

Interaction of Soybean (Glycine max (L.) Merr.) Class II ACBPs with MPK2 and SAPK2 Kinases: New Insights into the Regulatory Mechanisms of Plant ACBPs

Abstract
:1. Introduction
2. Results
2.1. Interaction between Class II GmACBPs and Soybean Kinases
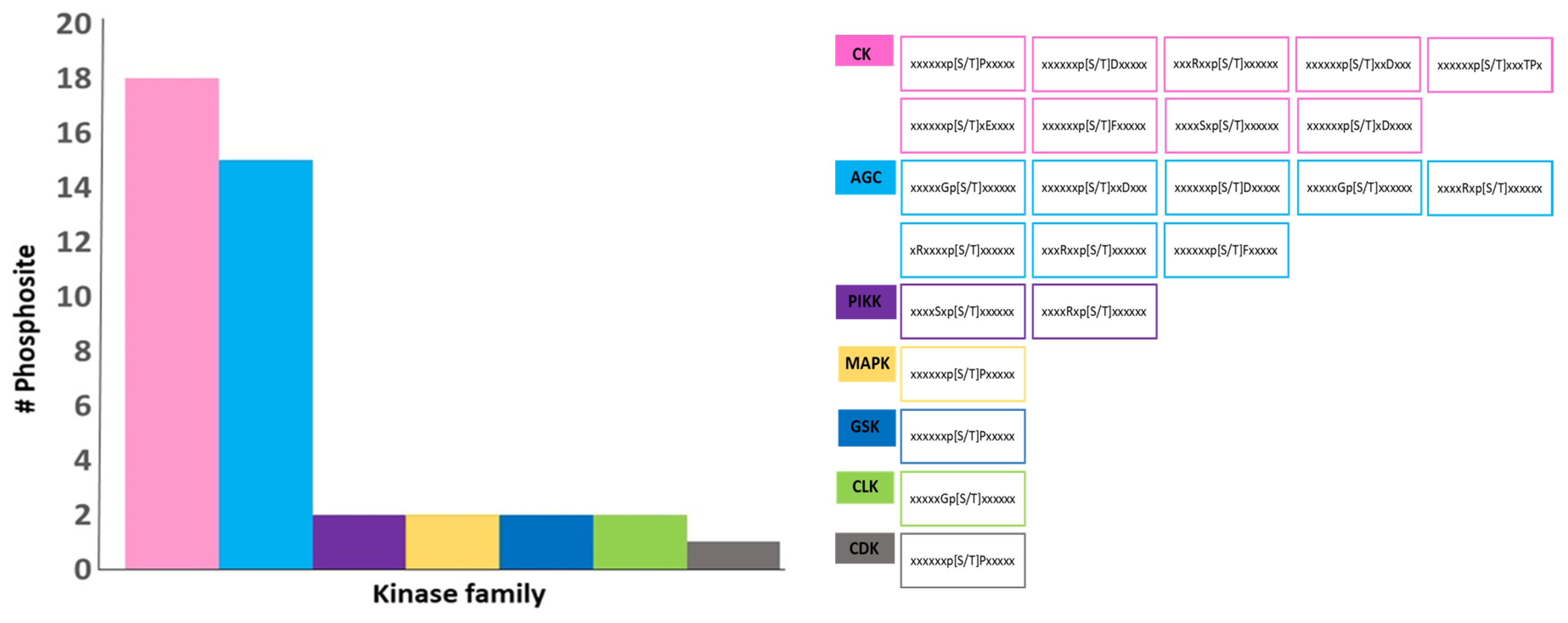
2.2. Prediction of Kinase-Specific Phosphorylation Sites in Class II GmACBPs and Docking-Based Binding Models of Class II GmACBPs and Two Kinases
3. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name in Arabidopsis thaliana | Accession Number (NCBI) | Glycine max Homolog a | Accession Number (NCBI) | Refs. |
|---|---|---|---|---|
| Mitogen-activated protein kinase 3 | NP_190150.1 | Mitogen-activated protein kinase 13 | XP_003538034.1 | [35,55] |
| Mitogen-activated protein kinase 6 | NP_181907.1 | Mitogen-activated protein kinase 2 | NP_001235426.1 | [34,35,55] |
| Snf1-related protein kinases SnRK2.8 | NP_001077839.1.1 | Serine/threonine-protein kinase SAPK2-1 | XP_003519175.1.1 | [39,42] |
| Snf1-related protein kinases SnRK2.7 | NP_195711.1 | Serine/threonine-protein kinase SAPK2-2 | XP_003531338.1 | [39,42] |
| Snf1-related protein kinases SnRK2.2 | NP_190619.1 | Serine/threonine-protein kinase SRK2I | XP_003550077.3 | [56] |
| Casein kinase II, Beta chain 1 | NP_001190483.1 | Putative casein kinase II subunit beta | NP_001344382.1 | [53,57,58] |
| Calcium-dependent protein kinase 4) | NP_192695.1 | Calcium-dependent protein kinase 4 | XP_006589459.2 | [59] |
4. Materials and Methods
4.1. Plasmids Construction for Yeast Two-Hybrid Assay
4.2. Yeast Two-Hybrid Assay
4.3. Generation of Bimolecular Fluorescence Complementation Screen (BiFC) Constructs
4.4. Confocal Laser Scanning Microscopy
4.5. Western Blot Assay
4.6. Recombinant Protein Expression and Purification for In Vitro Kinase Assay
4.7. In Vitro Kinase Assay and Phos-Tag SDS-PAGE
4.8. Prediction of Kinase-Specific Phosphorylation Sites in Class II GmACBPs and Docking-Based Binding Models of Class II GmACBPs and Two Kinases
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACB | acyl-CoA-binding domain |
| ACBPs | acyl-CoA-binding proteins |
| AGC | cAMP-dependent protein kinase |
| ANK | ankyrin-repeat |
| AREB1 | ABA-RESPONSIVE ELEMENT BINDING TRANSCRIPTION FACTOR1 |
| BiFC | bimolecular fluorescence complementation |
| CDK | cyclin-dependent kinase |
| CDPK | calcium-dependent protein kinase |
| CK | casein kinase |
| CLK | Cdc2-like kinase |
| DBI | diazepam binding inhibitor |
| DDO | synthetic dropout medium/-Leu/-Trp |
| ER | endoplasmic reticulum |
| GABA | gamma-aminobutyric acid |
| GmACBPs | Glycine max ACBPs |
| GmLOX1 | Glycine max LIPOXYGENASE1 |
| GSK | glycogen synthase kinase |
| MPK | mitogen-activated protein kinase |
| PIKK | phosphatidylinositol-3 kinase-related kinase |
| SAPK | serine/threonine-protein kinase |
| SnRK | SNF-related serine/threonine-protein kinase |
| TDO/X/A | synthetic dropout medium/−His/−Leu/−Trp containing 40 µg mL−1 5-bromo-4-chloro-3-indolyl-α-D-galactopyranoside (X-α-Gal) and 125 ng mL−1 aureobasidin A |
| Y2H | yeast-two hybrid |
References
- Mikkelsen, J.; Knudsen, J. Acyl-CoA-binding protein from cow. Binding characteristics and cellular and tissue distribution. Biochem. J. 1987, 15, 709–714. [Google Scholar] [CrossRef] [PubMed]
- Mandrup, S.; Hummel, R.; Ravn, S.; Jensen, G.; Andreasen, P.H.; Gregersen, N.; Knudsen, J.; Kristiansen, K. Acyl-CoA-binding protein/diazepam-binding inhibitor gene and pseudogenes. A typical housekeeping gene family. J. Mol. Biol. 1992, 5, 1011–1022. [Google Scholar] [CrossRef]
- Elle, K.T.; Simonsen, L.C.; Olsen, P.K.; Birck, S.; Ehmsen, S.; Tuck, N.J.; Færgeman, N.J. Tissue- and paralogue-specific functions of acyl-CoA-binding proteins in lipid metabolism in Caenorhabditis elegans. Biochem. J. 2011, 437, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Færgeman, N.J.; Knudsen, J. Acyl-CoA binding protein is an essential protein in mammalian cell lines. Biochem. J. 2002, 15, 679–682. [Google Scholar] [CrossRef]
- Lung, S.C.; Chye, M.L. Deciphering the roles of acyl-CoA-binding proteins in plant cells. Protoplasma 2016, 253, 1177–1195. [Google Scholar] [CrossRef]
- Hsiao, A.S.; Haslam, R.P.; Michaelson, L.V.; Liao, P.; Chen, Q.F.; Sooriyaarachchi, S.; Chye, M.L. Arabidopsis cytosolic acyl-CoA-binding proteins ACBP4, ACBP5 and ACBP6 have overlapping but distinct roles in seed development. Biosci. Rep. 2014, 35, e00165. [Google Scholar] [CrossRef]
- Hsiao, A.S.; Yeung, E.C.; Ye, Z.W.; Chye, M.L. The Arabidopsis cytosolic Acyl-CoA-binding proteins play combinatory roles in pollen development. Plant Cell Physiol. 2014, 56, 322–333. [Google Scholar] [CrossRef]
- Chen, Q.F.; Xiao, S.; Chye, M.L. Overexpression of the Arabidopsis 10-kilodalton acyl-coenzyme A-binding protein ACBP6 enhances freezing tolerance. Plant Physiol. 2008, 148, 304–315. [Google Scholar] [CrossRef]
- Liao, P.; Chen, Q.F.; Chye, M.L. Transgenic Arabidopsis flowers overexpressing acyl-CoA-binding protein ACBP6 are freezing tolerant. Plant Cell Physiol. 2014, 55, 1055–1071. [Google Scholar] [CrossRef]
- Du, Z.Y.; Xiao, S.; Chen, Q.F.; Chye, M.L. Arabidopsis acyl-CoA-binding proteins ACBP1 and ACBP2 show different roles in freezing stress. Plant Signal. Behavior. 2010, 5, 607–609. [Google Scholar] [CrossRef]
- Xiao, M.L.; Chye, M.L. Overexpression of Arabidopsis ACBP3 enhances NPR1-dependent plant resistance to Pseudomonas syringe pv tomato DC3000. Plant Physiol. 2011, 156, 2069–2081. [Google Scholar] [CrossRef]
- Du, Z.Y.; Chen, M.X.; Chen, Q.F.; Xiao, S.; Chye, M.L. Overexpression of Arabidopsis acyl-CoA-binding protein ACBP2 enhances drought tolerance. Plant Cell Environ. 2013, 36, 300–314. [Google Scholar] [CrossRef]
- Meng, W.; Su, Y.C.; Saunders, R.M.; Chye, M.L. The rice acyl-CoA-binding protein gene family: Phylogeny, expression and functional analysis. New Phytol. 2011, 189, 1170–1184. [Google Scholar] [CrossRef]
- Lung, S.C.; Lai, S.H.; Wang, H.; Zhang, X.; Liu, A.; Guo, Z.H.; Chye, M.L. Oxylipin signaling in salt-stressed soybean is modulated by ligand-dependent interaction of Class II acyl-CoA-binding proteins with lipoxygenase. Plant Cell 2022, 34, 1117–1143. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Xiao, S.; Li, H.Y.; Tsao, S.W.; Chye, M.L. Arabidopsis thaliana acyl-CoA-binding protein ACBP2 interacts with heavy-metal-binding farnesylated protein AtFP6. New Phytol. 2009, 181, 89–102. [Google Scholar] [CrossRef]
- Du, Z.Y.; Chen, M.X.; Chen, Q.F.; Gu, J.D.; Chye, M.L. Expression of Arabidopsis acyl-CoA-binding proteins AtACBP1 and AtACBP4 confers Pb(II) accumulation in Brassica juncea roots. Plant Cell Environ. 2015, 38, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.J.; Yu, L.J.; Chen, Q.F.; Wang, F.Z.; Huang, L.; Xia, F.N.; Xiao, S. Arabidopsis acyl-CoA-binding protein ACBP3 participates in plant response to hypoxia by modulating very-long-chain fatty acid metabolism. Plant J. 2015, 81, 53–67. [Google Scholar] [CrossRef]
- Leung, K.C.; Li, H.Y.; Mishra, G.; Chye, M.L. ACBP4 and ACBP5, novel Arabidopsis acyl-CoA-binding proteins with kelch motifs that bind oleoyl-CoA. Plant Mol. Biol. 2004, 55, 297–309. [Google Scholar] [CrossRef]
- Liao, P.; Woodfield, H.K.; Harwood, J.L.; Chye, M.L.; Scofield, S. Comparative transcriptomics analysis of Brassica napus L. during seed maturation reveals dynamic changes in gene expression between embryos and seed coats and distinct expression profiles of acyl-CoA-binding proteins for lipid accumulation. Plant Cell Physiol. 2019, 60, 2812–2825. [Google Scholar] [CrossRef]
- Amiruddin, N.; Chan, P.L.; Azizi, N.; Morris, P.E.; Chan, K.L.; Ong, P.W.; Rosli, R.; Masura, S.S.; Murphy, D.J.; Sambanthamurthi, R.; et al. Characterisation of oil palm acyl-CoA-binding proteins and correlation of their gene expression with oil synthesis. Plant Cell Physiol. 2020, 61, 735–745. [Google Scholar] [CrossRef]
- Azlan, N.S.; Guo, Z.H.; Yung, W.S.; Wang, Z.; Lam, H.M.; Lung, S.C.; Chye, M.L. In silico analysis of acyl-CoA-binding protein expression in soybean. Front. Plant Sci. 2021, 12, 646938. [Google Scholar] [CrossRef] [PubMed]
- Moradi, A.; Dai, S.; Wong, E.O.Y.; Zhu, G.; Yu, F.; Lam, H.M.; Li, N. Isotopically dimethyl labeling-based quantitative proteomic analysis of phosphoproteomes of soybean cultivars. Biomolecules 2021, 11, 1218. [Google Scholar] [CrossRef] [PubMed]
- Maklad, H.R.; Gutierrez, G.L.; Esser, D.; Siebers, B.; Peeters, E. Phosphorylation of the acyl-CoA binding pocket of the FadR transcription regulator in Sulfolobus acidocaldarius. Biochimie 2020, 175, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Esmaili, F.; Pourmirzaei, M.; Ramazi, S.; Shojaeilangari, S.; Yavari, E. A review of machine learning and algorithmic methods for protein phosphorylation sites prediction. arXiv 2022, arXiv:2208.04311. [Google Scholar] [CrossRef] [PubMed]
- Groban, E.S.; Narayanan, A.; Jacobson, M.P. Conformational changes in protein loops and helices induced by post-translational phosphorylation. PLoS Comput. Biol. 2006, 2, e32. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Yaffe, M.B. Protein regulation in signal transduction. Cold Spring Harb. Perspect. Biol. 2016, 8, a005918. [Google Scholar] [CrossRef] [PubMed]
- Neess, D.; Bek, S.; Engelsby, H.; Gallego, S.F.; Færgeman, N.J. Long-chain acyl-CoA esters in metabolism and signaling: Role of acyl-CoA binding proteins. Prog. Lipid Res. 2015, 59, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Guidotti, A.; Forchetti, C.M.; Corda, M.G.; Konkel, D.; Bennett, C.D. Isolation, characterization, and purification to homogeneity of an endogenous polypeptide with agonistic action on benzodiazepine receptors. Proc. Natl. Acad. Sci. USA 1983, 80, 3531–3535. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, J.; Neergaard, T.B.; Gaigg, B.; Jensen, M.V.; Hansen, J.K. Role of acyl-CoA binding protein in acyl-CoA metabolism and acyl-CoA-mediated cell signaling. J. Nutr. 2000, 130, 294S–298S. [Google Scholar] [CrossRef]
- Fox, S.R.; Rawsthorne, S.; Hills, M.J. Role of acyl-CoAs and acyl-CoA-binding protein in the regulation of carbon supply for fatty acid biosynthesis. Biochem. Soc. Trans. 2000, 28, 672–674. [Google Scholar] [CrossRef]
- Udupa, P.; Kumar, A.; Parit, R.; Ghosh, D.K. Acyl-CoA binding protein regulates nutrient-dependent autophagy. Metabolism 2023, 145, 155338. [Google Scholar] [CrossRef] [PubMed]
- Bigeard, J.; Hirt, H. Nuclear signaling of plant MAPKs. Front. Plant Sci. 2018, 9, 469. [Google Scholar] [CrossRef] [PubMed]
- Droillard, M.J.; Boudsocq, M.; Barbier-Brygoo, H.; Laurière, C. Different protein kinase families are activated by osmotic stresses in Arabidopsis thaliana cell suspensions: Involvement of the MAP kinases AtMPK3 and AtMPK6. FEBS Lett. 2002, 527, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Nie, J.; Cao, C.; Jin, Y.; Yan, M.; Wang, F.; Zhang, W. Phosphatidic acid mediates salt stress response by regulation of MPK6 in Arabidopsis thaliana. New Phytol. 2010, 188, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, D.M.; Yu, W.W.; Shi, L.L.; Zhang, Y.; Lai, Y.X.; Xiao, S. Phosphatidic acid modulates MPK3- and MPK6-mediated hypoxia signaling in Arabidopsis. Plant Cell 2022, 34, 889–909. [Google Scholar] [CrossRef] [PubMed]
- Licausi, F.; Kosmacz, M.; Weits, D.A.; Giuntoli, B.; Giorgi, F.M.; Voesenek, L.M.; Van Dongen, J.T. Oxygen sensing in plants is mediated by an N-end rule pathway for protein destabilization. Nature 2011, 479, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.Y.; Chen, M.X.; Chen, Q.F.; Xiao, S.; Chye, M.L. Arabidopsis acyl-CoA-binding protein ACBP1 participates in the regulation of seed germination and seedling development. Plant J. 2013, 74, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Halford, N.G.; Hardie, D.G. SNF1-related protein kinases: Global regulators of carbon metabolism in plants? Plant Mol. Biol. 1998, 37, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Mizoguchi, M.; Umezawa, T.; Nakashima, K.; Kidokoro, S.; Takasaki, H.; Fujita, Y.; Shinozaki, K. Two closely related sub-class II SnRK2 protein kinases cooperatively regulate drought-inducible gene expression. Plant Cell Physiol. 2010, 51, 842–847. [Google Scholar] [CrossRef]
- Shin, R.; Alvarez, S.; Burch, A.Y.; Jez, J.M.; Schachtman, D.P. Phosphoproteomic identification of targets of the Arabidopsis sucrose nonfermenting-like kinase SnRK2.8 reveals a connection to metabolic processes. Proc. Natl. Acad. Sci. USA 2007, 104, 6460–6465. [Google Scholar] [CrossRef]
- Umezawa, T.; Yoshida, R.; Maruyama, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. SRK2C, a SNF1-related protein kinase 2, improves drought tolerance by controlling stress-responsive gene expression in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2004, 101, 17306–17311. [Google Scholar] [CrossRef] [PubMed]
- Boudsocq, M.; Barbier-Brygoo, H.; Lauriere, C. Identification of nine sucrose nonfermenting 1-related protein kinases 2 activated by hyperosmotic and saline stresses in Arabidopsis thaliana. J. Biol. Chem. 2004, 279, 41758–41766. [Google Scholar] [CrossRef]
- Fujita, K.; Nakashima, T.; Yoshida, T.; Katagiri, S.; Kidokoro, N.; Kanamori, K.; Yamaguchi-Shinozaki, K. Three SnRK2 protein kinases are the main positive regulators of abscisic acid signaling in response to water stress in Arabidopsis. Plant Cell Physiol. 2009, 50, 2123–2132. [Google Scholar] [CrossRef]
- Tse, M.H. Investigations on Recombinant Arabidopsis acyl-Coenzyme a Binding Protein 1. Master’s Thesis, The University of Hong Kong (HKU), Hong Kong, 2005. [Google Scholar]
- Furihata, T.; Maruyama, K.; Fujita, Y.; Umezawa, T.; Yoshida, R.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Abscisic acid-dependent multisite phosphorylation regulates the activity of a transcription activator AREB1. Proc. Natl. Acad. Sci. USA 2005, 103, 1988–1993. [Google Scholar] [CrossRef] [PubMed]
- Lung, S.C.; Liao, P.; Yeung, E.C.; Hsiao, A.S.; Xue, Y.; Chye, M.L. Acyl-CoA-binding protein ACBP1 modulates sterol synthesis during embryogenesis. Plant Physiol. 2017, 174, 1420–1435. [Google Scholar] [CrossRef]
- Lung, S.C.; Liao, P.; Yeung, E.C.; Hsiao, A.S.; Xue, Y.; Chye, M.L. Arabidopsis ACYL-COA-BINDING PROTEIN 1 interacts with STEROL C4-METHYL OXIDASE 1-2 to modulate gene expression of homeodomain-leucine zipper IV transcription factors. New Phytol. 2018, 218, 183–200. [Google Scholar] [CrossRef]
- Li, J.; Mahajan, A.; Tsai, M.D. Ankyrin repeat: A unique motif mediating protein-protein interactions. Biochemistry 2006, 45, 15168–15178. [Google Scholar] [CrossRef]
- Gao, W.; Li, H.Y.; Xiao, S.; Chye, M.L. Acyl-CoA-binding protein 2 binds lysophospholipase 2 and lysoPC to promote tolerance to cadmium-induced oxidative stress in transgenic Arabidopsis. Plant J. 2010, 62, 989–1003. [Google Scholar] [CrossRef]
- Li, H.Y.; Chye, M.L. Arabidopsis acyl-CoA-binding protein ACBP2 interacts with an ethylene-responsive element-binding protein, AtEBP, via its ankyrin repeats. Plant Mol. Biol. 2004, 54, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Rentel, M.C.; Lecourieux, D.; Ouaked, F.; Usher, S.L.; Petersen, L.; Okamoto, H.; Knight, H.; Peck, S.C.; Grierson, C.S.; Hirt, H.; et al. OXI1 kinase is necessary for oxidative burst-mediated signalling in Arabidopsis. Nature 2004, 26, 858–861. [Google Scholar] [CrossRef] [PubMed]
- Hirt, H.; Garcia, A.V.; Oelmüller, R. AGC kinases in plant development and defense. Plant Signal Behav. 2011, 6, 1030–1033. [Google Scholar] [CrossRef] [PubMed]
- Nagatoshi, Y.; Fujita, M.; Fujita, Y. Casein kinase 2 α and β subunits inversely modulate ABA signal output in Arabidopsis protoplasts. Planta 2018, 248, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Wei, Z.; Chen, H.H.; Liu, T.; Liao, K.; Xue, H.W. Plant casein kinases phosphorylate and destabilize a cyclin-dependent kinase inhibitor to promote cell division. Plant Physiol. 2021, 187, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Ahlfors, R.; Macioszek, V.; Rudd, J.; Brosché, M.; Schlichting, R.; Scheel, D.; Kangasjärvi, J. Stress hormone-independent activation and nuclear translocation of mitogen-activated protein kinases in Arabidopsis thaliana during ozone exposure. Plant J. 2004, 40, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Xue, L.; Batelli, G.; Lee, S.; Hou, Y.J.; Van Oosten, M.J.; Zhu, J.K. Quantitative phosphoproteomics identifies SnRK2 protein kinase substrates and reveals the effectors of abscisic acid action. Proc. Natl. Acad. Sci. USA 2013, 110, 11205–11210. [Google Scholar] [CrossRef] [PubMed]
- Salinas, P.; Fuentes, D.; Vidal, E.; Jordana, X.; Echeverria, M.; Holuigue, L. An extensive survey of CK2 α and β subunits in Arabidopsis: Multiple isoforms exhibit differential subcellular localization. Plant Cell Physiol. 2006, 47, 1295–1308. [Google Scholar] [CrossRef] [PubMed]
- Riera, M.; Figueras, M.; López, C.; Goday, A.; Pagès, M. Protein kinase CK2 modulates developmental functions of the abscisic acid responsive protein Rab17 from maize. Proc. Natl. Acad. Sci. USA 2004, 101, 9879–9884. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.Y.; Yu, X.C.; Wang, X.J.; Zhao, R.; Li, Y.; Fan, R.C.; Zhang, D.P. Two calcium-dependent protein kinases, CPK4 and CPK11, regulate abscisic acid signal transduction in Arabidopsis. Plant Cell 2007, 19, 3019–3036. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Fukuda, Y.; Murata, K.; Kimura, A. Transformation of intact yeast cells treated with alkali cations. J. Bacteriol. 1983, 153, 163–168. [Google Scholar] [CrossRef]
- Walter, M.; Chaban, C.; Schütze, K.; Batistic, O.; Weckermann, K.; Nake, C.; Blazevic, D.; Grefen, C.; Schumacher, K.; Oecking, C. Visualization of protein interactions in living plant cells using biomolecular fluorescence complementation. Plant J. 2004, 40, 428–438. [Google Scholar] [CrossRef]
- Jyothishwaran, G.; Kotresha, D.; Selvaraj, T.; Srideshikan, S.M.; Rajvanshi, P.K.; Jayabaskaran, C. A modified freeze-thaw method for efficient transformation of Agrobacterium tumefaciens. Curr. Sci. 2007, 93, 770–772. [Google Scholar]
- Zhang, T.; Lei, J.; Yang, H.; Xu, K.; Wang, R.; Zhang, Z. An improved method for whole protein extraction from yeast Saccharomyces cerevisiae. Yeast 2011, 28, 795–798. [Google Scholar] [CrossRef] [PubMed]
- Miao, R.; Lung, S.C.; Li, X.; Li, X.D.; Chye, M.L. Thermodynamic insights into an interaction between ACYL-CoA-BINDING PROTEIN2 and LYSOPHOSPHOLIPASE2 in Arabidopsis. J. Biol. Chem. 2019, 294, 6214–6226. [Google Scholar] [CrossRef]
- Guo, Z.H.; Chan, W.H.Y.; Kong, G.K.W.; Hao, Q.; Chye, M.L. The first plant acyl-CoA-binding protein structures: The close homologues OsACBP1 and OsACBP2 from rice. Acta Crystallogr. D 2017, 73, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Peck, S.C. Analysis of protein phosphorylation: Methods and strategies for studying kinases and substrates. Plant J. 2006, 45, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Wang, C.; Zhou, J.; Shi, Y.; Ruan, C.; Tu, Y.; Xue, Y. EPSD: A well-annotated data resource of protein phosphorylation sites in eukaryotes. Brief. Bioinform. 2011, 22, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zhang, W.; Gou, Y.; Xu, D.; Wei, Y.; Liu, D.; Xue, Y. GPS 6.0: An updated server for predicting kinase-specific phosphorylation sites in proteins. Nucleic Acids Res. 2023, 51, W243–W250. [Google Scholar] [CrossRef] [PubMed]
- Blom, N.; Sicheritz-Pontén, T.; Gupta, R.; Gammeltoft, S.; Brunak, S. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence. Proteomics 2004, 4, 1633–1649. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zhang, D.; Zhou, P.; Li, B.; Huang, S.Y. HDOCK: A web server for protein-protein and protein-DNA/RNA docking based on a hybrid strategy. Nucleic Acids Res. 2017, 45, W365–W373. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction, and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moradi, A.; Lung, S.-C.; Chye, M.-L. Interaction of Soybean (Glycine max (L.) Merr.) Class II ACBPs with MPK2 and SAPK2 Kinases: New Insights into the Regulatory Mechanisms of Plant ACBPs. Plants 2024, 13, 1146. https://doi.org/10.3390/plants13081146
Moradi A, Lung S-C, Chye M-L. Interaction of Soybean (Glycine max (L.) Merr.) Class II ACBPs with MPK2 and SAPK2 Kinases: New Insights into the Regulatory Mechanisms of Plant ACBPs. Plants. 2024; 13(8):1146. https://doi.org/10.3390/plants13081146
Chicago/Turabian StyleMoradi, Atieh, Shiu-Cheung Lung, and Mee-Len Chye. 2024. "Interaction of Soybean (Glycine max (L.) Merr.) Class II ACBPs with MPK2 and SAPK2 Kinases: New Insights into the Regulatory Mechanisms of Plant ACBPs" Plants 13, no. 8: 1146. https://doi.org/10.3390/plants13081146