
Effects of Salt Stress on Physiological and Agronomic Traits of Rice Genotypes with Contrasting Salt Tolerance


 ,
,
Abstract
:1. Introduction
2. Results
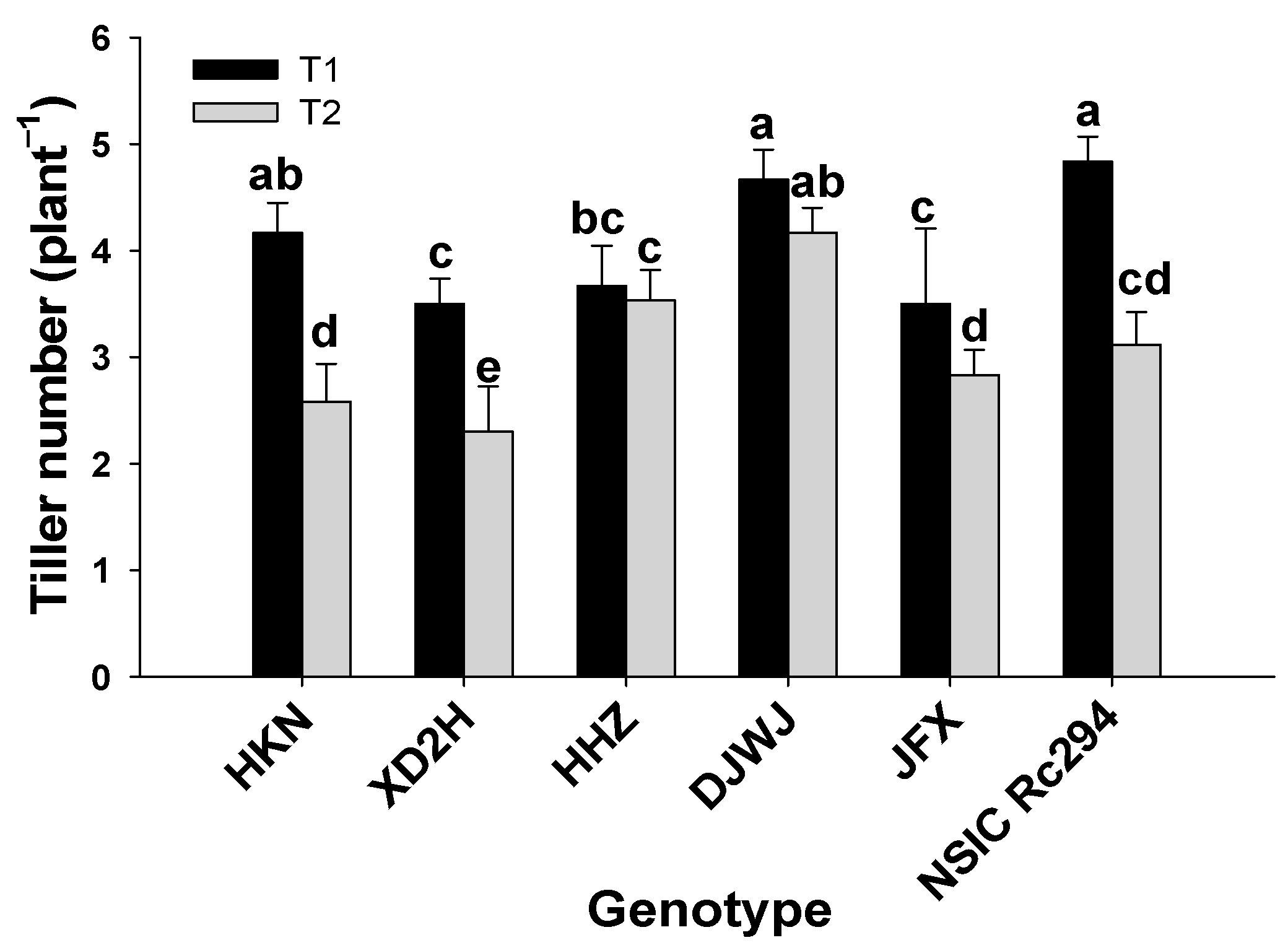
2.1. Growth Characteristics of Different Rice Genotypes under Salt Stress
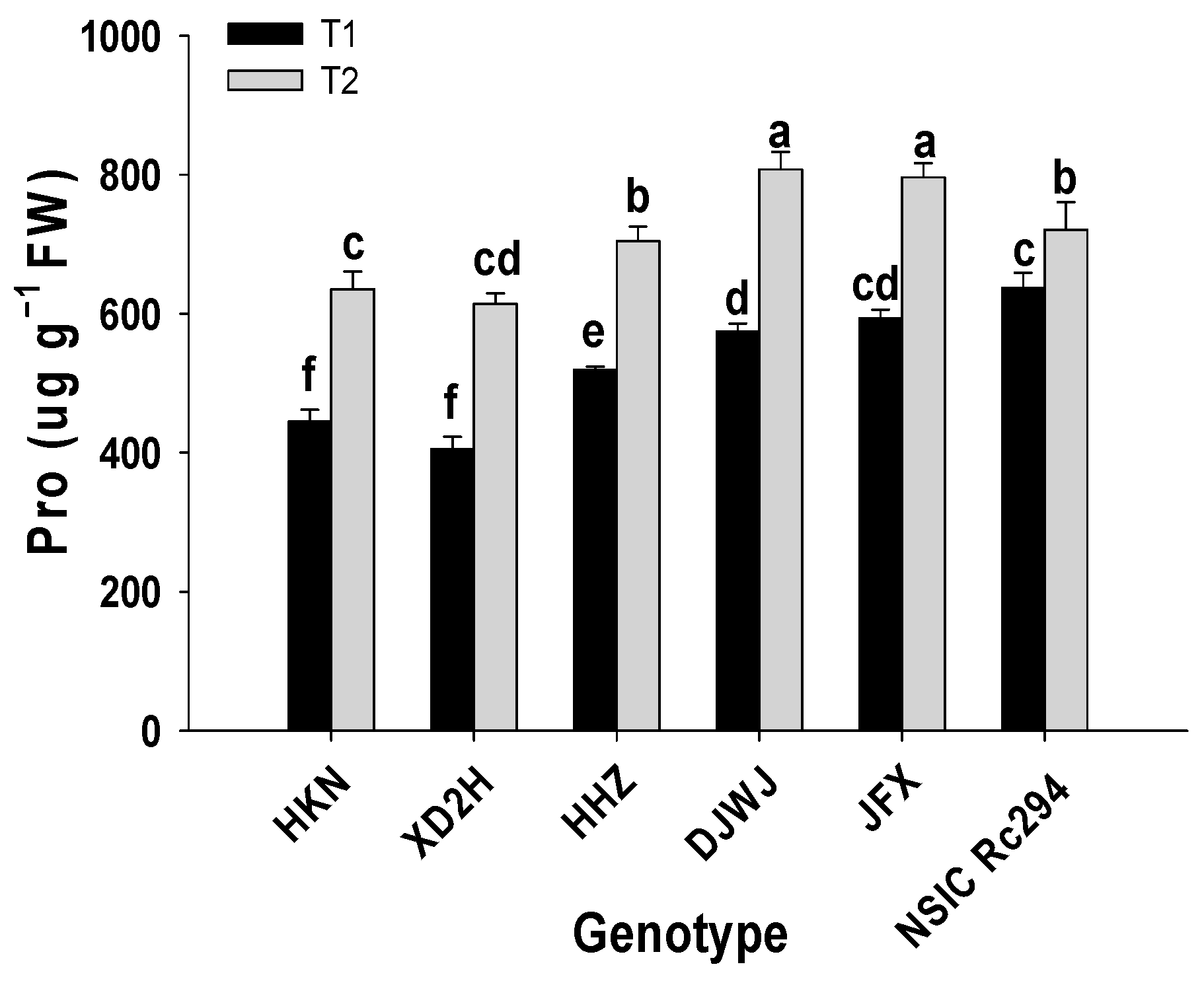
2.2. Physiological Characteristics of Different Rice Genotypes under Salt Stress
2.2.1. Membrane Lipid Peroxidation
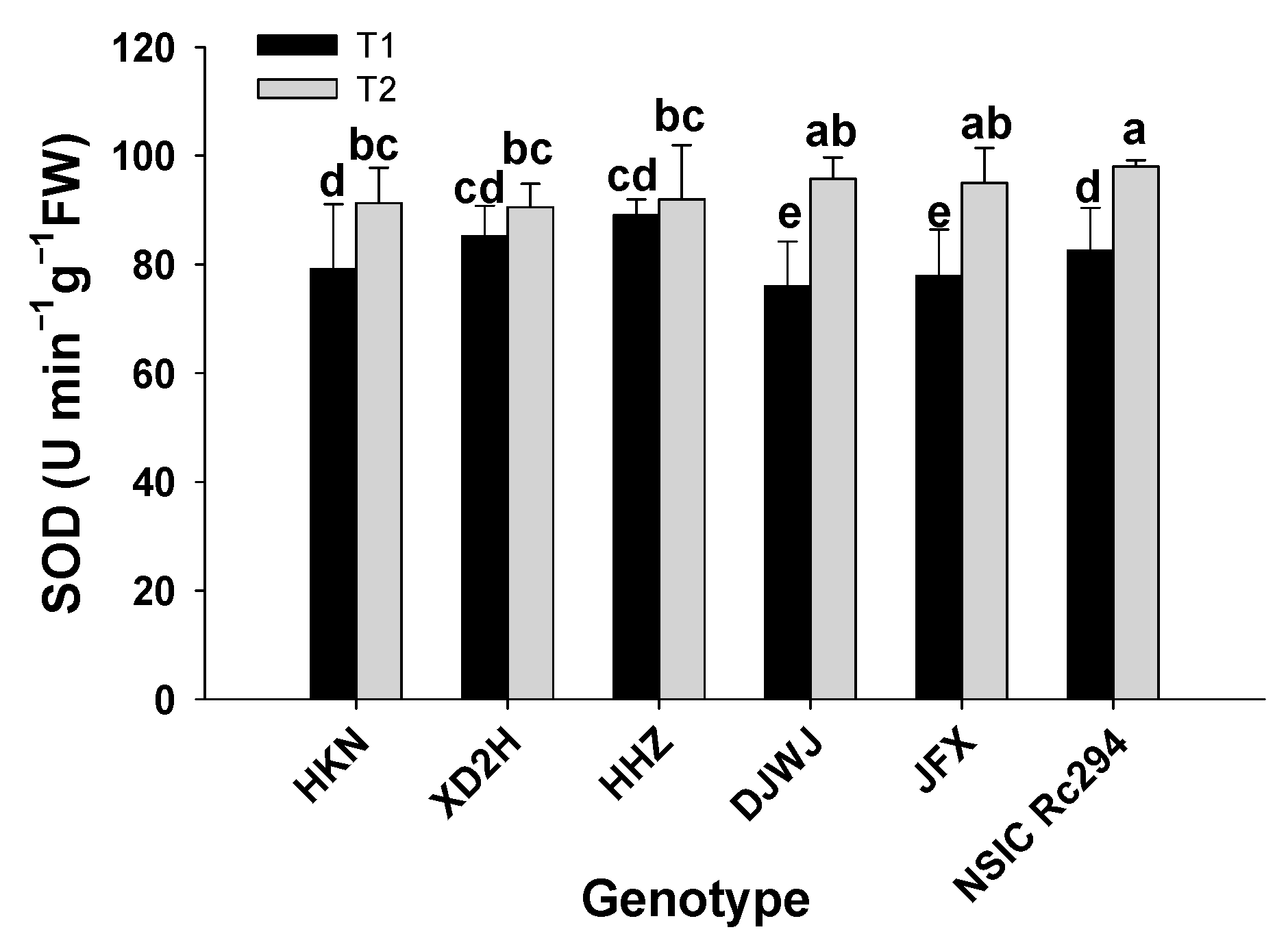
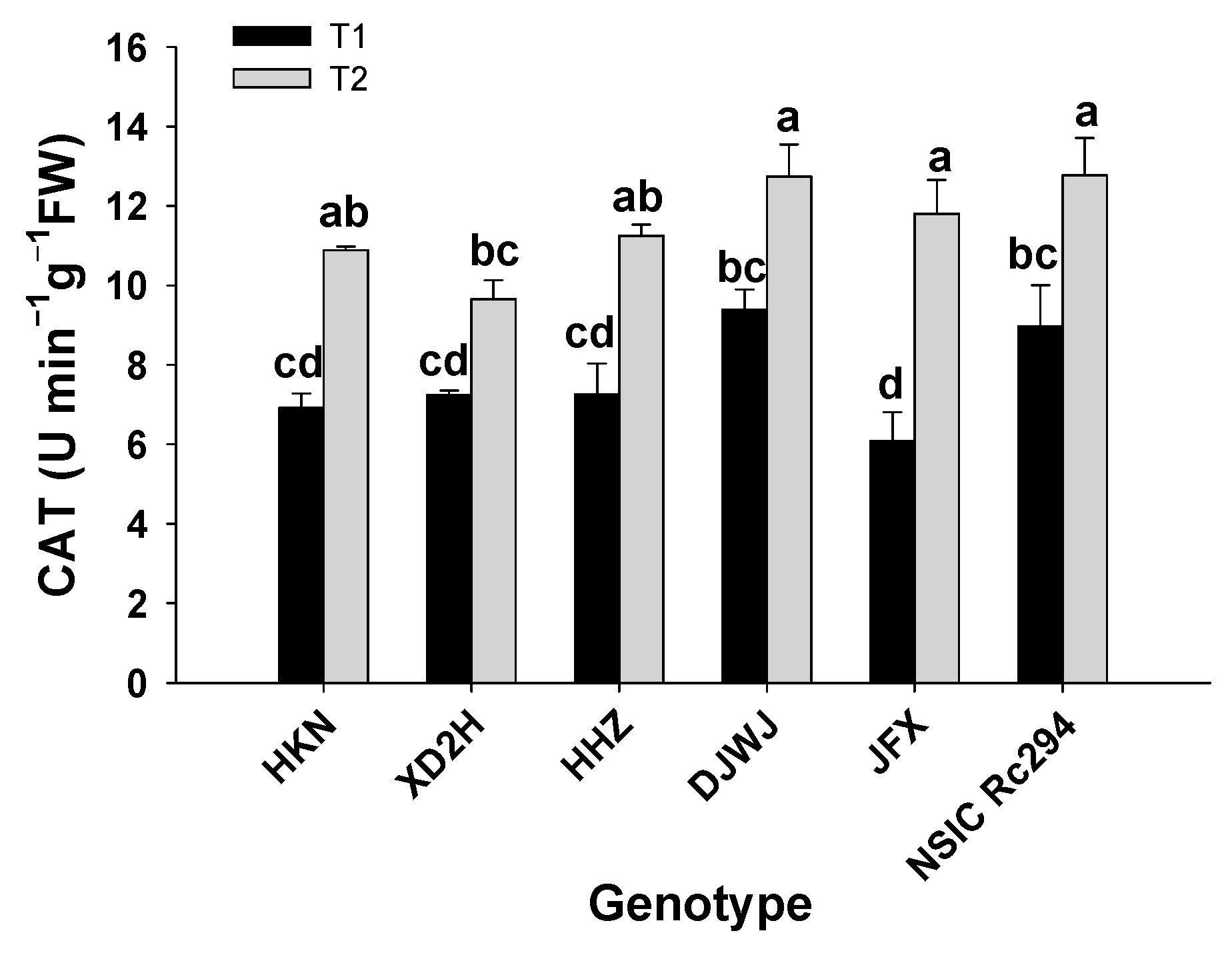
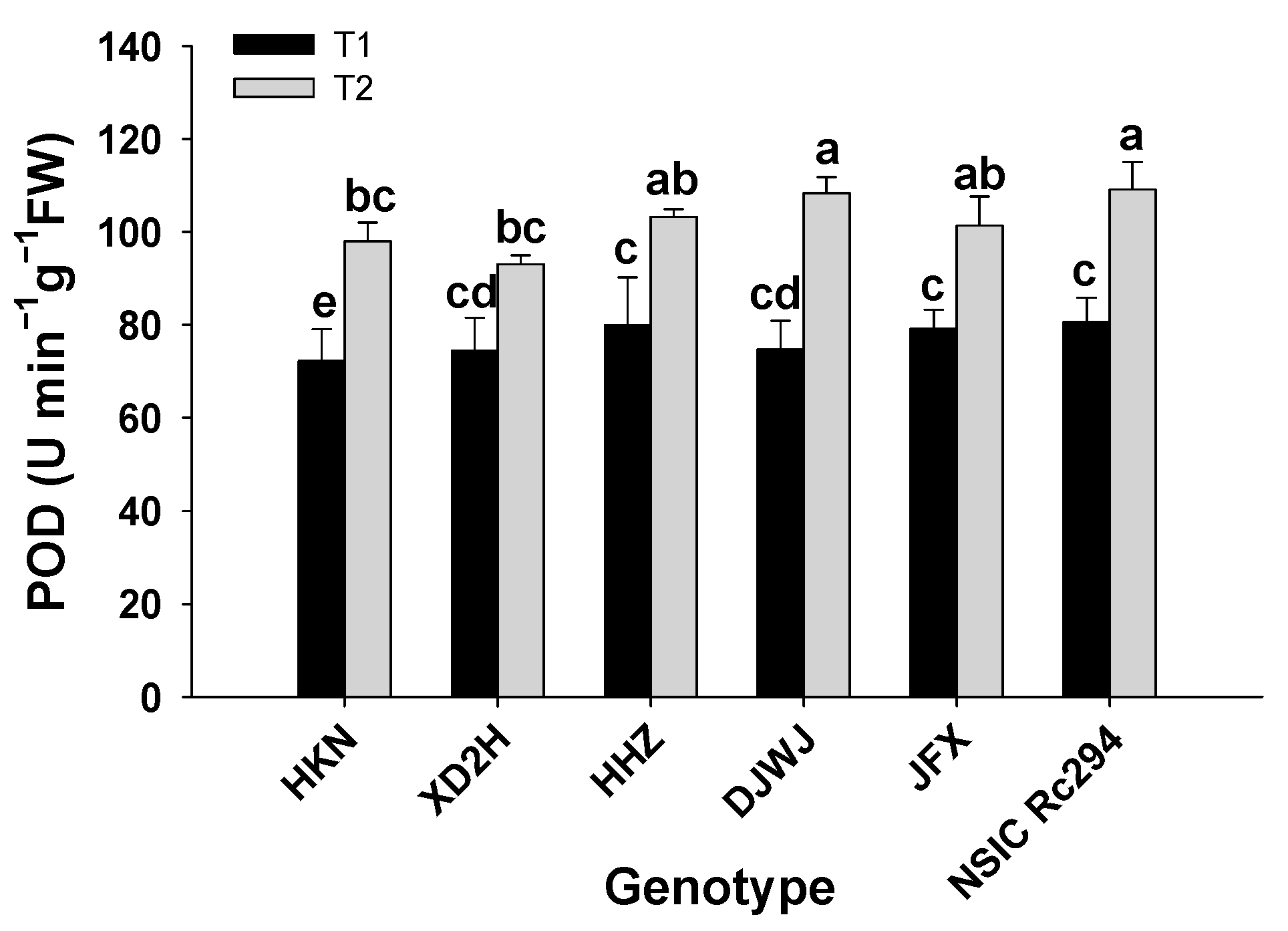
2.2.2. Antioxidant Enzyme Activity
2.2.3. Osmoregulatory Substances
2.2.4. Na+, K+ and Na+/K+
2.3. Yield and Yield Components of Different Rice Genotypes under Salt Stress
2.4. Correlation Analysis
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Growth Parameter Measurement
4.3. Physiological Parameter Measurements
4.4. Yield and Yield Component Measurements
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, G.; Xu, Y.; Xu, Z.; Ahmad, I.; Nimir, N.E.A.; Zhou, G. Improving productivity of Sesbania pea in saline soils by enhancing antioxidant capacity with optimum application of nitrogen and phosphate combination. Front. Plant Sci. 2022, 13, 1027227. [Google Scholar] [CrossRef] [PubMed]
- Sadak, M.S.; Dawood, M.G. Biofertilizer Role in alleviating the deleterious effects of salinity on wheat growth and productivity. Gesunde Pflanzen 2023, 75, 1207–1219. [Google Scholar] [CrossRef]
- Zhu, G.; Lu, H.; Shi, X.; Wang, Y.; Zhou, G. Nitrogen management enhanced plant growth, antioxidant ability, and grain yield of rice under salinity stress. Agron. J. 2020, 112, 550–563. [Google Scholar] [CrossRef]
- Zhen, Y.; Wang, B. Present status of saline soil resources and countermeasures for improvement and utilization in China. Shandong Agric. Sci. 2015, 47, 125–130. [Google Scholar] [CrossRef]
- Zhu, G.; Wu, H.; Chen, Y.; Mondal, S.; Ismail, A.M. Growth characteristics and yield of contrasting rice genotypes under long-term stagnant flooding. Field Crops Res. 2023, 301, 109020. [Google Scholar] [CrossRef]
- National Bureau of Statistics, China. 2023. Available online: https://data.stats.gov.cn/easyquery.htm?cn=C01 (accessed on 10 April 2023).
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Ann. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Joseph, B.; Jini, D.; Sujatha, S. Biological and physiological perspectives of specificity in abiotic salt stress response from various rice plants. Asian J. Agric. Sci. 2010, 2, 99–105. [Google Scholar]
- Prasad, S.R.; Bagali, P.G.; Hittalmani, S.; Shashidhar, H. Molecular mapping of quantitative trait loci associated with seedling tolerance to salt stress in rice (Oryza sativa L.). Curr. Sci. India 2000, 78, 162–164. [Google Scholar]
- Islam, M.Z.; Sattar, M.A.; Ashrafuzzaman, M.; Zulkerami, B.; Shamsuddoha, A.T.M. Evaluating some salinity tolerant rhizobacterial strains to lentil production under salinity stress. Int. J. Agric. Biol. 2013, 15, 499–504. [Google Scholar]
- Ismail, A.M.; Horie, T. Genomics, physiology, and molecular breeding approaches for improving salt tolerance. Annu. Rev. Plant Biol. 2017, 68, 405–434. [Google Scholar] [CrossRef]
- Ismail, A.M.; Heuer, S.; Thomson, M.J.; Wissuwa, M. Genetic and genomic approaches to develop rice germplasm for problem soils. Plant Mol. Biol. 2007, 65, 547–570. [Google Scholar] [CrossRef] [PubMed]
- Sadak, M.S.; Sekara, A.; Al-ashkar, I.; Habib-ur-Rahman, M.; Skalicky, M.; Brestic, M.; Kumar, A.; Sabagh, A.E.; Abdelhamid, M.T. Exogenous aspartic acid alleviates salt stress induced decline in growth by enhancing antioxidants and compatible solutes while reducing reactive oxygen species in wheat. Front. Plant Sci. 2022, 13, 987641. [Google Scholar] [CrossRef] [PubMed]
- Sadak, M.S.; Abd El-Monem, A.A.; El-Bassiouny, H.M.S.; Badr, N.M. Physiological response of sunflower (Helianthus annuus L.) to exogenous arginine and putrescine treatments under salinity Stress. J. Appl. Sci. Res. 2012, 8, 4943–4957. [Google Scholar]
- Meng, T.Y.; Zhang, X.B.; Ge, J.L.; Chen, X.; Yang, Y.L.; Zhu, G.L.; Chen, Y.L.; Zhou, G.S.; Wei, H.H.; Dai, Q.G. Agronomic and physiological traits facilitating better yield performance of japonica/indica hybrids in saline fields. Field Crops Res. 2021, 271, 108255. [Google Scholar] [CrossRef]
- Yang, W.; Gao, Y.; Wang, X.; Li, S.; Zheng, H.; Chen, Z.; Wu, F.; Du, X.; Sui, N. Exogenous calcium application enhances salt tolerance of sweet sorghum seedlings. J. Agron. Crop Sci. 2022, 208, 441–453. [Google Scholar] [CrossRef]
- Zheng, Y.J. Effect of salt stress on rice and research on salt tolerant breeding. North Rice 2013, 43, 71–74, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Kibria, M.G.; Hossain, M.; Murata, Y.; Hoque, M.A. Antioxidant defense mechanisms of salinity tolerance in rice genotypes. Rice Sci. 2017, 24, 155–162. [Google Scholar] [CrossRef]
- Jini, D.; Joseph, B. Physiological mechanism of salicylic acid for alleviation of salt stress in rice. Rice Sci. 2017, 24, 97–108. [Google Scholar] [CrossRef]
- Ahmad, I.; Zhu, G.; Zhou, G.; Song, X.; Hussein, I.M.E.; Ibrahim, S.E.G. Effect of N on growth, antioxidant capacity, and chlorophyll content of sorghum. Agronomy 2022, 12, 501. [Google Scholar] [CrossRef]
- Mohanty, A.; Chakraborty, K.; Mondal, S.; Jena, P.; Panda, R.K.; Samal, K.C.; Chattopadhyay, K. Relative contribution of ion exclusion and tissue tolerance traits govern the differential response of rice towards salt stress at seedling and reproductive stages. Environ. Exp. Bot. 2023, 206, 105131. [Google Scholar] [CrossRef]
- Zahra, J.; Nazim, H.; Cai, S.G.; Han, Y.; Wu, D.Z.; Zhang, B.L.; Haider, S.I.; Zhang, G.P. The influence of salinity on cell ultrastructures and photosynthetic apparatus of barley genotypes differing in salt stress tolerance. Acta Physiol. Plant 2014, 36, 1261–1269. [Google Scholar] [CrossRef]
- Roy, C.A.; Basu, S. Over expression of an abiotic stress responsive plant protein in bacteria E. coli. Afr. J. Plant Biotechnol. 2008, 7, 3231–3234. [Google Scholar]
- Dai, J.Y.; Zhang, G.Y.; Hu, L.; Sun, H.Q.; Wan, L.S.; Han, P.P.; Ni, Z.B. Grey correlation analysis of the relationship between the yield and main agronomic characters of salt-tolerant rice varieties. Barl. Cereal Sci. 2020, 37, 9–13, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Cha-Um, S.; Charoenpanich, A.; Roytrakul, S.; Kirdmanee, C. Sugar accumulation, photosynthesis and growth of two indica rice varieties in response to salt stress. Acta Physiol. Plant 2009, 31, 477–486. [Google Scholar] [CrossRef]
- Kanawapee, N.; Sanitchon, J.; Srihaban, P.; Theerakulpisut, P. Physiological changes during development of rice (Oryza sativa L.) varieties differing in salt tolerance under saline field condition. Plant Soil 2013, 370, 89–101. [Google Scholar] [CrossRef]
- Xie, Z.Y.; Wang, C.C.; Zhu, S.B.; Wang, W.S.; Xu, J.L.; Zhao, X.Q. Characterizing the metabolites related to rice salt tolerance with introgression lines exhibiting contrasting performances in response to saline conditions. Plant Growth Regul. 2020, 92, 157–176. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, M.; Guo, R.; Shi, D.; Liu, B.; Lin, X.; Yang, C. Effects of salt stress on ion balance and nitrogen metabolism of old and young leaves in rice (Oryza sativa L.). BMC Plant Biol. 2012, 12, 194. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.L.; Wang, Y.; Shi, X.X.; Lu, H.T.; Ren, Z.; Shi, Y.; Jiao, X.R.; Ibrahim, M.E.H.; Irshad, A.; Zhu, W.B.; et al. Optimum nitrogen management enhances growth, antioxidant ability and yield performance of rice in saline soil of coastal area of China. Chilean J. Agric. Res. 2020, 80, 629–639. [Google Scholar] [CrossRef]
- Platten, J.D.; Egdane, J.A.; Ismail, A.M. Salinity tolerance, Na+ exclusion and allele mining of HKT1;5 in Oryza sativa and O. glaberrima: Many sources, many genes, one mechanism? BMC Plant Biol. 2013, 13, 32. [Google Scholar] [CrossRef]
- Yang, J.Q.; Zhang, J.H. Crop management techniques to enhance harvest index in rice. J. Exp. Bot. 2012, 61, 3177–3189. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Kumari, S.; Nazir, F.; Khanna, R.R.; Gupta, R.; Chhillar, H. Defensive role of plant hormones in advancing abiotic stress-resistant rice plants. Rice Sci. 2023, 30, 15–35. [Google Scholar] [CrossRef]
- Zhu, G.; Peng, S.; Huang, J.; Cui, K.; Nie, L.; Wang, F. Genetic improvements in rice yield and concomitant increases in radiation- and nitrogen-use efficiency in middle reaches of Yangtze River. Sci. Rep. 2016, 6, 21049. [Google Scholar] [CrossRef] [PubMed]
- Abeer, A.R.; Fatma, A.F.; Afaf, M.H. Physiological and biochemical responses of salt-tolerant and salt-sensitive wheat and bean cultivars to salinity. J. Biol. Earth Sci. 2013, 3, 72–88. [Google Scholar]
- Gerona, M.E.B.; Deocampo, M.P.; Egdane, J.A.; Ismail, A.M.; Dionisio-Sese, M.L. Physiological responses of contrasting rice genotypes to salt stress at reproductive stage. Rice Sci. 2019, 26, 207–219. [Google Scholar] [CrossRef]
- Wasinee, P.; Chakkree, L.; Sarunyaporn, M. Evaluation and clustering of rice cultivars tolerance to salt and drought stresses using morpho-physiological and biochemical indices. J. Crop Sci. Biotechnol. 2023, 26, 573–584. [Google Scholar] [CrossRef]
- Saddiq, M.S.; Afzal, I.; Basra, S.M.A.; Ali, Z.; Ibrahim, A.M.H. Sodium exclusion is a reliable trait for the improvement of salinity tolerance in bread wheat. Arch. Agron. Soil Sci. 2017, 64, 272–284. [Google Scholar] [CrossRef]
- Cervilla, L.M.; Blasco, B.; Ríos, J.J.; Romero, L.; Ruiz, J.M. Oxidative stress and antioxidants in tomato plants subjected to boron toxicity. Ann. Bot. 2007, 100, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Frukh, A.; Siddiqi, T.O.; Khan, M.I.R.; Ahmad, A. Modulation in growth, biochemical attributes and proteome profile of rice cultivars under salt stress. Plant Physiol. Bio. 2020, 146, 55–70. [Google Scholar] [CrossRef]
- Zhao, D.Y.; Zhang, Z.W.; Liu, G.L.; Zhao, S.P.; Zhang, X.L.; Zhao, W.F.; Siddique, K.H.M. Effects of phosphorus supply and salt stress on plant growth, organic acid concentration, and sodium, potassium, calcium, magnesium, and phosphorus utilization in wheat. Cereal Res. Commun. 2023, 51, 905–915. [Google Scholar] [CrossRef]
- Huang, C.C.; Lee, Y.T.; Ly, T.T.; Wang, C.Y.; Chang, Y.T.; Hou, P.F. Volatile cinnamaldehyde induces systemic salt tolerance in the roots of rice (Oryza sativa). Physiol. Plant. 2023, 175, e13938. [Google Scholar] [CrossRef]
- Zhu, G.; Chen, Y.; Ella, E.S.; Ismail, A.M. Mechanisms associated with tiller suppression under stagnant flooding in rice. J. Agron. Crop Sci. 2019, 205, 235–247. [Google Scholar] [CrossRef]
- Assaha, D.V.M.; Mekawy, A.M.M.; Liu, L.; Noori, M.S.; Kokulan, K.S.; Ueda, A.; Nagaoka, T.; Saneoka, H. Na+ retention in the root is a key adaptive mechanism to low and high salinity in the glycophyte, Talinum paniculatum (Jacq.) gaertn. (Portulacaceae). J. Agron. Crop Sci. 2017, 203, 56–67. [Google Scholar] [CrossRef]
- Zhou, G.S.; Nimir, N.; Lu, S.Y.; Zhai, F.Y.; Wang, Y.H. Gibberellic acid and salinity affected growth and antioxidant enzyme activities in castor bean plants at early growth stage. Agron. J. 2014, 106, 1340–1348. [Google Scholar] [CrossRef]
- Seckin, B.; Sekmen, A.H.; Türkan, S. An enhancing effect of exogenous mannitol on the antioxidant enzyme activities in roots of wheat under salt stress. J. Plant Growth Regul. 2009, 28, 12–20. [Google Scholar] [CrossRef]
- Chen, Y.L.; Li, R.K.; Ge, J.F.; Liu, J.G.; Wang, W.B.; Xu, M.F.; Hussain, S.; Wei, H.H.; Dai, Q.G. Exogenous melatonin confers enhanced salinity tolerance in rice by blocking the ROS burst and improving Na+/K+ homeostasis. Environ. Exp. Bot. 2021, 189, 104530. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Treatment | Fresh Weight (g Plant−1) | Dry Weight (g Plant−1) | RGR (g Plant−1 d−1) |
|---|---|---|---|---|
| HKN | T1 | 74.8 e | 5.8 f | 0.11 f |
| T2 | 31.7 h | 2.8 g | 0.05 g | |
| XD2H | T1 | 46.5 g | 8.5 cd | 0.16 c |
| T2 | 36.0 h | 3.1 g | 0.06 g | |
| HHZ | T1 | 106.8 d | 10.1 b | 0.19 b |
| T2 | 59.7 f | 6.3 f | 0.12 f | |
| DJWJ | T1 | 172.7 a | 10.0 b | 0.19 b |
| T2 | 96.1 d | 9.0 c | 0.17 c | |
| JFX | T1 | 123.6 c | 10.7 ab | 0.20 b |
| T2 | 65.3 ef | 7.0 e | 0.13 e | |
| NSIC Rc294 | T1 | 156.9 b | 11.8 a | 0.22 a |
| T2 | 67.7 ef | 7.9 d | 0.15 d | |
| LSD (0.05) | 52.678 | 3.350 | 0.035 | |
| T | *** | *** | *** | |
| V | *** | *** | *** | |
| T × V | ** | ns | ns |
| Variety | Treatment | Na+ (mg g−1 DW) | K+ (mg g−1 DW) | Na+/K+ (mg g−1 DW) |
|---|---|---|---|---|
| HKN | T1 | 1.06 cd | 20.0 c | 0.053 f |
| T2 | 5.93 a | 14.1 f | 0.421 c | |
| XD2 H | T1 | 0.98 d | 19.2 cd | 0.051 f |
| T2 | 5.57 a | 11.4 g | 0.489 b | |
| HHZ | T1 | 1.09 cd | 20.6 bc | 0.053 f |
| T2 | 5.43 ab | 10.8 g | 0.503 a | |
| DJWJ | T1 | 0.85 d | 21.1 abc | 0.040 g |
| T2 | 4.81 bc | 17.5 de | 0.275 d | |
| JFX | T1 | 0.90 d | 23.2 a | 0.039 g |
| T2 | 4.20 c | 16.0 ef | 0.263 e | |
| NSIC | T1 | 0.90 d | 22.3 ab | 0.040 g |
| T2 | 4.49 c | 17.3 de | 0.260 h | |
| LSD (0.05) | 0.64 | 2.19 | 4.98 | |
| T | *** | *** | *** | |
| V | ** | *** | * | |
| T × V | * | * | ns |
| Variety | Treatment | Yield (g pot−1) | Panicles (pot−1) | Spikelets Per Panicle | Total Spikelets (103 pot−1) | Grain Filling (%) | 1000-Grain Weight (g) |
|---|---|---|---|---|---|---|---|
| HKN | T1 | 145.6 a | 54.1 c | 178.7 a | 9.65 a | 67.82 d | 22.2 a |
| T2 | 52.9 g | 39.3 e | 118.3 c | 4.65 d | 57.04 e | 20.2 c | |
| XD2H | T1 | 77.1 f | 52.4 c | 92.8 d | 4.86 d | 76.29 b | 20.9 c |
| T2 | 32.2 h | 44.1 d | 61.3 f | 2.70 f | 62.95 d | 19.0 d | |
| HHZ | T1 | 81.9 e | 77.5 a | 79.6 e | 6.17 c | 72.24 c | 18.4 e |
| T2 | 38.4 h | 59.8 c | 62.0 f | 3.71 e | 61.86 d | 17.1 f | |
| DJWJ | T1 | 102.6 d | 77.5 a | 89.8 d | 6.96 c | 73.09 bc | 20.2 c |
| T2 | 54.8 g | 61.5 b | 69.2 f | 4.26 d | 66.50 d | 19.5 d | |
| JFX | T1 | 111.8 c | 70.2 ab | 94.1 d | 6.61 c | 75.68 b | 22.5 a |
| T2 | 58.7 g | 49.5 d | 74.6 e | 3.69 e | 70.84 c | 22.4 a | |
| NSIC | T1 | 134.4 b | 65.3 b | 134.9 b | 8.81 b | 84.34 a | 21.0 b |
| T2 | 87.9 e | 53.4 c | 93.6 d | 5.00 d | 75.92 b | 20.2 c | |
| LSD (0.05) | 19.5 | 6.2 | 15.1 | 950.1 | 0.06 | 1.2 | |
| T | *** | *** | *** | *** | *** | ** | |
| V | *** | *** | *** | *** | *** | *** | |
| T × V | * | ns | ** | *** | * | ns |
| Yield | Panicles | Spikelets Per Panicle | Total Spikelets | Grain Filling | 1000-Grain Weight | |
|---|---|---|---|---|---|---|
| PH | 0.3174 ns | 0.6338 *** | −0.1033 ns | 0.2513 ns | 0.3287 ns | 0.2793 ns |
| FW | 0.6006 ** | 0.8373 *** | 0.1483 ns | 0.6154 ** | 0.5610 ** | 0.1410 ns |
| DW | 0.5327 ** | 0.7736 *** | 0.0216 ns | 0.4727 * | 0.7743 *** | 0.1623 ns |
| TN | 0.5801 ** | 0.6478 *** | 0.0791 ns | 0.2488 ns | 0.2471 ns | −0.0412 ns |
| SOD | −0.5447 ** | −0.2332 ns | −0.4926 * | −0.5818 ** | −0.2172 ns | −0.2965 ns |
| POD | −0.6274 *** | −0.6115 ** | −0.4426 * | −0.7003 *** | −0.2726 ns | −0.1744 ns |
| CAT | −0.5888 ** | −0.4629 * | −0.2969 ns | −0.5259 ** | −0.4962 * | −0.6022 ** |
| MDA | −0.7049 *** | −0.4687 * | −0.3589 ns | −0.5932 ** | −0.6914 *** | −0.5011 * |
| SPT | −0.7677 *** | −0.2788 ns | −0.5846 ** | −0.6963 *** | −0.5606 ** | −0.6321 *** |
| Pro | 0.4412 * | 0.1342 ns | 0.4703 * | 0.4843 * | 0.1604 ns | 0.1432 ns |
| K+ | 0.8067 *** | 0.6207 ** | 0.451 * | 0.749 *** | 0.7026 *** | 0.5445 ** |
| Na+ | −0.7974 *** | −0.6494 *** | −0.451 * | −0.7733 *** | −0.6597 *** | −0.4219 * |
| Variety | Abbreviation | Type | Salt Tolerance |
|---|---|---|---|
| Hongkenuo | HKN | japonica | salt-sensitive |
| Xudao2hao | XD2H | japonica | salt-sensitive |
| Huanghuazhan | HHZ | indica | salt-sensitive |
| Dijiaowujian | DJWJ | indica | salt-tolerant |
| Jiefangxian | JFX | indica | salt-tolerant |
| NSIC Rc294 | NSIC | indica | salt-tolerant |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Bu, W.; Xu, Y.; Fei, H.; Zhu, Y.; Ahmad, I.; Nimir, N.E.A.; Zhou, G.; Zhu, G. Effects of Salt Stress on Physiological and Agronomic Traits of Rice Genotypes with Contrasting Salt Tolerance. Plants 2024, 13, 1157. https://doi.org/10.3390/plants13081157
Xu Y, Bu W, Xu Y, Fei H, Zhu Y, Ahmad I, Nimir NEA, Zhou G, Zhu G. Effects of Salt Stress on Physiological and Agronomic Traits of Rice Genotypes with Contrasting Salt Tolerance. Plants. 2024; 13(8):1157. https://doi.org/10.3390/plants13081157
Chicago/Turabian StyleXu, Yunming, Weicheng Bu, Yuchao Xu, Han Fei, Yiming Zhu, Irshad Ahmad, Nimir Eltyb Ahmed Nimir, Guisheng Zhou, and Guanglong Zhu. 2024. "Effects of Salt Stress on Physiological and Agronomic Traits of Rice Genotypes with Contrasting Salt Tolerance" Plants 13, no. 8: 1157. https://doi.org/10.3390/plants13081157