
Oxidative Stress Response Mechanisms Sustain the Antibacterial and Antioxidant Activity of Quercus ilex

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Chemical Characterization of Q. ilex Leaves Extracts
2.1.1. Hydroxicinammic Acids
2.1.2. Total Flavonoid Content
2.1.3. Total Phenolic Content
2.1.4. Reduced Glutathione
2.1.5. Ascorbic Acid
2.2. In Vitro Antioxidant Activities of Q. ilex Leaves Extracts
2.2.1. Ferric Reducing Antioxidant Power (FRAP) Assay
2.2.2. 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Assay
2.2.3. 2,2′-Azino-bis-3-ethylbenzothiazoline-6-sulfonic Acid (ABTS) Assay
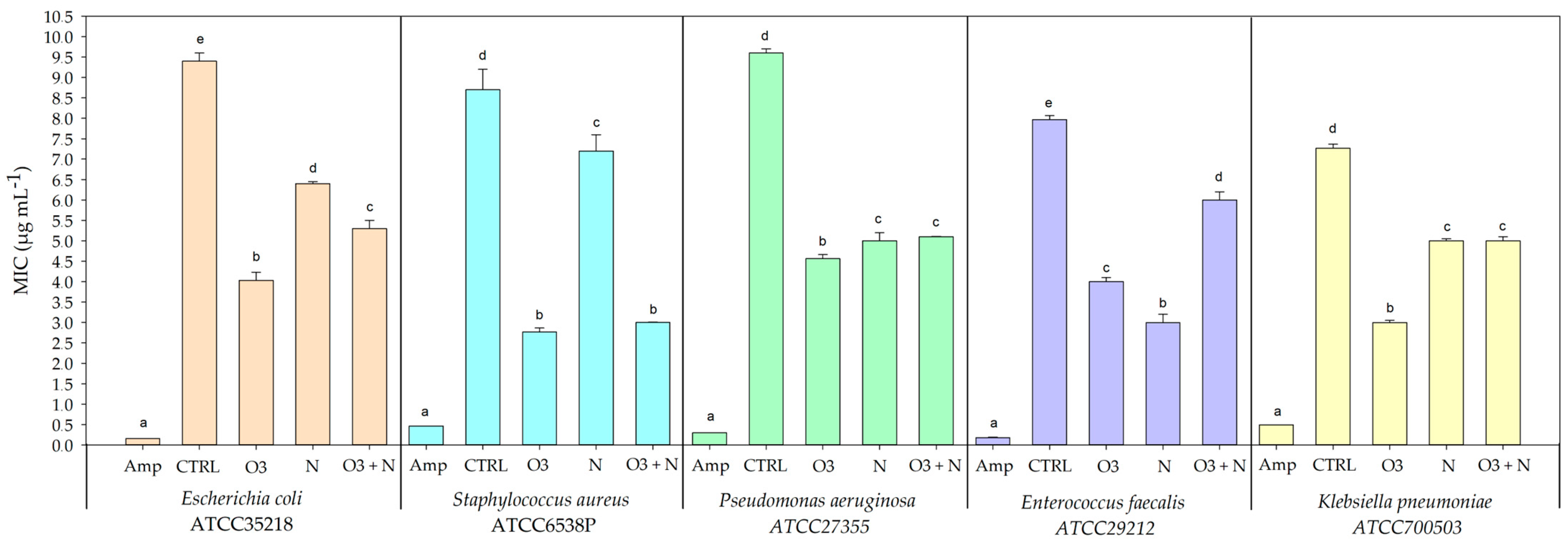
2.3. Antibacterial Activity of Q. ilex Leaves Extracts
3. Discussion
4. Materials and Methods
4.1. Growth Conditions
4.2. Experimental Design
4.3. Plants Extraction
4.4. Determination of Total Phenolic Compounds, Hydroxycinnamic Acids, and Flavonoid Content
Determination of Ascorbic Acid and Reduced Glutation (GSH) Content
4.5. Antioxidant Activity
4.6. Antimicrobial Activity
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lushchak, V.I. Free Radicals, Reactive Oxygen Species, Oxidative Stress and Its Classification. Chem. Biol. Interact. 2014, 224, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive Oxygen Species, Abiotic Stress and Stress Combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Chan, Z. ROS Regulation During Abiotic Stress Responses in Crop Plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [PubMed]
- Castagna, A.; Ranieri, A. Detoxification and Repair Process of Ozone Injury: From O3 Uptake to Gene Expression Adjustment. Environ. Pollut. 2009, 157, 1461–1469. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Miller, G.; Sejima, H.; Harper, J.; Mittler, R. Enhanced Seed Production under Prolonged Heat Stress Conditions in Arabidopsis Thaliana Plants Deficient in Cytosolic Ascorbate Peroxidase 2. J. Exp. Bot. 2013, 64, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Chakraborty, S.; Mukherjee, S.; Sau, A.; Das, S.; Chakraborty, B.; Mitra, S.; Adak, S.; Goswami, A.; Hessel, V. A Comparative Study on the Modulatory Role of Mesoporous Silica Nanoparticles MCM 41 and MCM 48 on Growth and Metabolism of Dicot Vigna Radiata. Plant Physiol. Biochem. 2022, 187, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Shang, B.; Li, Z.; Calatayud, V.; Agathokleous, E. Ozone Will Remain a Threat for Plants Independently of Nitrogen Load. Funct. Ecol. 2019, 33, 1854–1870. [Google Scholar] [CrossRef]
- Moura, B.B.; Paoletti, E.; Badea, O.; Ferrini, F.; Hoshika, Y. Visible Foliar Injury and Ecophysiological Responses to Ozone and Drought in Oak Seedlings. Plants 2022, 11, 1836. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, L.; Salvatori, E.; Manes, F. Effects of Nitrogen Deposition, Drought and Their Interaction, on Functional and Structural Traits of Fraxinus ornus L. and Q. ilex L. Plant Biosyst. 2017, 151, 174–189. [Google Scholar] [CrossRef]
- Fusaro, L.; Palma, A.; Salvatori, E.; Basile, A.; Maresca, V.; Karam, E.A.; Manes, F. Functional Indicators of Response Mechanisms to Nitrogen Deposition, Ozone, and Their Interaction in Two Mediterranean Tree Species. PLoS ONE 2017, 12, e0185836. [Google Scholar] [CrossRef]
- Fusaro, L.; Gerosa, G.; Salvatori, E.; Marzuoli, R.; Monga, R.; Kuzminsky, E.; Angelaccio, C.; Quarato, D.; Fares, S. Early and Late Adjustments of the Photosynthetic Traits and Stomatal Density in Q. Uercus Ilex L. Grown in an Ozone-enriched Environment. Plant Biol. J. 2016, 18, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Marzuoli, R.; Monga, R.; Finco, A.; Gerosa, G. Biomass and Physiological Responses of Quercus robur (L.) Young Trees during 2 Years of Treatments with Different Levels of Ozone and Nitrogen Wet Deposition. Trees 2016, 30, 1995–2010. [Google Scholar] [CrossRef]
- Teshome, D.T.; Zharare, G.E.; Naidoo, S. The Threat of the Combined Effect of Biotic and Abiotic Stress Factors in Forestry Under a Changing Climate. Front. Plant Sci. 2020, 11, 601009. [Google Scholar] [CrossRef]
- Ochoa-Hueso, R.; Allen, E.B.; Branquinho, C.; Cruz, C.; Dias, T.; Fenn, M.E.; Manrique, E.; Pérez-Corona, M.E.; Sheppard, L.J.; Stock, W.D. Nitrogen Deposition Effects on Mediterranean-Type Ecosystems: An Ecological Assessment. Environ. Pollut. 2011, 159, 2265–2279. [Google Scholar] [CrossRef] [PubMed]
- Grulke, N.E.; Heath, R.L. Ozone Effects on Plants in Natural Ecosystems. Plant Biol. J. 2020, 22, 12–37. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Parvin, K.; Bhuiyan, T.F.; Anee, T.I.; Nahar, K.; Hossen, M.S.; Zulfiqar, F.; Alam, M.M.; Fujita, M. Regulation of ROS Metabolism in Plants under Environmental Stress: A Review of Recent Experimental Evidence. Int. J. Mol. Sci. 2020, 21, 8695. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, L.; Mereu, S.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Manes, F.; Salvatori, E.; Marzuoli, R.; Gerosa, G.; Tattini, M. Photosynthetic Performance and Biochemical Adjustments in Two Co-Occurring Mediterranean Evergreens, Q. ilex and Arbutus Unedo, Differing in Salt-Exclusion Ability. Funct. Plant Biol. 2014, 41, 391–400. [Google Scholar] [CrossRef]
- Wendling, M.; Büchi, L.; Amossé, C.; Sinaj, S.; Walter, A.; Charles, R. Influence of Root and Leaf Traits on the Uptake of Nutrients in Cover Crops. Plant Soil. 2016, 409, 419–434. [Google Scholar] [CrossRef]
- Sun, Y.; Guo, J.; Li, Y.; Luo, G.; Li, L.; Yuan, H.; Mur, L.A.J.; Guo, S. Negative Effects of the Simulated Nitrogen Deposition on Plant Phenolic Metabolism: A Meta-Analysis. Sci. Total Environ. 2020, 719, 137442. [Google Scholar] [CrossRef]
- Jan, R.; Asaf, S.; Numan, M.; Lubna; Kim, K.-M. Plant Secondary Metabolite Biosynthesis and Transcriptional Regulation in Response to Biotic and Abiotic Stress Conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Liu, W.; Zhu, D.-W.; Liu, D.-H.; Geng, M.-J.; Zhou, W.-B.; Mi, W.-J.; Yang, T.-W.; Hamilton, D. Influence of Nitrogen on the Primary and Secondary Metabolism and Synthesis of Flavonoids in Chrysanthemum Morifolium Ramat. J. Plant Nutr. 2010, 33, 240–254. [Google Scholar] [CrossRef]
- Kováčik, J.; Klejdus, B.; Bačkor, M.; Repčák, M. Phenylalanine Ammonia-Lyase Activity and Phenolic Compounds Accumulation in Nitrogen-Deficient Matricaria Chamomilla Leaf Rosettes. Plant Sci. 2007, 172, 393–399. [Google Scholar] [CrossRef]
- Løvdal, T.; Olsen, K.M.; Slimestad, R.; Verheul, M.; Lillo, C. Synergetic Effects of Nitrogen Depletion, Temperature, and Light on the Content of Phenolic Compounds and Gene Expression in Leaves of Tomato. Phytochemistry 2010, 71, 605–613. [Google Scholar] [CrossRef]
- Bacelar, E.A.; Santos, D.L.; Moutinho-Pereira, J.M.; Lopes, J.I.; Gonçalves, B.C.; Ferreira, T.C.; Correia, C.M. Physiological Behaviour, Oxidative Damage and Antioxidative Protection of Olive Trees Grown under Different Irrigation Regimes. Plant Soil. 2007, 292, 1–12. [Google Scholar] [CrossRef]
- Salbitani, G.; Maresca, V.; Cianciullo, P.; Bossa, R.; Carfagna, S.; Basile, A. Non-Protein Thiol Compounds and Antioxidant Responses Involved in Bryophyte Heavy-Metal Tolerance. Int. J. Mol. Sci. 2023, 24, 5302. [Google Scholar] [CrossRef] [PubMed]
- Schopfer, P.; Liszkay, A.; Bechtold, M.; Frahry, G.; Wagner, A. Evidence That Hydroxyl Radicals Mediate Auxin-Induced Extension Growth. Planta 2002, 214, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-Mediated Abiotic Stress-Induced Programmed Cell Death in Plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef]
- Akhter, R.; Sofi, S.A. Precision Agriculture Using IoT Data Analytics and Machine Learning. J. King Saud. Univ. Comput. Inf. Sci. 2022, 34, 5602–5618. [Google Scholar] [CrossRef]
- Iriti, M.; Faoro, F. Chemical Diversity and Defence Metabolism: How Plants Cope with Pathogens and Ozone Pollution. Int. J. Mol. Sci. 2009, 10, 3371–3399. [Google Scholar] [CrossRef]
- Yeshi, K.; Crayn, D.; Ritmejerytė, E.; Wangchuk, P. Plant Secondary Metabolites Produced in Response to Abiotic Stresses Has Potential Application in Pharmaceutical Product Development. Molecules 2022, 27, 313. [Google Scholar] [CrossRef]
- Sivaranjani, S.; Prasath, V.A.; Pandiselvam, R.; Kothakota, A.; Mousavi Khaneghah, A. Recent Advances in Applications of Ozone in the Cereal Industry. LWT 2021, 146, 111412. [Google Scholar] [CrossRef]
- Joana Gil-Chávez, G.; Villa, J.A.; Fernando Ayala-Zavala, J.; Basilio Heredia, J.; Sepulveda, D.; Yahia, E.M.; González-Aguilar, G.A. Technologies for Extraction and Production of Bioactive Compounds to Be Used as Nutraceuticals and Food Ingredients: An Overview. Compr. Rev. Food Sci. Food Saf. 2013, 12, 5–23. [Google Scholar] [CrossRef]
- Paissoni, M.A.; Río Segade, S.; Giacosa, S.; Torchio, F.; Cravero, F.; Englezos, V.; Rantsiou, K.; Carboni, C.; Gerbi, V.; Teissedre, P.-L.; et al. Impact of Post-Harvest Ozone Treatments on the Skin Phenolic Extractability of Red Winegrapes Cv Barbera and Nebbiolo (Vitis vinifera L.). Food Res. Int. 2017, 98, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Rodoni, L.; Casadei, N.; Concellón, A.; Chaves Alicia, A.R.; Vicente, A.R. Effect of Short-Term Ozone Treatments on Tomato (Solanum lycopersicum L.) Fruit Quality and Cell Wall Degradation. J. Agric. Food Chem. 2010, 58, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, D.W.; Cornic, G. Photosynthetic Carbon Assimilation and Associated Metabolism in Relation to Water Deficits in Higher Plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Cao, X.; Hu, J.; Zhu, L.; Zhang, J.; Huang, J.; Jin, Q. Nitrogen Metabolism in Adaptation of Photosynthesis to Water Stress in Rice Grown under Different Nitrogen Levels. Front. Plant Sci. 2017, 8, 1079. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, H.C.; Eriksson, D.; Møller, I.S.; Schjoerring, J.K. Cytosolic Glutamine Synthetase: A Target for Improvement of Crop Nitrogen Use Efficiency? Trends Plant Sci. 2014, 19, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Rahayu, Y.S.; Walch-Liu, P.; Neumann, G.; Römheld, V.; von Wirén, N.; Bangerth, F. Root-Derived Cytokinins as Long-Distance Signals for NO3−-Induced Stimulation of Leaf Growth. J. Exp. Bot. 2005, 56, 1143–1152. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, B.G.; Falcioni, R.; Guedes, L.M.; Avritzer, S.C.; Antunes, W.C.; Souza, L.A.; Isaias, R.M.S. Preventing False Negatives for Histochemical Detection of Phenolics and Lignins in PEG-Embedded Plant Tissues. J. Histochem. Cytochem. 2017, 65, 105–116. [Google Scholar] [CrossRef]
- Venezia, V.; Verrillo, M.; Avallone, P.R.; Silvestri, B.; Cangemi, S.; Pasquino, R.; Grizzuti, N.; Spaccini, R.; Luciani, G. Waste to Wealth Approach: Improved Antimicrobial Properties in Bioactive Hydrogels through Humic Substance–Gelatin Chemical Conjugation. Biomacromolecules 2023, 24, 2691–2705. [Google Scholar] [CrossRef]
- Verrillo, M.; Koellensperger, G.; Puehringer, M.; Cozzolino, V.; Spaccini, R.; Rampler, E. Evaluation of Sustainable Recycled Products to Increase the Production of Nutraceutical and Antibacterial Molecules in Basil Plants by a Combined Metabolomic Approach. Plants 2023, 12, 513. [Google Scholar] [CrossRef] [PubMed]
- Kovalska, E.; Wu, B.; Liao, L.; Mazanek, V.; Luxa, J.; Marek, I.; Lajaunie, L.; Sofer, Z. Electrochemical Decalcification–Exfoliation of Two-Dimensional Siligene, SixGey: Material Characterization and Perspectives for Lithium-Ion Storage. ACS Nano 2023, 17, 11374–11383. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of Water Stress-Induced Changes in the Levels of Endogenous Ascorbic Acid and Hydrogen Peroxide in Vigna Seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Peng, X.; Wang, N.; Sun, S.; Geng, L.; Guo, N.; Liu, A.; Chen, S.; Ahammed, G.J. Reactive Oxygen Species Signaling Is Involved in Melatonin-Induced Reduction of Chlorothalonil Residue in Tomato Leaves. J. Hazard. Mater. 2023, 443, 130212. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verrillo, M.; Cianciullo, P.; Cozzolino, V.; De Ruberto, F.; Maresca, V.; Di Fraia, A.; Fusaro, L.; Manes, F.; Basile, A. Oxidative Stress Response Mechanisms Sustain the Antibacterial and Antioxidant Activity of Quercus ilex. Plants 2024, 13, 1154. https://doi.org/10.3390/plants13081154
Verrillo M, Cianciullo P, Cozzolino V, De Ruberto F, Maresca V, Di Fraia A, Fusaro L, Manes F, Basile A. Oxidative Stress Response Mechanisms Sustain the Antibacterial and Antioxidant Activity of Quercus ilex. Plants. 2024; 13(8):1154. https://doi.org/10.3390/plants13081154
Chicago/Turabian StyleVerrillo, Mariavittoria, Piergiorgio Cianciullo, Vincenza Cozzolino, Francesca De Ruberto, Viviana Maresca, Alessia Di Fraia, Lina Fusaro, Fausto Manes, and Adriana Basile. 2024. "Oxidative Stress Response Mechanisms Sustain the Antibacterial and Antioxidant Activity of Quercus ilex" Plants 13, no. 8: 1154. https://doi.org/10.3390/plants13081154