auxin-responsive GH3 family genes in wheat (Triticum aestivum L.). Mol. Biol. Rep. 2020, 47, 3885–3907. [Google Scholar] [CrossRef]
- Ludwig-Muller, J.; Julke, S.; Bierfreund, N.M.; Decker, E.L.; Reski, R. Moss (Physcomitrella patens) GH3 proteins act in auxin homeostasis. New Phytol. 2009, 181, 323–338. [Google Scholar] [CrossRef]
- Sherp, A.M.; Westfall, C.S.; Alvarez, S.; Jez, J.M. Arabidopsis thaliana GH3.15 acyl acid amido synthetase has a highly specific substrate preference for the auxin precursor indole-3-butyric acid. J. Biol. Chem. 2018, 293, 4277–4288. [Google Scholar] [CrossRef]
- Böttcher, C.; Burbidge, C.A.; Boss, P.K.; Davies, C. Interactions between ethylene and auxin are crucial to the control of grape (Vitis vinifera L.) berry ripening. BMC Plant Biol. 2013, 13, 222. [Google Scholar] [CrossRef]
- Oetiker, J.H.; Aeschbacher, G. Temperature-sensitive plant cells with shunted indole-3-acetic acid conjugation. Plant Physiol. 1997, 114, 1385–1395. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, M.; Ciarkowska, A.; Jakubowska, A. The auxin conjugate indole-3-acetyl-aspartate affects responses to cadmium and salt stress in Pisum sativum L. J. Plant Physiol. 2016, 191, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Staswick, P.E. The tryptophan conjugates of jasmonic and indole-3-acetic acids are endogenous auxin inhibitors. Plant Physiol. 2009, 150, 1310–1321. [Google Scholar] [CrossRef]
- Chen, Q.; Westfall, C.S.; Hicks, L.M.; Wang, S.; Jez, J.M. Kinetic basis for the conjugation of auxin by a GH3 family indole-acetic acid-amido synthetase. J. Biol. Chem. 2010, 285, 29780–29786. [Google Scholar] [CrossRef]
- Xu, G.L.; Zhang, Y.K.; Li, M.Y.; Jiao, X.; Zhou, L.; Ming, Z.H. Crystal structure of the acyl acid amido synthetase GH3-8 from Oryza sativa. Biochem. Biophys. Res. Commun. 2021, 534, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Takase, T.; Nakazawa, M.; Ishikawa, A.; Manabe, K.; Matsui, M. DFL2, a new member of the Arabidopsis GH3 gene family, is involved in red light-specific hypocotyl elongation. Plant Cell Physiol. 2003, 44, 1071–1080. [Google Scholar] [CrossRef]
- Peat, T.S.; Bottcher, C.; Newman, J.; Lucent, D.; Cowieson, N.; Davies, C. Crystal structure of an indole-3-acetic acid amido synthetase from grapevine involved in auxin homeostasis. Plant Cell 2012, 24, 4525–4538. [Google Scholar] [CrossRef]
- Westfall, C.S.; Zubieta, C.; Herrmann, J.; Kapp, U.; Nanao, M.H.; Jez, J.M. Structural basis for prereceptor modulation of plant hormones by GH3 proteins. Science 2012, 336, 1708–1711. [Google Scholar] [CrossRef]
- Bottcher, C.; Dennis, E.G.; Booker, G.W.; Polyak, S.W.; Boss, P.K.; Davies, C. A novel tool for studying auxin-metabolism: The inhibition of grapevine indole-3-acetic acid-amido synthetases by a reaction intermediate analogue. PLoS ONE 2012, 7, e37632. [Google Scholar] [CrossRef]
- Xie, Y.; Zhu, Y.; Wang, N.; Luo, M.; Ota, T.; Guo, R.; Takahashi, I.; Yu, Z.; Aizezi, Y.; Zhang, L.; et al. Chemical genetic screening identifies nalacin as an inhibitor of GH3 amido synthetase for auxin conjugation. Proc. Natl. Acad. Sci. USA 2022, 119, e2209256119. [Google Scholar] [CrossRef]
- Fukui, K.; Arai, K.; Tanaka, Y.; Aoi, Y.; Kukshal, V.; Jez, J.M.; Kubes, M.F.; Napier, R.; Zhao, Y.; Kasahara, H.; et al. Chemical inhibition of the auxin inactivation pathway uncovers the roles of metabolic turnover in auxin homeostasis. Proc. Natl. Acad. Sci. USA 2022, 119, e2206869119. [Google Scholar] [CrossRef] [PubMed]
- Koochak, H.; Ludwig-Muller, J. Physcomitrium patens mutants in auxin conjugating GH3 proteins show salt stress tolerance but auxin homeostasis is not involved in regulation of oxidative stress factors. Plants 2021, 10, 10071398. [Google Scholar] [CrossRef] [PubMed]
- Mittag, J.; Gabrielyan, A.; Ludwig-Müller, J. Knockout of GH3 genes in the moss Physcomitrella patens leads to increased IAA levels at elevated temperature and in darkness. Plant Physiol. Biochem. 2015, 97, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Ma, L.L.; Huang, H.A.; Ke, S.W.; Gui, C.S.; Ning, X.Y.; Zhang, X.Q.; Zhong, T.X.; Xie, X.M.; Chen, S. Overexpression of SgGH3.1 from fine-stem stylo (Stylosanthes guianensis var. intermedia) enhances chilling and cold tolerance in Arabidopsis thaliana. Genes 2021, 12, 1367. [Google Scholar] [CrossRef] [PubMed]
- Cano, A.; Sanchez-Garcia, A.B.; Albacete, A.; Gonzalez-Bayon, R.; Justamante, M.S.; Ibanez, S.; Acosta, M.; Perez-Perez, J.M. Enhanced conjugation of auxin by GH3 enzymes leads to poor adventitious rooting in carnation stem cuttings. Front. Plant Sci. 2018, 9, 566. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Wang, Z.; Kong, X.; Chen, Y.; Li, J. Exogenous tryptophan application improves cadmium tolerance and inhibits cadmium upward transport in broccoli (Brassica oleracea var. italica). Front. Plant Sci. 2022, 13, 969675. [Google Scholar] [CrossRef]
- Ostrowski, M.; Świdziński, M.; Ciarkowska, A.; Jakubowska, A. IAA-amido synthetase activity and GH3 expression during development of pea seedlings. Acta Physiol. Plant 2014, 36, 3029–3037. [Google Scholar] [CrossRef]
- Dong, C.J.; Liu, X.Y.; Xie, L.L.; Wang, L.L.; Shang, Q.M. Salicylic acid regulates adventitious root formation via competitive inhibition of the auxin conjugation enzyme CsGH3.5 in cucumber hypocotyls. Planta 2020, 252, 75. [Google Scholar] [CrossRef]
- Liu, K.D.; Kang, B.C.; Jiang, H.; Moore, S.L.; Li, H.X.; Watkins, C.B.; Setter, T.L.; Jahn, M.M. A GH3-like gene, CcGH3, isolated from Capsicum chinense L. fruit is regulated by auxin and ethylene. Plant Mol. Biol. 2005, 58, 447–464. [Google Scholar] [CrossRef]
- Sun, M.H.; Li, H.; Li, Y.B.; Xiang, H.Z.; Liu, Y.D.; He, Y.; Qi, M.F.; Li, T.L. Tomato YABBY2b controls plant height through regulating indole-3-acetic acid-amido synthetase (GH3.8) expression. Plant Sci. 2020, 297, 110530. [Google Scholar] [CrossRef]
- Sravankumar, T.; Akash; Naik, N.; Kumar, R. A ripening-induced SlGH3-2 gene regulates fruit ripening via adjusting auxin-ethylene levels in tomato (Solanum lycopersicum L.). Plant Mol. Biol. 2018, 98, 455–469. [Google Scholar] [CrossRef]
- Ai, G.; Huang, R.; Zhang, D.; Li, M.; Li, G.; Li, W.; Ahiakpa, J.K.; Wang, Y.; Hong, Z.; Zhang, J. SlGH3.15, a member of the GH3 gene family, regulates lateral root development and gravitropism response by modulating auxin homeostasis in tomato. Plant Sci. 2023, 330, 111638. [Google Scholar] [CrossRef]
- Ayil-Gutierrez, B.; Galaz-Avalos, R.; Pena-Cabrera, E.; Loyola-Vargas, V. Dynamics of the concentration of IAA and some of its conjugates during the induction of somatic embryogenesis in Coffea canephora. Plant Signal. Behav. 2013, 8, e26998. [Google Scholar] [CrossRef]
- Mendez-Hernandez, H.A.; Quintana-Escobar, A.O.; Uc-Chuc, M.A.; Loyola-Vargas, V.M. Genome-wide analysis, modeling, and identification of amino acid binding motifs suggest the involvement of GH3 genes during somatic embryogenesis of Coffea canephora. Plants 2021, 10, 2034. [Google Scholar] [CrossRef]
- Han, Q.; Chen, K.; Yan, D.; Hao, G.; Qi, J.; Wang, C.; Dirk, L.M.A.; Bruce Downie, A.; Gong, J.; Wang, J.; et al. ZmDREB2A regulates ZmGH3.2 and ZmRAFS, shifting metabolism towards seed aging tolerance over seedling growth. Plant J. 2020, 104, 268–282. [Google Scholar] [CrossRef]
- Zou, X.; Long, J.; Zhao, K.; Peng, A.; Chen, M.; Long, Q.; He, Y.; Chen, S. Overexpressing GH3.1 and GH3.1L reduces susceptibility to Xanthomonas citri subsp. citri by repressing auxin signaling in citrus (Citrus sinensis Osbeck). PLoS ONE 2019, 14, e0220017. [Google Scholar] [CrossRef]
- Zhao, D.; Wang, Y.; Feng, C.; Wei, Y.; Peng, X.; Guo, X.; Guo, X.; Zhai, Z.; Li, J.; Shen, X.; et al. Overexpression of MsGH3.5 inhibits shoot and root development through the auxin and cytokinin pathways in apple plants. Plant J. 2020, 103, 166–183. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.G.; Wang, N.; Wang, D.H.; Cheng, L.; Wang, Y.X.; Zhao, Y.W.; Ding, J.Y.; Gu, K.D.; Xiao, X.; Hao, Y.J. A basic/helix-loop-helix transcription factor controls leaf shape by regulating auxin signaling in apple. New Phytol. 2020, 228, 1897–1913. [Google Scholar] [CrossRef] [PubMed]
- Vielba, J.M.; VERico, S.; Covelo, P.; Vidal, N.; Sánchez, C. Expression of a GH3 gene during adventitious rooting in Chestnut. In Proceedings of the 4th International Conference of the IUFRO Unit 2.09.02 on “Development and Application of Vegetative Propagation Technologies in Plantation Forestry to Cope with a Changing Climate and Environment”, La Plata, Argentina, 19–23 September 2016. [Google Scholar]
- Xu, D.; Yang, Y.; Tao, S.; Wang, Y.; Yuan, H.; Sharma, A.; Wang, X.; Shen, C.; Yan, D.; Zheng, B. Identification and expression analysis of auxin-responsive GH3 family genes in Chinese hickory (Carya cathayensis) during grafting. Mol. Biol. Rep. 2020, 47, 4495–4506. [Google Scholar] [CrossRef] [PubMed]
- Brunoni, F.; Collani, S.; Casanova-Saez, R.; Simura, J.; Karady, M.; Schmid, M.; Ljung, K.; Bellini, C. Conifers exhibit a characteristic inactivation of auxin to maintain tissue homeostasis. New Phytol. 2020, 226, 1753–1765. [Google Scholar] [CrossRef]
- Zhang, R.S.; Wang, Y.C.; Wang, C.; Wei, Z.G.; Xia, D.; Wang, Y.F.; Liu, G.F.; Yang, C.P. Time-course analysis of levels of indole-3-acetic acid and expression of auxin-responsive GH3 genes in Betula platyphylla. Plant Mol. Biol. Rep. 2011, 29, 898–905. [Google Scholar] [CrossRef]
- Takase, T.; Nakazawa, M.; Ishikawa, A.; Kawashima, M.; Ichikawa, T.; Takahashi, N.; Shimada, H.; Manabe, K.; Matsui, M. ydk1-D, an auxin-responsive GH3 mutant that is involved in hypocotyl and root elongation. Plant J. 2004, 37, 471–483. [Google Scholar] [CrossRef]
- Nakazawa, M.; Yabe, N.; Ichikawa, T.; Yamamoto, Y.Y.; Yoshizumi, T.; Hasunuma, K.; Matsui, M. DFL2, an auxin-responsive GH3 gene homologue, negatively regulates shoot cell elongation and lateral root formation, and positively regulates the light response of hypocotyl length. Plant J. 2001, 25, 213–221. [Google Scholar]
- Khan, S.; Stone, J.M. Arabidopsis thaliana GH3.9 in auxin and jasmonate cross talk. Plant Signal Behav. 2007, 2, 483–485. [Google Scholar] [CrossRef]
- Di, D.W.; Zhang, C.G.; Guo, G.Q. Involvement of secondary messengers and small organic molecules in auxin perception and signaling. Plant Cell Rep. 2015, 34, 895–904. [Google Scholar] [CrossRef]
- Tian, C.; Muto, H.; Higuchi, K.; Matamura, T.; Tatematsu, K.; Koshiba, T.; Yamamoto, K.T. Disruption and overexpression of auxin response factor 8 gene of Arabidopsis affect hypocotyl elongation and root growth habit, indicating its possible involvement in auxin homeostasis in light condition. Plant J. 2004, 40, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Mallory, A.C.; Bartel, D.P.; Bartel, B. MicroRNA-directed regulation of Arabidopsis is essential for proper development and modulates expression of early auxin response genes. Plant Cell 2005, 17, 1360–1375. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, L.; Mongelard, G.; Flokova, K.; Pacurar, D.I.; Novak, O.; Staswick, P.; Kowalczyk, M.; Pacurar, M.; Demailly, H.; Geiss, G.; et al. Auxin controls Arabidopsis adventitious root initiation by regulating jasmonic acid homeostasis. Plant Cell 2012, 24, 2515–2527. [Google Scholar] [CrossRef] [PubMed]
- Shin, R.; Burch, A.Y.; Huppert, K.A.; Tiwari, S.B.; Murphy, A.S.; Guilfoyle, T.J.; Schachtman, D.P. The Arabidopsis transcription factor MYB77 modulates auxin signal transduction. Plant Cell 2007, 19, 2440–2453. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.Y.; Xue, H.W. PI4Kγ2 interacts with E3 ligase MIEL1 to regulate auxin metabolism and root development. Plant Physiol. 2020, 184, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Low, P.M.; Lim, A.R.Q.; Yang, Y.; Yuan, L.; Ma, W.; Kong, Q.; Low, P.M.; Lim, A.R.Q.; Yang, Y.; et al. Functional antagonism of WRI1 and TCP20 modulates GH3.3 expression to maintain auxin homeostasis in roots. Plants 2022, 11, 454. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Ma, W.; Yang, H.; Ma, G.; Mantyla, J.J.; Benning, C. The Arabidopsis WRINKLED1 transcription factor affects auxin homeostasis in roots. J. Exp. Bot. 2017, 68, 4627–4634. [Google Scholar] [CrossRef] [PubMed]
- Weiste, C.; Dröge-Laser, W. The Arabidopsis transcription factor bZIP11 activates auxin-mediated transcription by recruiting the histone acetylation machinery. Nat. Commun. 2014, 5, 3883. [Google Scholar] [CrossRef] [PubMed]
- Di Mambro, R.; De Ruvo, M.; Pacifici, E.; Salvi, E.; Sozzani, R.; Benfey, P.N.; Busch, W.; Novak, O.; Ljung, K.; Di Paola, L.; et al. Auxin minimum triggers the developmental switch from cell division to cell differentiation in the Arabidopsis root. Proc. Natl. Acad. Sci. USA 2017, 114, E7641–E7649. [Google Scholar] [CrossRef] [PubMed]
- Ohto, M.A.; Hayashi, S.; Sawa, S.; Hashimoto-Ohta, A.; Nakamura, K. Involvement of HLS1 in sugar and auxin signaling in Arabidopsis leaves. Plant Cell Physiol. 2006, 47, 1603–1611. [Google Scholar] [CrossRef] [PubMed]
- Sohlberg, J.J.; Myrenas, M.; Kuusk, S.; Lagercrantz, U.; Kowalczyk, M.; Sandberg, G.; Sundberg, E. STY1 regulates auxin homeostasis and affects apical-basal patterning of the Arabidopsis gynoecium. Plant J. 2006, 47, 112–123. [Google Scholar] [CrossRef]
- Sun, Y.; Yang, Y.; Yuan, Z.; Ludwig-Müller, J.; Yu, C.; Xu, Y.F.; Shao, X.H.; Li, X.F.; Decker, E.L.; Reski, R.; et al. Overexpression of the Arabidopsis gene UPRIGHT ROSETTE reveals a homeostatic control for indole-3-acetic acid. Plant Physiol. 2010, 153, 1311–1320. [Google Scholar] [CrossRef]
- Dai, Z.; Wang, J.; Yang, X.; Lu, H.; Miao, X.; Shi, Z. Modulation of plant architecture by the miR156f-OsSPL7-OsGH3.8 pathway in rice. J. Exp. Bot. 2018, 69, 5117–5130. [Google Scholar] [CrossRef]
- Domingo, C.; Andrés, F.; Tharreau, D.; Iglesias, D.J.; Talón, M. Constitutive Expression of OsGH3.1 Reduces Auxin Content and Enhances Defense Response and Resistance to a Fungal Pathogen in Rice. Mol. Plant-Microbe Interact. 2009, 22, 201–210. [Google Scholar] [CrossRef]
- Du, H.; Wu, N.; Fu, J.; Wang, S.P.; Li, X.H.; Xiao, J.H.; Xiong, L.Z. A GH3 family member, OsGH3-2, modulates auxin and abscisic acid levels and differentially affects drought and cold tolerance in rice. J. Exp. Bot. 2012, 63, 6467–6480. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.N.; Wang, S.K.; Xu, Y.X.; Yu, C.L.; Shen, C.J.; Qian, Q.; Geisler, M.; Jiang, D.A.; Qi, Y.H. The auxin response factor, OsARF19, controls rice leaf angles through positively regulating OsGH3-5 and OsBRI1. Plant Cell Environ. 2015, 38, 638–654. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.W.; Li, C.H.; Cao, J.; Zhang, Y.C.; Zhang, S.Q.; Xia, Y.F.; Sun, D.Y.; Sun, Y. Altered architecture and enhanced drought tolerance in rice via the down-regulation of indole-3-acetic acid by TLD1/OsGH3.13 activation. Plant Physiol. 2009, 151, 1889–1901. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.H.; Lin, X.H.; Zuo, Y.; Yu, Z.L.; Baerson, S.R.; Pan, Z.Q.; Zeng, R.S.; Song, Y.Y. Transcription factor OsbZIP49 controls tiller angle and plant architecture through the induction of indole-3-acetic acid-amido synthetases in rice. Plant J. 2021, 108, 1346–1364. [Google Scholar] [CrossRef]
- Yang, J.H.; Han, S.J.; Yoon, E.K.; Lee, W.S. Evidence of an auxin signal pathway, microRNA167-ARF8-GH3, and its response to exogenous auxin in cultured rice cells. Nucleic Acids Res. 2006, 34, 1892–1899. [Google Scholar] [CrossRef]
- Yadav, S.R.; Khanday, I.; Majhi, B.B.; Veluthambi, K.; Vijayraghavan, U. Auxin-responsive OsMGH3, a common downstream target of OsMADS1 and OsMADS6, controls rice floret fertility. Plant Cell Physiol. 2011, 52, 2123–2135. [Google Scholar] [CrossRef]
- Luo, P.; Di, D.; Wu, L.; Yang, J.; Lu, Y.; Shi, W. MicroRNAs are involved in regulating plant development and stress response through fine-tuning of TIR1/AFB-dependent auxin signaling. Int. J. Mol. Sci. 2022, 23, 510. [Google Scholar] [CrossRef]
- Park, J.E.; Park, J.Y.; Kim, Y.S.; Staswick, P.E.; Jeon, J.; Yun, J.; Kim, S.Y.; Kim, J.; Lee, Y.H.; Park, C.M. GH3-mediated auxin homeostasis links growth regulation with stress adaptation response in Arabidopsis. J. Biol. Chem. 2007, 282, 10036–10046. [Google Scholar] [CrossRef] [PubMed]
- Seo, P.J.; Xiang, F.; Qiao, M.; Park, J.Y.; Lee, Y.N.; Kim, S.G.; Lee, Y.H.; Park, W.J.; Park, C.M. The MYB96 transcription factor mediates abscisic acid signaling during drought stress response in Arabidopsis. Plant Physiol. 2009, 151, 275–289. [Google Scholar] [CrossRef]
- Casanova-Saez, R.; Mateo-Bonmati, E.; Simura, J.; Pencik, A.; Novak, O.; Staswick, P.; Ljung, K. Inactivation of the entire Arabidopsis group II GH3s confers tolerance to salinity and water deficit. New Phytol. 2022, 235, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Seo, P.J.; Lee, A.K.; Jung, J.H.; Kim, Y.S.; Park, C.M. An Arabidopsis GH3 gene, encoding an auxin-conjugating enzyme, mediates phytochrome B-regulated light signals in hypocotyl growth. Plant Cell Physiol. 2007, 48, 1236–1241. [Google Scholar] [CrossRef]
- Han, S.; Jia, M.Z.; Yang, J.F.; Jiang, J. The integration of ACS2-generated ACC with GH3-mediated IAA homeostasis in NaCl-stressed primary root elongation of Arabidopsis seedlings. Plant Growth Regul. 2019, 88, 151–158. [Google Scholar] [CrossRef]
- Ding, Z.J.; Yan, J.Y.; Li, C.X.; Li, G.X.; Wu, Y.R.; Zheng, S.J. Transcription factor WRKY46 modulates the development of Arabidopsis lateral roots in osmotic/salt stress conditions via regulation of ABA signaling and auxin homeostasis. Plant J. 2015, 84, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Fang, Y.; Wang, S.; He, K. The alleviation of ammonium toxicity in plants. J. Integr. Plant Biol. 2023, 65, 1362–1368. [Google Scholar] [CrossRef] [PubMed]
- Di, D.W. New molecular mechanisms of plant response to ammonium nutrition. Appl. Sci. 2023, 13, 11570. [Google Scholar] [CrossRef]
- Di, D.W.; Li, G.J.; Sun, L.; Wu, J.J.; Wang, M.; Kronzucker, H.J.; Fang, S.; Chu, J.F.; Shi, W.M. High ammonium inhibits root growth in Arabidopsis thaliana by promoting auxin conjugation rather than inhibiting auxin biosynthesis. J. Plant Physiol. 2021, 261, 153415. [Google Scholar] [CrossRef]
- Di, D.W.; Sun, L.; Zhang, X.N.; Li, G.J.; Kronzucker, H.J.; Shi, W.M. Involvement of auxin in the regulation of ammonium tolerance in rice (Oryza sativa L.). Plant Soil. 2018, 432, 373–387. [Google Scholar] [CrossRef]
- Dziewit, K.; Pencik, A.; Dobrzynska, K.; Novak, O.; Szal, B.; Podgorska, A. Spatiotemporal auxin distribution in Arabidopsis tissues is regulated by anabolic and catabolic reactions under long-term ammonium stress. BMC Plant Biol. 2021, 21, 602. [Google Scholar] [CrossRef]
- Di, D.W.; Sun, L.; Wang, M.; Wu, J.; Kronzucker, H.J.; Fang, S.; Chu, J.; Shi, W.; Li, G. WRKY46 promotes ammonium tolerance in Arabidopsis by repressing NUDX9 and indole-3-acetic acid-conjugating genes and by inhibiting ammonium efflux in the root elongation zone. New Phytol. 2021, 232, 190–207. [Google Scholar] [CrossRef]
- Zhang, Z.Q.; Li, Q.; Li, Z.M.; Staswick, P.E.; Wang, M.Y.; Zhu, Y.; He, Z.H. Dual regulation role of GH3.5 in salicylic acid and auxin signaling during Arabidopsis-Pseudomonas syringae interaction. Plant Physiol. 2007, 145, 450–464. [Google Scholar] [CrossRef]
- González-Lamothe, R.; El Oirdi, M.; Brisson, N.; Bouarab, K. The conjugated auxin indole-3-acetic acid-aspartic acid promotes plant disease development. Plant Cell 2012, 24, 762–777. [Google Scholar] [CrossRef]
- Fu, J.; Liu, H.B.; Li, Y.; Yu, H.H.; Li, X.H.; Xiao, J.H.; Wang, S.P. Manipulating broad-spectrum disease resistance by suppressing pathogen-induced auxin accumulation in rice. Plant Physiol. 2011, 155, 589–602. [Google Scholar] [CrossRef]
- Ding, X.; Cao, Y.; Huang, L.; Zhao, J.; Xu, C.; Li, X.; Wang, S. Activation of the indole-3-acetic acid-amido synthetase GH3-8 suppresses expansin expression and promotes salicylate- and jasmonate-independent basal immunity in rice. Plant Cell 2008, 20, 228–240. [Google Scholar] [CrossRef]
- Jez, J.M. Connecting primary and specialized metabolism: Amino acid conjugation of phytohormones by GRETCHEN HAGEN 3 (GH3) acyl acid amido synthetases. Curr. Opin. Plant Biol. 2022, 66, 102194. [Google Scholar] [CrossRef]
- Zolman, B.K.; Martinez, N.; Millius, A.; Adham, A.R.; Bartel, B. Identification and characterization of Arabidopsis indole-3-butyric acid response mutants defective in novel peroxisomal enzymes. Genetics 2008, 180, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, M.; Ciarkowska, A. Pea GH3 acyl acid amidosynthetase conjugates IAA to proteins in immature seeds of Pisum sativum L.—A new perspective on formation of high-molecular weight conjugates of auxin. J. Plant Physiol. 2021, 256, 153312. [Google Scholar] [CrossRef] [PubMed]
- Seidel, C.; Walz, A.; Park, S.; Cohen, J.D.; Ludwig-Muller, J. Indole-3-acetic acid protein conjugates: Novel players in auxin homeostasis. Plant Biol. 2006, 8, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Aoi, Y.; Tanaka, K.; Cook, S.D.; Hayashi, K.I.; Kasahara, H. GH3 auxin-amido synthetases alter the ratio of indole-3-acetic acid and phenylacetic acid in Arabidopsis. Plant Cell Physiol. 2020, 61, 596–605. [Google Scholar] [CrossRef]
- Mashiguchi, K.; Hisano, H.; Takeda-Kamiya, N.; Takebayashi, Y.; Ariizumi, T.; Gao, Y.; Ezura, H.; Sato, K.; Zhao, Y.; Hayashi, K.I.; et al. Agrobacterium tumefaciens enhances biosynthesis of two distinct auxins in the formation of crown galls. Plant Cell Physiol. 2019, 60, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, S.; Mashiguchi, K.; Tanaka, K.; Hishiyama, S.; Sakai, T.; Hanada, K.; Kinoshita-Tsujimura, K.; Yu, H.; Dai, X.; Takebayashi, Y.; et al. Distinct characteristics of indole-3-acetic acid and phenylacetic acid, two common auxins in plants. Plant Cell Physiol. 2015, 56, 1641–1654. [Google Scholar] [CrossRef]
- Sherp, A.M.; Lee, S.G.; Schraft, E.; Jez, J.M. Modification of auxinic phenoxyalkanoic acid herbicides by the acyl acid amido synthetase GH3.15 from Arabidopsis. J. Biol. Chem. 2018, 293, 17731–17738. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; De Gernier, H.; Duan, X.L.; Xie, Y.M.; Geelen, D.; Hayashi, K.; Xuan, W.; Geisler, M.; ten Tusscher, K.; Beeckman, T.; et al. GH3-mediated auxin inactivation attenuates multiple stages of lateral root development. New Phytol. 2023, 240, 1900–1912. [Google Scholar] [CrossRef] [PubMed]
- Bartel, B.; Fink, G.R. Ilr1, an amidohydrolase that releases active indole-3-acetic-acid from conjugates. Science 1995, 268, 1745–1748. [Google Scholar] [CrossRef] [PubMed]
- Davies, R.T.; Goetz, D.H.; Lasswell, J.; Anderson, M.N.; Bartel, B. IAR3 encodes an auxin conjugate hydrolase from Arabidopsis. Plant Cell 1999, 11, 365–376. [Google Scholar] [CrossRef]
Figure 1.
Catalytic reaction and inhibitor structures of Group II GH3 amido synthetase. (A) Schematic representation of the total reaction mediated by Group II GH3 amido synthetases. (B) Chemical structures of inhibitors targeting Group II GH3 amido synthetases: AIEP (adenosine-59-[2-(1H-indol-3-yl) ethyl] phosphate), KKI (kakeimide), and nalacin (N-[4-[[6-(1H-pyrazol-1-yl)-3-pyridazinyl] amino] phenyl]-3-(trifluoromethyl)benzamide).
Figure 1.
Catalytic reaction and inhibitor structures of Group II GH3 amido synthetase. (A) Schematic representation of the total reaction mediated by Group II GH3 amido synthetases. (B) Chemical structures of inhibitors targeting Group II GH3 amido synthetases: AIEP (adenosine-59-[2-(1H-indol-3-yl) ethyl] phosphate), KKI (kakeimide), and nalacin (N-[4-[[6-(1H-pyrazol-1-yl)-3-pyridazinyl] amino] phenyl]-3-(trifluoromethyl)benzamide).

Figure 2.
Regulatory network of Arabidopsis GH3-dependent IAA conjugation in abiotic and biotic stress responses. Abbreviation: NH4+, ammonium; WRKY46, WRKY DNA-BINDING PROTEIN46; bZIP11, basic leucine Zipper11; ARF6/7/8/17, auxin response factor6/7/8/17; MYB30/77/96, myeloblastosis30/77/96; WRI1, WRINKLED1; TCP20, TEOSINTE BRANCHED 1; CYCLOIDEA, PCF (TCP)-DOMAIN FAMILY PROTEIN 20; ARR1, Arabidopsis response regulator1; miR160, microRNA; IAA, indole-3-acetic acid; ABA, abscisic acid; JA, jasmonic acid; ET, ethylene; SA, salicylic acid; CK, cytokinin.
Figure 2.
Regulatory network of Arabidopsis GH3-dependent IAA conjugation in abiotic and biotic stress responses. Abbreviation: NH4+, ammonium; WRKY46, WRKY DNA-BINDING PROTEIN46; bZIP11, basic leucine Zipper11; ARF6/7/8/17, auxin response factor6/7/8/17; MYB30/77/96, myeloblastosis30/77/96; WRI1, WRINKLED1; TCP20, TEOSINTE BRANCHED 1; CYCLOIDEA, PCF (TCP)-DOMAIN FAMILY PROTEIN 20; ARR1, Arabidopsis response regulator1; miR160, microRNA; IAA, indole-3-acetic acid; ABA, abscisic acid; JA, jasmonic acid; ET, ethylene; SA, salicylic acid; CK, cytokinin.
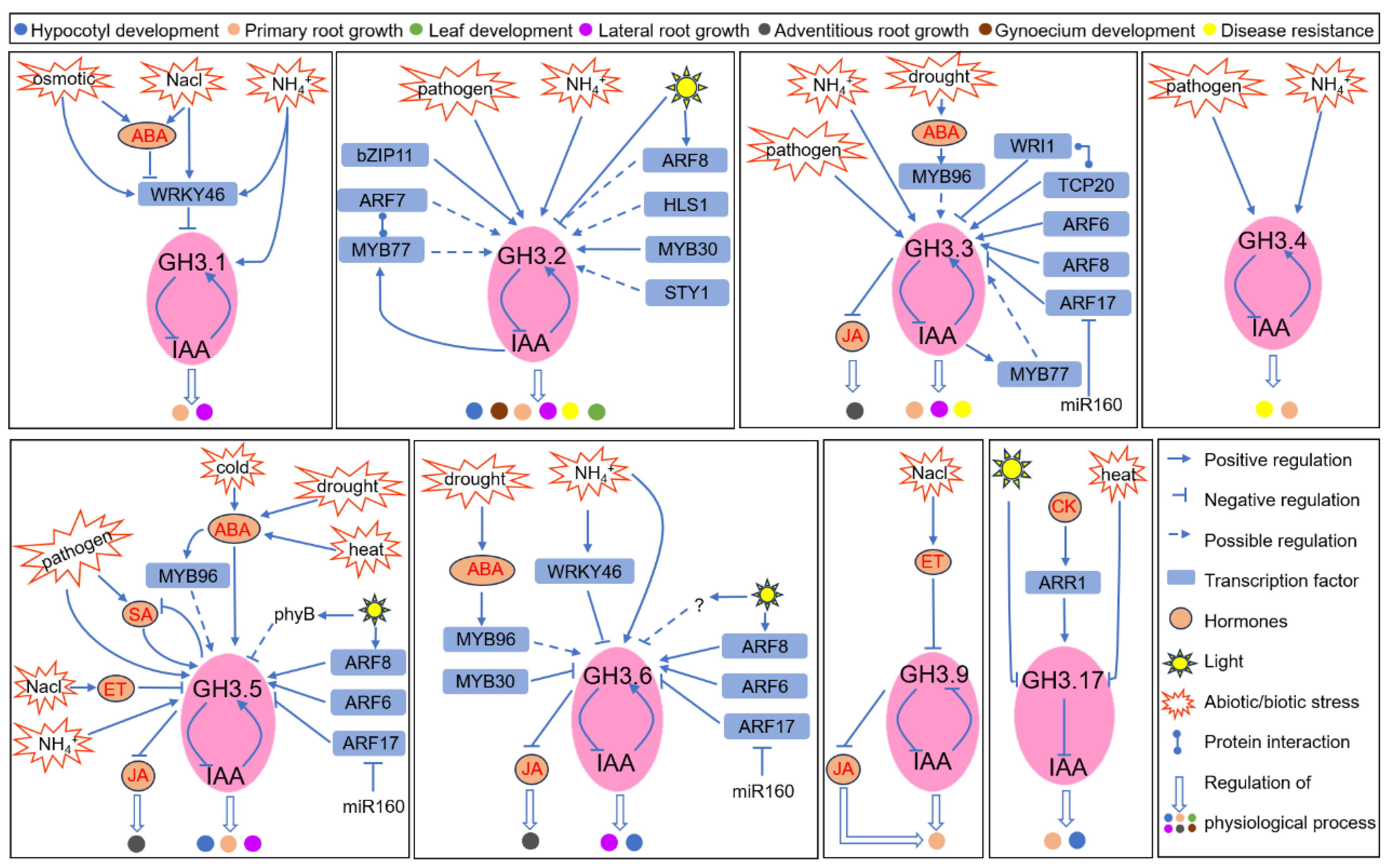
Figure 3.
Role of rice GH3-dependent IAA conjugation in response to abiotic and biotic stresses and its transcriptional regulation. Abbreviation: bZIP49, basic leucine Zipper49; ARF8/19, auxin response factor; MADS1/6, MADS-domain transcription factor1/6; SPL7, SQUAMOSA PROMOTER BINDING PROTEIN-LIKE 7; miR156f/167, microRNA156f/167; IAA, indole-3-acetic acidl; BR, brassinolide; ABA, abscisic acid.
Figure 3.
Role of rice GH3-dependent IAA conjugation in response to abiotic and biotic stresses and its transcriptional regulation. Abbreviation: bZIP49, basic leucine Zipper49; ARF8/19, auxin response factor; MADS1/6, MADS-domain transcription factor1/6; SPL7, SQUAMOSA PROMOTER BINDING PROTEIN-LIKE 7; miR156f/167, microRNA156f/167; IAA, indole-3-acetic acidl; BR, brassinolide; ABA, abscisic acid.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Compilation of Group II GH3 genes across various species and their identified biological roles.
Table 1.
Compilation of Group II GH3 genes across various species and their identified biological roles.
| Species | Members | TFs | Biological Process | Ref. |
|---|---|---|---|---|
| Physcomitrella patens | PpGH3.1 | High temperature and salt tolerance | [33,34] | |
| PpGH3.2 | ||||
| Stylosanthes guianensis | SgGH3.1 | Chilling and cold tolerance | [35] | |
| Dianthus caryophyllus | DcGH3.1 | Adventitious root development | [36] | |
| Brassica oleracea | BoGH3.12 | Cadmium tolerance | [37] | |
| Pisum sativum | PsGH3.5 | Seedlings development | [38] | |
| Cucumis sativus | CsGH3.5 | Adventitious root formation | [39] | |
| Capsicum chinense | CcGH3 | Fruit ripening | [40] | |
| Solanum lycopersicum | SlGH3.8 | YABBY2b | Plant height | [41] |
| SlGH3.2 | Fruit ripening | [42] | ||
| SlGH3.15 | Lateral root development; gravitropism | [43] | ||
| Coffea canephora | CcGH3.1 | Somatic embryogenesis | [44,45] | |
| CcGH3.6 | ||||
| CcGH3.17 | ||||
| Zea mays | ZmGH3.2 | DREB2A | Seed aging tolerance | [46] |
| Vitis vinifera | VvGH3.1 | Berry ripening | [30] | |
| VvGH3.2 | [21] | |||
| VvGH3.6 | Tissue auxin homeostasis | [28,30] | ||
| Citrus sinensis | CsGH3.1 | Susceptibility to pathogen | [47] | |
| CsGH3.1L | ||||
| Malus sieversii | MsGH3.5 | RR1a | Shoot and root development | [48] |
| Malus domestica | MdGH3-2 | bHLH3 | Leaf shape | [49] |
| Castanea sativa | CsGH3.1 | Adventitious root development | [50] | |
| Carya cathayensis | CcGH3 | Grafting | [51] | |
| Picea abies | PaGH3.gII.8 | Tissue auxin homeostasis | [52] | |
| PaGH3.gII.9 | ||||
| PaGH3.17 | ||||
| Betula platyphylla | BpGH3.3 | Tissue auxin homeostasis | [53] | |
| BpGH3.5a | ||||
| BpGH3.5b | ||||
| BpGH3.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Luo, P.; Li, T.-T.; Shi, W.-M.; Ma, Q.; Di, D.-W. The Roles of GRETCHEN HAGEN3 (GH3)-Dependent Auxin Conjugation in the Regulation of Plant Development and Stress Adaptation. Plants 2023, 12, 4111. https://doi.org/10.3390/plants12244111
AMA Style
Luo P, Li T-T, Shi W-M, Ma Q, Di D-W. The Roles of GRETCHEN HAGEN3 (GH3)-Dependent Auxin Conjugation in the Regulation of Plant Development and Stress Adaptation. Plants. 2023; 12(24):4111. https://doi.org/10.3390/plants12244111
Chicago/Turabian StyleLuo, Pan, Ting-Ting Li, Wei-Ming Shi, Qi Ma, and Dong-Wei Di. 2023. "The Roles of GRETCHEN HAGEN3 (GH3)-Dependent Auxin Conjugation in the Regulation of Plant Development and Stress Adaptation" Plants 12, no. 24: 4111. https://doi.org/10.3390/plants12244111
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.