
Spatial Distribution and Ecological Determinants of Coexisting Hybrid Oak Species: A Study in Yushan’s Mixed Forest

Abstract
:1. Introduction
2. Results
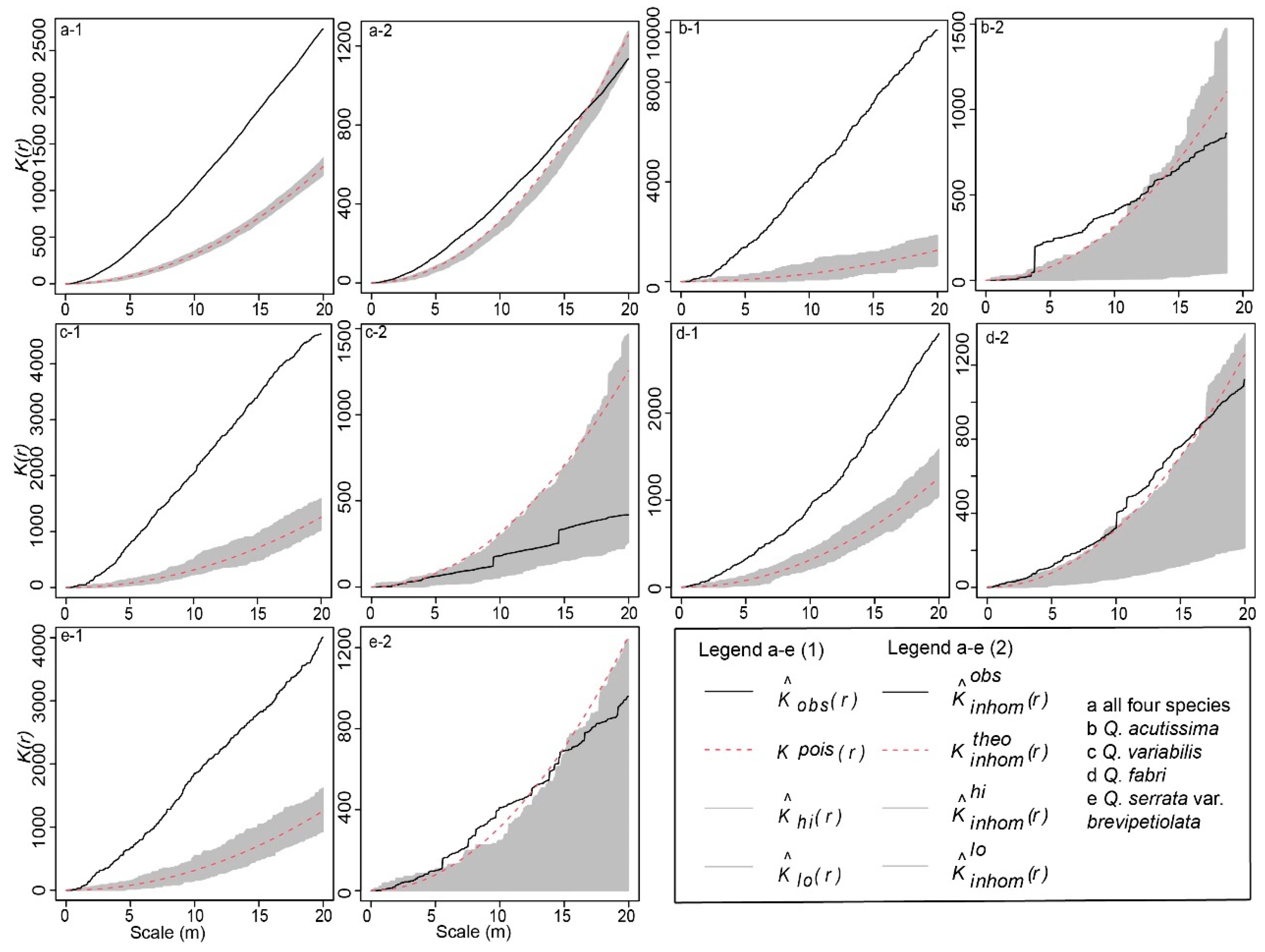
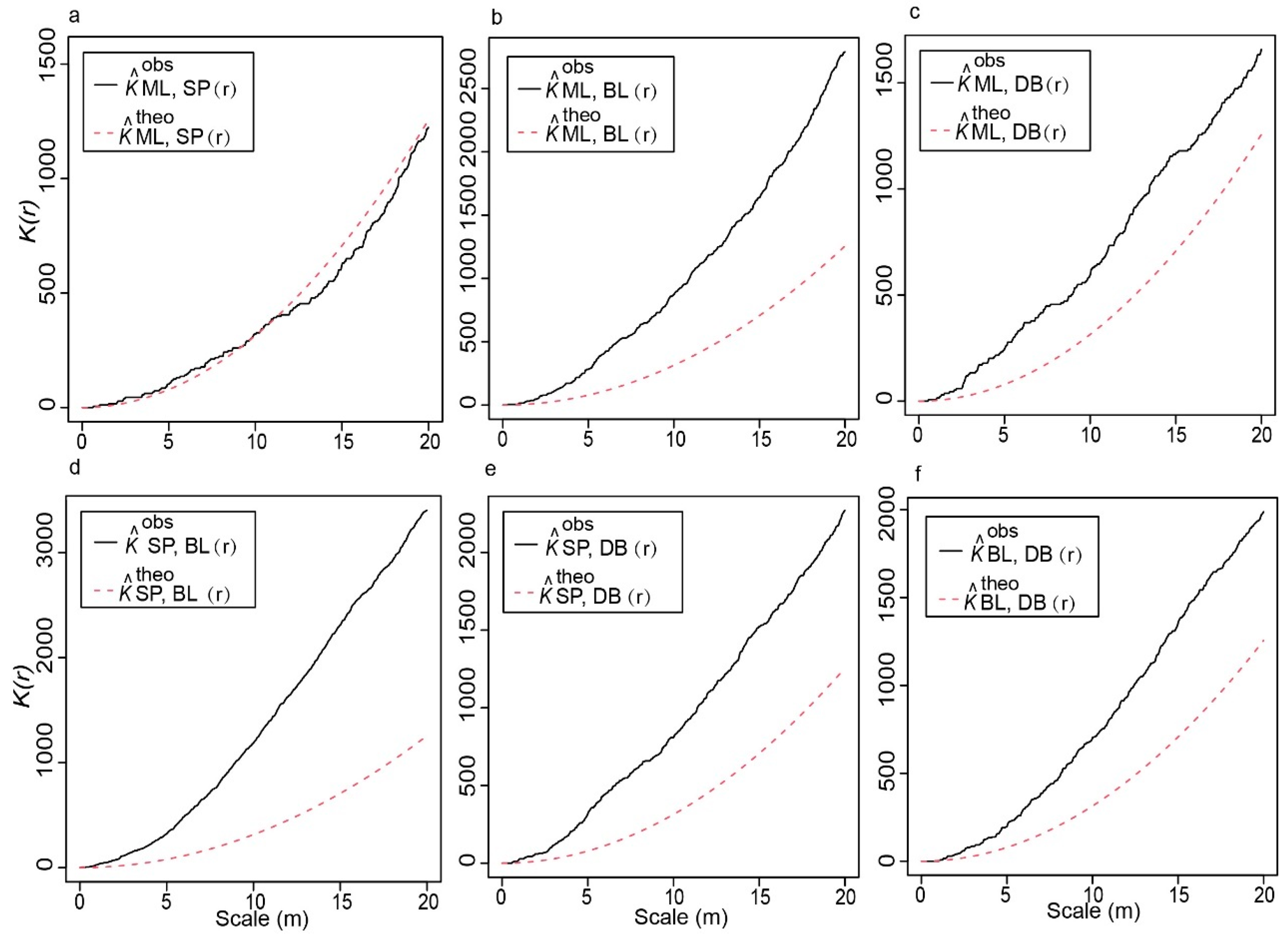
2.1. Intraspecific and Interspecific Distribution Patterns
2.2. Size Class Analysis
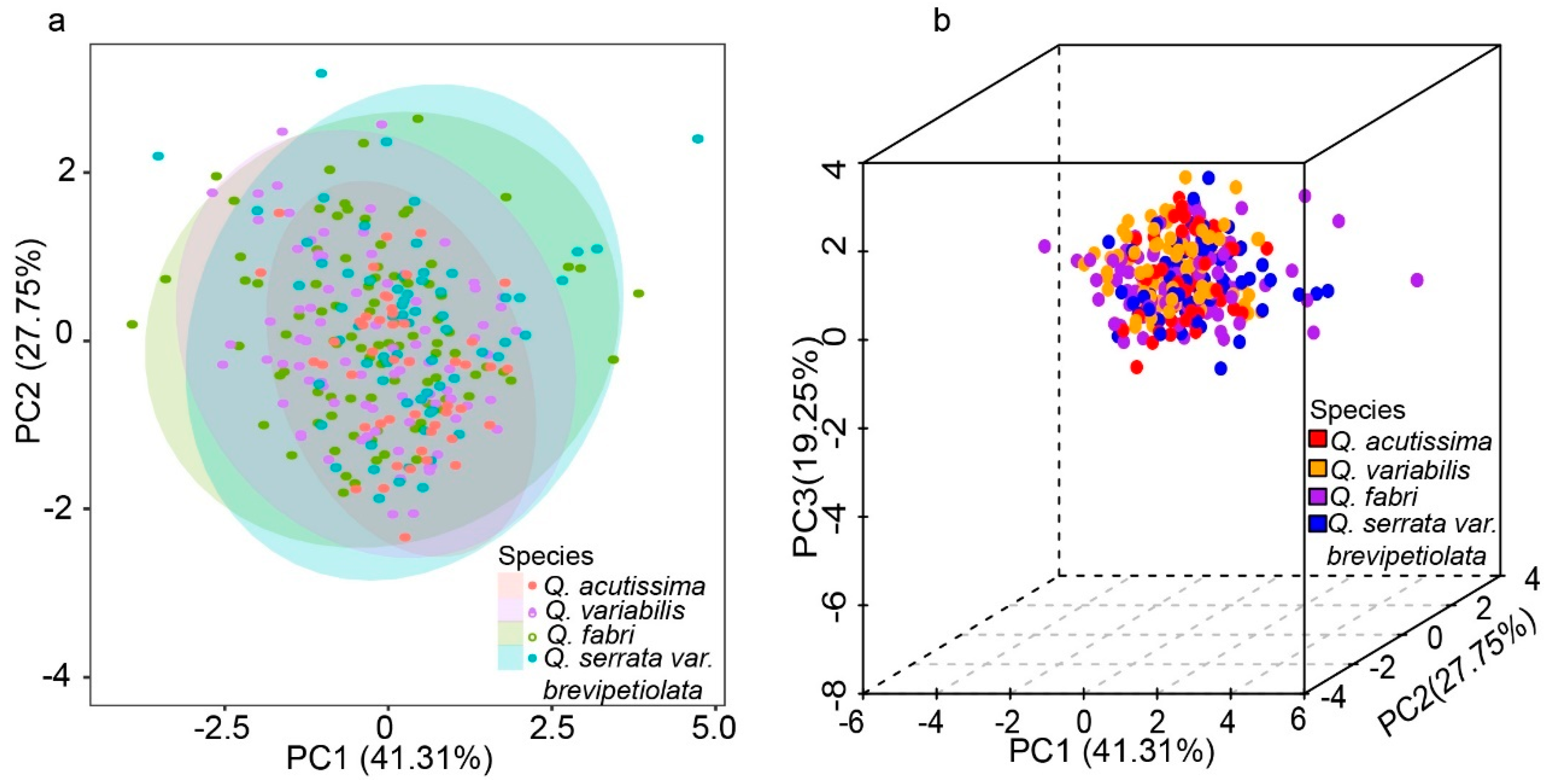
2.3. Variation Analysis of Leaf Functional Traits
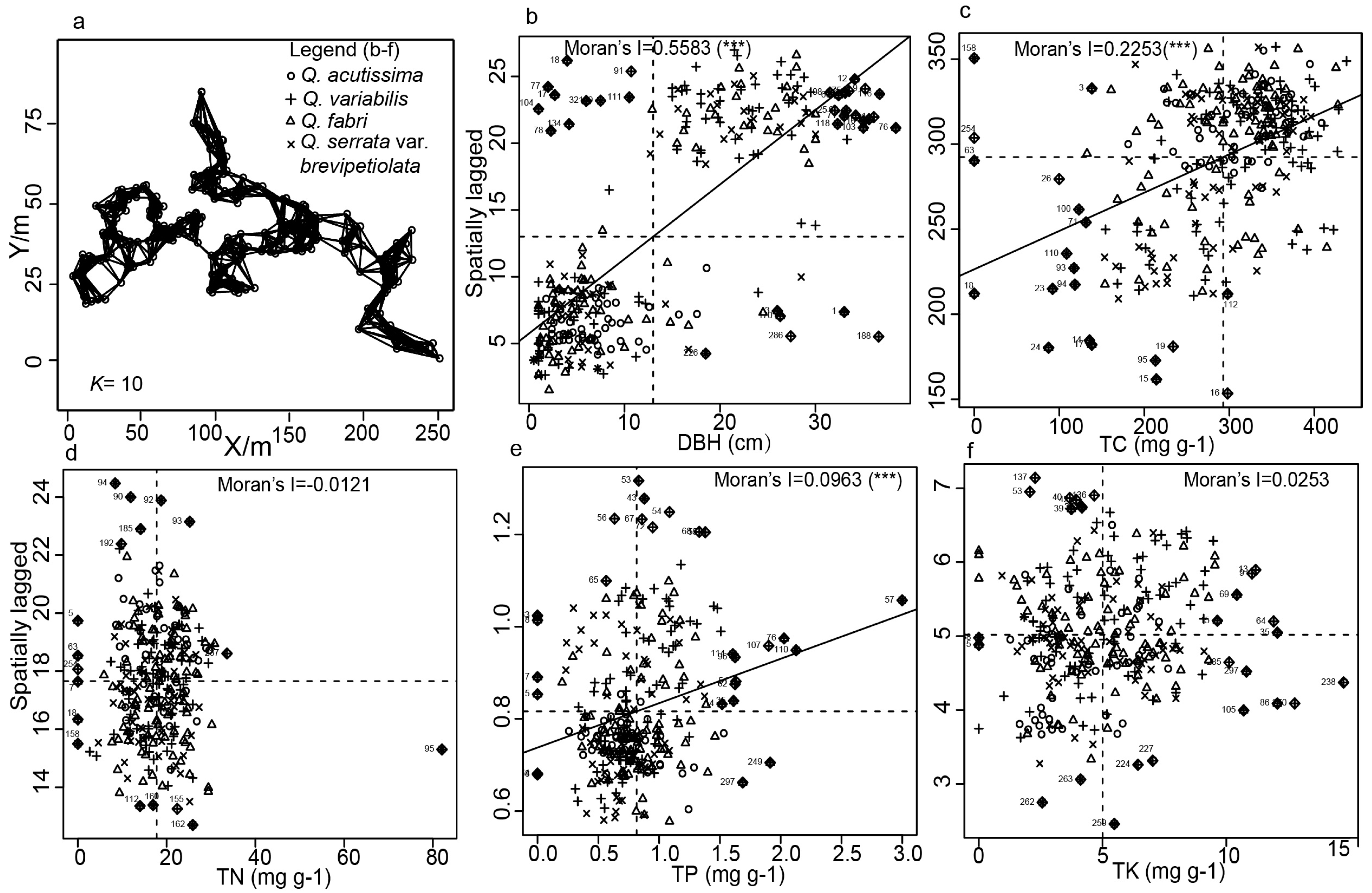
2.4. Spatial Autocorrelation Analysis of DBH and Leaf Functional Traits
3. Discussion
3.1. Spatial Distribution Patterns of Oak Mixed Forest
3.2. Influence of Leaf Functional Traits on Coexistence in Oak Species
4. Materials and Methods
4.1. Study Area
4.2. Plant Materials and Measurements
4.3. Data Analysis
4.3.1. Point Pattern Analysis
4.3.2. Breast Diameter and Nutrient Element Analysis
4.3.3. Spatial Autocorrelation Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- García-Callejas, D.; Bartomeus, I.; Godoy, O. The spatial configuration of biotic interactions shapes coexistence-area relationships in an annual plant community. Nat. Commun. 2021, 12, 6192. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, T.; Wang, X.; Anderson-Teixeira, K.J.; Bourget, N.A.; Cao, M.; Ci, X.; Davies, S.J.; Hao, Z.; Howe, R.W.; Kress, W.J.; et al. Consequences of spatial patterns for coexistence in species-rich plant communities. Nat. Ecol. Evol. 2021, 5, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Loke, L.H.; Chisholm, R.A. Measuring habitat complexity and spatial heterogeneity in ecology. Ecol. Lett. 2022, 25, 2269–2288. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, T.; Boyko, J.D.; Beaulieu, J.M. Linking mode of seed dispersal and climatic niche evolution in flowering plants. J. Biogeogr. 2023, 50, 43–56. [Google Scholar] [CrossRef]
- Snell, R.S.; Beckman, N.G.; Fricke, E.; Loiselle, B.A.; Carvalho, C.S.; Jones, L.R.; Lichti, N.I.; Lustenhouwer, N.; Schreiber, S.; Strickland, C.; et al. Consequences of intraspecific variation in seed dispersal for plant demography, communities, evolution, and global change. AoB Plants 2019, 11, plz016. [Google Scholar] [CrossRef] [PubMed]
- Broekman, M.J.E.; Muller-Landau, H.C.; Visser, M.D.; Jongejans, E.; Wright, S.; de-Kroon, H.; Adler, F. Signs of stabilisation and stable coexistence. Ecol. Lett. 2019, 22, 1957–1975. [Google Scholar] [CrossRef] [PubMed]
- Gailing, O.; Curtu, A.L. Interspecific gene flow and maintenance of species integrity in oaks. Ann. For. Res. 2014, 579, 5–18. [Google Scholar] [CrossRef]
- Cavender-Bares, J. Diversification, adaptation, and community assembly of the American oaks (Quercus), a model clade for integrating ecology and evolution. New Phytol. 2019, 221, 669–692. [Google Scholar] [CrossRef]
- Chen, J.; Zeng, Y.F.; Zhang, D.Y. Dispersal as a result of asymmetrical hybridization between two closely related oak species in China. Mol. Phylogenet. Evol. 2021, 154, 106964. [Google Scholar] [CrossRef]
- Sujii, P.S.; Tambarussi, E.V.; Grando, C.; de-Aguiar-Silvestre, E.; Viana, J.P.G.; Brancalion, P.H.S.; Zucchi, M.I. High gene flow through pollen partially compensates spatial limited gene flow by seeds for a Neotropical tree in forest conservation and restoration areas. Conserv. Genet. 2021, 22, 383–396. [Google Scholar] [CrossRef]
- Pollock, L.J.; Bayly, M.J.; Vesk, P.A.; Benjamin, M.B.; Judith, L.B. The roles of ecological and evolutionary processes in plant community assembly: The environment, hybridization, and introgression influence co-occurrence of eucalyptus. Am. Nat. 2015, 185, 784–796. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.K.; Liu, X.Y.; He, N.P.; Pan, X.; Long, S.Y.; Li, W.; Zhang, M.Y.; Cui, L.J.; Kerkhoff, A.; Kerkhoff, A. Global patterns in leaf stoichiometry across coastal wetlands. Glob. Ecol. Biogeogr. 2021, 30, 852–869. [Google Scholar] [CrossRef]
- Tong, R.; Zhou, B.; Jiang, L.; Ge, X.; Cao, Y. Spatial patterns of leaf carbon, nitrogen, and phosphorus stoichiometry and nutrient resorption in Chinese fir across subtropical China. Catena 2021, 201, 105221. [Google Scholar] [CrossRef]
- Ramírez-Valiente, J.A.; Deacon, N.J.; Etterson, J.; Center, A.; Sparks, J.P.; Sparks, K.L.; Longwell, T.; Pilz, G.; Cavender-Bares, J. Natural selection and neutral evolutionary processes contribute to genetic divergence in leaf traits across a precipitation gradient in the tropical oak Quercus oleoides. Mol. Ecol. 2018, 27, 2176–2192. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Kuramae, A.; van-Leeuwen, C.H.A.; Velthuis, M.; van-Donk, E.; Xu, J.; Bakker, E.S. Interactive effects of rising temperature and nutrient enrichment on aquatic plant growth, stoichiometry and palatability. Front. Plant Sci. 2020, 11, 58. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Shi, F.; Chen, H.; Zhang, Y.; Guo, Y.; Mao, R. Relationship between relative growth rate and C:N:P stoichiometry for the marsh herbaceous plants under water-level stress conditions. Glob. Ecol. Conserv. 2021, 25, e1416. [Google Scholar] [CrossRef]
- Wang, M.; Gong, Y.; Lafleur, P.; Wu, Y. Patterns and drivers of carbon, nitrogen and phosphorus stoichiometry in Southern China’s grasslands. Sci. Total Environ. 2021, 785, 147201. [Google Scholar] [CrossRef]
- Riva, D.L.E.G.; Villar, R.; Pérez-Ramos, I.M.; Quero, J.L.; Matías, L.; Poorter, L.; Marañón, T. Relationships between leaf mass per area and nutrient concentrations in 98 Mediterranean woody species are determined by phylogeny, habitat and leaf habit. Trees 2018, 32, 497–510. [Google Scholar] [CrossRef]
- Bai, K.; Wei, Y.; Zhang, D.; Fu, L.; Lv, S.; Deng, L. Contrasting effects of light, soil chemistry and phylogeny on leaf nutrient concentrations in cave-dwelling plants. Plant Soil 2020, 448, 105–120. [Google Scholar] [CrossRef]
- Beyns, R.; Bauman, D.; Drouet, T.; Pugnaire, F.; Pugnaire, F. Fine-scale tree spatial patterns are shaped by dispersal limitation which correlates with functional traits in a natural temperate forest. J. Veg. Sci. 2021, 32, e13070. [Google Scholar] [CrossRef]
- Gittins, R. Canonical Analysis: A Review with Applications in Ecology; Springer: Berlin/Heidelberg, Germany, 1985. [Google Scholar]
- Campbell, J.E.; Franklin, S.B.; Gibson, D.J.; Newman, J.A. Permutation of Two-Term Local Quadrat Variance Analysis: General concepts for interpretation of peaks. J. Veg. Sci. 1998, 9, 41–44. [Google Scholar] [CrossRef]
- Zhang, J. Analysis of spatial point pattern for plant species. Acta Phytoecol. Sin. 1998, 22, 344–349. [Google Scholar]
- Perea, A.J.; Wiegand, T.; Garrido, J.L.; Rey, P.J.; Alcántara, J.M. Legacy effects of seed dispersal mechanisms shape the spatial interaction network of plant species in Mediterranean forests. J. Ecol. 2021, 109, 3670–3684. [Google Scholar] [CrossRef]
- Ben-Said, M. Spatial point-pattern analysis as a powerful tool in identifying pattern-process relationships in plant ecology: An updated review. Ecol. Process. 2021, 10, 56. [Google Scholar] [CrossRef]
- Wu, J.D.; Jones, B.; Li, H.; Loucks, O.L. Scaling and Uncertainty Analysis in Ecology. Methods and Applications; Springer: Dordrecht, The Netherlands, 2006. [Google Scholar]
- Paradinas, I.; Illian, J.; Smout, S. Understanding spatial effects in species distribution models. PLoS ONE 2023, 18, e0285463. [Google Scholar] [CrossRef] [PubMed]
- Flora of China; Flora of China Editorial Committee: Beijing, China, 2018.
- Li, X.; Wei, G.; El-Kassaby, Y.A.; Fang, Y. Hybridization and introgression in sympatric and allopatric populations of four oak species. BMC Plant Biol. 2021, 21, 266. [Google Scholar] [CrossRef]
- Wei, G.; Li, X.; Fang, Y. Sympatric genome size variation and hybridization of four oak species as determined by flow cytometry genome size variation and hybridization. Ecol. Evol. 2021, 11, 1729–1740. [Google Scholar] [CrossRef]
- Wu, L.; Sun, C.; Fan, F. Estimating the characteristic spatiotemporal variation in habitat quality using the invest model—A case study from Guangdong–Hong Kong–Macao Greater Bay Area. Remote Sens. 2021, 13, 1008. [Google Scholar] [CrossRef]
- Bar-Massada, A. Complex relationships between species niches and environmental heterogeneity affect species co-occurrence patterns in modelled and real communities. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150927. [Google Scholar] [CrossRef]
- Udy, K.; Fritsch, M.; Meyer, K.M.; Grass, I.; Hanß, S.; Hartig, F.; Kneib, T.; Kreft, H.; Kukunda, C.B.; Pe’er, G.; et al. Environmental heterogeneity predicts global species richness patterns better than area. Glob. Ecol. Biogeogr. 2021, 30, 842–851. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.W.; Fang, Y.M. Predicting the impact of global warming on the geographical distribution pattern of Quercus variabilis in China Chinese. J. Appl. Ecol. 2014, 2512, 3381–3389. [Google Scholar]
- Zhang, X.; Li, Y.; Liu, C.; Xia, T.; Zhang, Q.; Fang, Y. Phylogeography of the temperate tree species Quercus acutissima in China: Inferences from chloroplast DNA variations. Biochem. Syst. Ecol. 2015, 63, 190–197. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Fang, Y. Prediction of potential suitable distribution areas of Quercus fabri in China based on an optimized Maxent model. Sci. Silvae Sin. 2018, 54, 153–164. [Google Scholar]
- Kremer, A.; Hipp, A.L. Oaks: An evolutionary success story. New Phytol. 2020, 226, 987–1011. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Wang, Z.H. Leaf morphological traits: Ecological function, geographic distribution and drivers. Chin. J. Plant Ecol. 2021, 45, 1154. [Google Scholar] [CrossRef]
- Chávez-Vergara, B.M.; González-Rodríguez, A.; Etchevers, J.D.; Oyama, K.; García-Oliva, F. Foliar nutrient resorption constrains soil nutrient transformations under two native oak species in a temperate deciduous forest in Mexico. Eur. J. For. Res. 2015, 134, 803–817. [Google Scholar] [CrossRef]
- Axer, M.; Schlicht, R.; Wagner, S. Modelling potential density of natural regeneration of European oak species (Quercus robur L., Quercus petraea (Matt.) Liebl.) depending on the distance to the potential seed source: Methodological approach for modelling dispersal from inventory data at forest enterprise level. For. Ecol. Manag. 2021, 482, 118802. [Google Scholar]
- Bai, X.; Wang, B.; An, S.; Zeng, Q.; Zhang, H. Response of forest species to C:N:P in the plant-litter-soil system and stoichiometric homeostasis of plant tissues during afforestation on the Loess Plateau, China. Catena 2021, 183, 104186. [Google Scholar] [CrossRef]
- Ripley, B.D. The second-order analysis of stationary point processes. J. Appl. Probab. 1976, 13, 255–266. [Google Scholar] [CrossRef]
- Diggle, P. Statistical Analysis of Spatial Point Patterns; Academic: London, UK, 1983. [Google Scholar]
- Baddeley, A.; Diggle, P.J.; Hardegen, A.; Lawrence, T.; Milne, R.K.; Nair, G. On tests of spatial pattern based on simulation envelopes. Ecol. Monogr. 2014, 84, 477–489. [Google Scholar] [CrossRef]
- Qu, Z.; Wen, Z.; Zhu, K. Analysis of the current state of the forest at Linggu Temple, Nanjing. J. Integr. Plant Biol. 1952, 1, 18–45. [Google Scholar]
- Moran, P. Notes on continuous stochastic phenomena. Biometrika 1950, 37, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Premo, L.S. Local spatial autocorrelation statistics quantify multi-scale patterns in distributional data: An example from the Maya Lowlands. J. Archaeol. Sci. 2004, 31, 855–866. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://wwwR-projectorg/ (accessed on 20 October 2021).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Max (cm) | Min (cm) | Average (cm) |
|---|---|---|---|
| Q. acutissima | 18.50 | 1.00 | 6.50 |
| Q. variabilis | 36.60 | 0.50 | 15.79 |
| Q. fabri | 38.40 | 1.00 | 12.37 |
| Q. serrata var. brevipetiolata | 36.70 | 0.50 | 14.89 |
| All species | 38.40 | 0.50 | 13.00 |
| Species | TC (mg g−1) | TN (mg g−1) | TP (mg g−1) | TK (mg g−1) | ||||
|---|---|---|---|---|---|---|---|---|
| Mean ± SD | Fold | Mean ± SD | Fold | Mean ± SD | Fold | Mean ± SD | Fold | |
| Q. acutissima | 313.41 ± 51.21 a | 2.26 | 17.89 ± 6.11 a | 4.00 | 0.73 ± 0.25 b | 6.08 | 4.47 ± 2.35 c | 11.89 |
| Q. variabilis | 312.66 ± 73.55 a | 4.38 | 17.69 ± 5.26 a | 11.05 | 0.87 ± 0.32 a | 6.13 | 5.34 ± 2.62 ab | 12.63 |
| Q. fabri | 290.56 ± 74.45 ab | 4.89 | 18.64 ± 5.78 a | 5.24 | 0.88 ± 0.33 a | 10.15 | 5.57 ± 2.43 a | 8.43 |
| Q. serrata var. brevipetiolata | 272.52 ± 75.22 b | 3.64 | 18.14 ± 5.69 a | 6.52 | 0.80 ± 0.44 ab | 13.04 | 4.60 ± 1.99 bc | 9.68 |
| All species | 296.85 ± 72.59 | 5.01 | 18.12 ± 6.78 | 11.58 | 0.83 ± 0.34 | 15.00 | 5.10 ± 2.42 | 15.68 |
| Moran’s I | Expectation | Variance | Standard Deviate | p-Value | |
|---|---|---|---|---|---|
| DBH | 0.5583 | −0.0033 | 0.0005 | 24.1678 | <0.000 *** |
| TC | 0.2253 | −0.0033 | 0.0005 | 9.8785 | <0.000 *** |
| TN | −0.0121 | −0.0033 | 0.0005 | −0.3952 | 0.6536 |
| TP | 0.0963 | −0.0033 | 0.0005 | 4.3324 | <0.000 *** |
| TK | 0.0253 | −0.0033 | 0.0005 | 1.2352 | 0.1084 |
| Population | Code | Section | Altitude (m) | Site | Latitude (N)/ Longitude (E) | Sample Size |
|---|---|---|---|---|---|---|
| Q. acutissima | ML | Cerris | 263 | Yushan | N31°36′/E120°40′ | 48 |
| Q. variabilis | SP | Cerris | 89 | |||
| Q. fabri | BL | Quercus | 96 | |||
| Q. serrata var. brevipetiolata | DB | Quercus | 67 | |||
| Total | 300 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Li, Y.; El-Kassaby, Y.A.; Fang, Y. Spatial Distribution and Ecological Determinants of Coexisting Hybrid Oak Species: A Study in Yushan’s Mixed Forest. Plants 2024, 13, 1000. https://doi.org/10.3390/plants13071000
Li X, Li Y, El-Kassaby YA, Fang Y. Spatial Distribution and Ecological Determinants of Coexisting Hybrid Oak Species: A Study in Yushan’s Mixed Forest. Plants. 2024; 13(7):1000. https://doi.org/10.3390/plants13071000
Chicago/Turabian StyleLi, Xuan, Yongfu Li, Yousry A. El-Kassaby, and Yanming Fang. 2024. "Spatial Distribution and Ecological Determinants of Coexisting Hybrid Oak Species: A Study in Yushan’s Mixed Forest" Plants 13, no. 7: 1000. https://doi.org/10.3390/plants13071000
APA StyleLi, X., Li, Y., El-Kassaby, Y. A., & Fang, Y. (2024). Spatial Distribution and Ecological Determinants of Coexisting Hybrid Oak Species: A Study in Yushan’s Mixed Forest. Plants, 13(7), 1000. https://doi.org/10.3390/plants13071000