
Impacts of Soil Properties on Species Diversity and Structure in Alternanthera philoxeroides-Invaded and Native Plant Communities

1
College of Life Sciences, Xinyang Normal University, Xinyang 464000, China
2
Henan Dabieshan National Field Observation and Research Station of Forest Ecosystem, Zhengzhou 450046, China
3
Xinyang Academy of Ecological Research, Xinyang 464000, China
4
Dabie Mountain Laboratory, Xinyang 464000, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Plants 2024, 13(9), 1196; https://doi.org/10.3390/plants13091196
Submission received: 18 March 2024
/
Revised: 21 April 2024
/
Accepted: 24 April 2024
/
Published: 25 April 2024
(This article belongs to the Special Issue Interactions within Invasive Ecosystems)
Abstract
:Soil properties can affect plant population dynamics and the coexistence of native and invasive plants, thus potentially affecting community structure and invasion trends. However, the different impacts of soil physicochemical properties on species diversity and structure in native and invaded plant communities remain unclear. In this study, we established a total of 30 Alternanthera philoxeroides-invaded plots and 30 control plots in an area at the geographical boundary between North and South China. We compared the differences in species composition between the invaded and native plant communities, and we then used the methods of regression analysis, redundancy analysis (RDA), and canonical correspondence analysis (CCA) to examine the impacts of soil physicochemical properties on four α-diversity indices and the species distribution of these two types of communities. We found that A. philoxeroides invasion increased the difference between the importance values of dominant plant species, and the invasion coverage had a negative relationship with the soil-available potassium (R2 = 0.135; p = 0.046) and Patrick richness index (R2 = 0.322; p < 0.001). In the native communities, the species diversity was determined with soil chemical properties, the Patrick richness index, the Simpson dominance index, and the Shannon–Wiener diversity index, which all decreased with the increase in soil pH value, available potassium, organic matter, and ammonium nitrogen. However, in the invaded communities, the species diversity was determined by soil physical properties; the Pielou evenness index increased with increasing non-capillary porosity but decreased with increasing capillary porosity. The determinants of species distribution in the native communities were soil porosity and nitrate nitrogen, while the determinants in the invaded communities were soil bulk density and available potassium. In addition, compared with the native communities, the clustering degree of species distribution in the invaded communities intensified. Our study indicates that species diversity and distribution have significant heterogeneous responses to soil physicochemical properties between A. philoxeroides-invaded and native plant communities. Thus, we need to intensify the monitoring of soil properties in invaded habitats and conduct biotic replacement strategies based on the heterogeneous responses of native and invaded communities to effectively prevent the biotic homogenization that is caused by plant invasions under environmental changes.
1. Introduction
Plant invasions seriously threaten the function and biodiversity of the global ecosystem [1] and cause the loss of native functional groups [2]. Alien plants can successfully invade due to their strong phenotypic plasticity and interspecific competitiveness and through interactions with soil, microorganisms, and native plants [3,4]. For example, the widespread invasion of Solidago canadensis, Solanum elaeagnifolium, and Ageratina adenophora has significantly decreased the species diversity of native plant communities [5,6]. The invasive species Phragmites australis in Australia weakens the competitiveness of native plants, and plant diversity decreases with increasing P. australis density [7]. The invasive species Imperata cylindrica has a higher water use efficiency than the accompanying species, thus inhibiting the latter’s growth and reproduction [2]. Increased Lupinus polyphyllus invasion coverage intensifies the biohomogenization of native plant communities [8]. Impatiens glandulifera can enhance its invasive competitiveness by reducing the number of beneficial epiphytic fungi on native accompanying plants [9]. Due to its great niche breadth, long flowering period, large seed quantity, and high root–shoot ratio, Gaillardia aristata can transform from an alien ornamental species into an invader under stressful environmental conditions, and it decreases the plant richness and diversity indices in invaded habitats [10]. Thus, exploring the correlation between plant invasions and native species diversity, as well as their environmental determinants, is beneficial for predicting community succession dynamics and elucidating species coexistence mechanisms in invaded ecosystems.
Soil is the main source of plant nutrition, and its physicochemical properties and resource availability affect plant physiological and ecological characteristics [11], which further affect plant population dynamics and species diversity [12,13]. The spread of invasive plants and their effects on native communities are also regulated by soil properties [14,15,16]. For example, high organic matter content in soil can increase the population density and root numbers of the invasive species Ludwigia grandiflora and thus exacerbate its invasion process [17]. High soil nitrogen can increase the abundance, height, biomass, and seed quantity of the invasive species Mikania micrantha and Bidens pilosa, thereby weakening native plant diversity [18,19]. Heterogeneous soil nutrients can promote the aboveground biomass accumulation of the invasive species Ageratum conyzoides, B. pilosa, and Erigeron annuus [13]. Plant invasions can also alter the soil conditions in microhabitats. For example, Chromolaena odorata invasion causes a significant increase in soil nitrogen, available phosphorus, and pH value, which promote the biomass accumulation of subsequent invasive plants, although this promoting effect is weakened by the increase in native plant richness [16]. Agerina adenophora invasion leads to an increase in soil nitrogen and a decrease in pH, which inhibit the seed germination and seedling growth of native plants [20]. However, studies that examine the impacts of soil physicochemical properties on invaded and native plant communities at the same spatiotemporal scale are relatively rare.
Alternanthera philoxeroides, also known as alligator weed, a noxious invasive weed native to South America belonging to the Amaranthaceae family, grows in both aquatic and terrestrial environments [21]. A. philoxeroides has a robust asexual reproductive ability, which enables it to propagate asexually through the fracture of stem segments and adventitious roots on stem nodes, and thus, it rapidly spreads and establishes populations across diverse habitats [21]. Furthermore, A. philoxeroides can produce a variety of secondary metabolites, including alkaloids, organic acids, and flavonoids, to inhibit the growth of other accompanying plants, providing favorable conditions for its diffusion and establishment [22]. Due to its low genetic diversity, strong phenotypic plasticity, and fast growth rate, A. philoxeroides has invaded various parts of the world, including the United States, China, South Africa, India, and Australia, causing damage to the ecological environment, biodiversity, and economic development [22,23]. In China, A. philoxeroides has widely invaded more than 20 provinces and is included in the list of 59 invasive alien species under key management, and climate change threatens to further exacerbate the damage of A. philoxeroides to native plants at higher latitudinal regions [24]. Previous studies have found that A. philoxeroides can enhance its photosynthesis rate by increasing its nitrogen absorption rate and resource utilization, seriously inhibiting native plant growth in interspecific competition [12]. In environments with high heterogeneity, A. philoxeroides has a stronger clonal integration ability than native plants [25], and the high phenotypic plasticity increases this invader’s tolerance to heavy metal stress, thus enhancing its competitiveness against native plants [26]. A. philoxeroides can reduce soil enzyme activity and disrupt the balance of soil microbial communities through allelopathic effects, thus decreasing the abundance of microorganisms that are beneficial to native plants [27]. The performance of A. philoxeroides is also influenced by soil factors; e.g., soil nitrogen addition can increase the stem length and biomass accumulation of A. philoxeroides, and its invasion trend is positively correlated with nitrogen concentration [28,29]. The most abundant soil nutrients, such as phosphorus and potassium, can promote the secretion of more allelopathic substances in A. philoxeroides roots, causing more intense interspecies competition [27,30]. The height, biomass, and chlorophyll of A. philoxeroides all decrease with decreased soil pH [31]. Under high nutrient levels, an increase in the soil N:P ratio can significantly inhibit A. philoxeroides growth [32]. In addition, A. philoxeroides invasion causes an increase in soil total carbon, nitrogen, and water content and a decrease in soil N:P and C:P ratios [28,33,34].
However, the impacts of soil properties on species diversity and distribution in A. philoxeroides-invaded and non-invaded plant communities are currently unclear, although this is crucial for elucidating the ecological mechanisms of A. philoxeroides invasions during environmental changes. In this study, we investigated herb communities that are invaded by A. philoxeroides (Figure 1) and the native communities without A. philoxeroides at the geographical boundary between North and South China. We hypothesize that there are differences in the ecological effects of soil physicochemical properties on plant diversity and species distribution between these two types of communities. Specifically, we addressed the following questions: (1) What is the coupling relationship between species diversity and soil properties in invaded and native plant communities? (2) What are the limited soil factors that determine species distribution in these two types of communities?
2. Results
2.1. Plant Species Composition and Invasion Trend
In total, 114 plant species belonging to 49 families and 103 genera were recorded in 30 control plots; the families with the highest richness were Asteraceae (16 genera; 17 spp.), Poaceae (13 genera; 13 spp.), and Leguminosae (6 genera; 8 spp.). A total of 108 plant species belonging to 42 families and 96 genera were recorded in 30 invaded plots; the families with the highest richness were Poaceae (17 genera; 17 spp.), Asteraceae (15 genera; 17 spp.), and Leguminosae (8 genera; 8 spp.). The dominant species (total IV > 0.5) in the control plots were Oxalis articulata, Cynodon dactylon, Eleusine indica, Galium odoratum, Duchesnea indica, Dichondra micrantha, etc.; O. articulata, C. dactylon, and E. indica were the constructive species in the control communities (Table 1). The dominant species (total IV > 0.5) in invaded plots were A. philoxeroides, Cynodon dactylon, Galium odoratum, Oxalis articulata, Eleusine indica, etc., among which, the total IV of A. philoxeroides was much higher than that of other species, showing the dominance of this invader as the monodominant constructive species (Table 1). Compared with the control plots, A. philoxeroides invasion increased the difference between the total IV of dominant species in the invaded plots, and only a few species that had strong ecological adaptability, such as C. dactylon, G. odoratum, O. articulata, and E. indica, could continue maintaining the highest dominance in invaded communities (Table 1). Regression analysis showed that A. philoxeroides invasion coverage significantly decreased with an increase in soil AK (R2 = 0.135 and p = 0.046; Figure 2A) and the Patrick richness index (R2 = 0.322 and p < 0.001; Figure 2B).
2.2. RDA Ordination of Soil Properties and Species Diversity
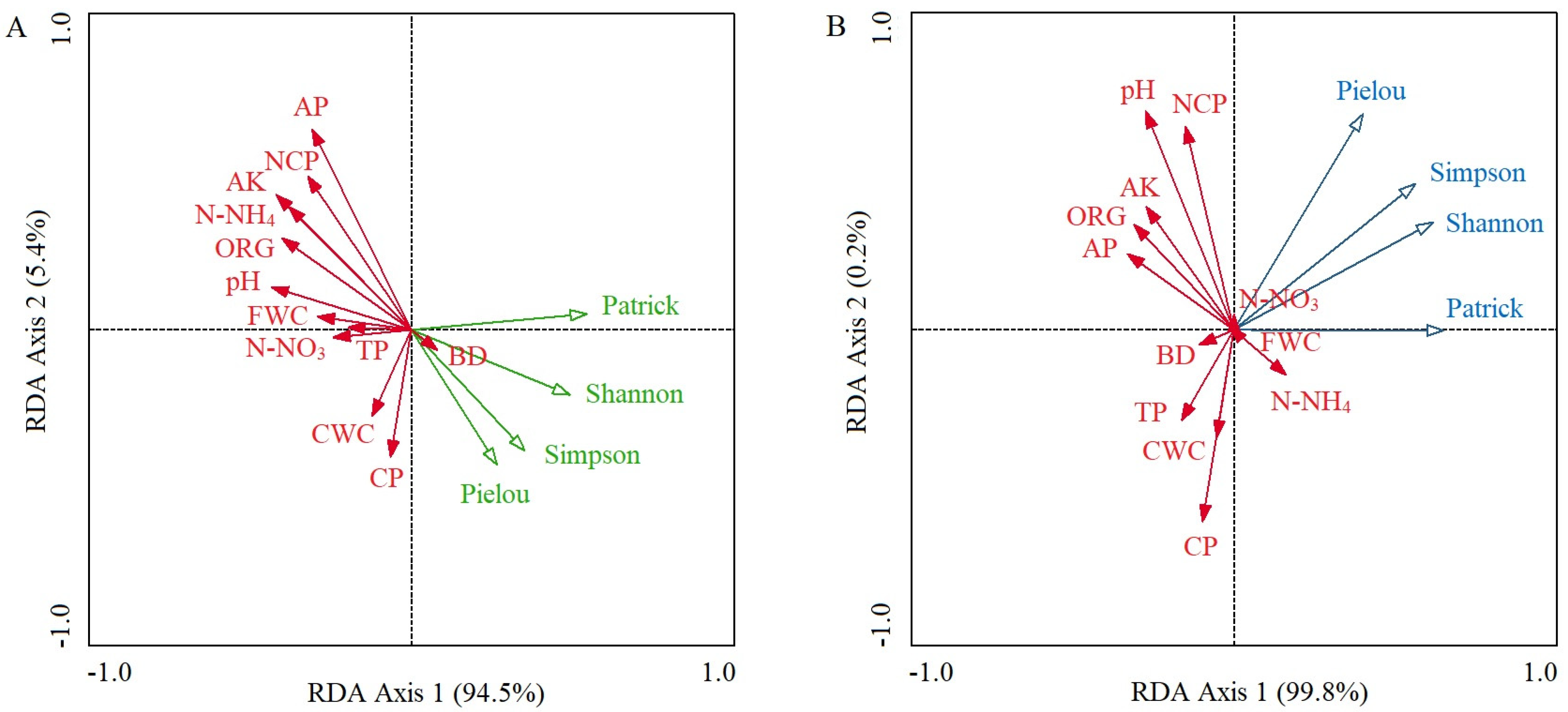
In the control plots, the cumulative percentage of variance explaining the soil properties–species diversity relationships of the first two RDA axes reached 99.9% (94.5% for axis 1 and 5.4% for axis 2), and the first axis had the decisive effect (Figure 3A). The dominant soil factors that determined RDA axis 1 were soil pH value (r = −0.433; p < 0.05), AK (r = −0.418; p < 0.05), ORG (r = −0.400; p < 0.05), and N-NH4 (r = −0.379; p < 0.05), and the values of these four soil factors gradually decreased from left to right along RDA axis 1 (Table 2; Figure 3A). The four α-species diversity indices all showed a strong negative relationship with soil pH, AK, ORG, and N-NH4 (Figure 3A). The determinant of RDA axis 2 was soil AP (r = 0.634; p < 0.01), and it had a strong negative relationship with the Simpson dominance index, the Pielou evenness index, and the Shannon–Wiener diversity index (Table 2; Figure 3A).
In the invaded plots, the accumulated percentage explanation of the first two RDA axes reached 100.0% (99.8% for axis 1 and 0.2% for axis 2); the first axis determined the relationship between soil properties and species diversity (Figure 3B). There was no significant correlation between any of the soil factors and RDA axis 1 (Table 2). The determinants of RDA axis 2 were soil pH value (r = 0.677; p < 0.01), NCP (r = 0.629; p < 0.01), and CP (r = −0.594; p < 0.01) (Table 2). Among these, the Pielou evenness index had a strong positive relationship with soil pH and NCP and a strong negative relationship with CP (Figure 3B).
2.3. Regression Analysis of Soil Properties and Species Diversity
In the control plots, the optimal fitting relationship between CWC and the Patrick richness index was a cubic equation (R2 = 0.262; p = 0.017); with increasing CWC, the Patrick richness index first increased but then decreased (Figure 4A). The variation in the Patrick richness index (R2 = 0.140; p = 0.041), the Simpson dominance index (R2 = 0.147; p = 0.036), and the Shannon–Wiener diversity index (R2 = 0.159; p = 0.029) along the soil N-NH4 gradient was characterized by inverse equations; these three diversity indices all decreased with increasing N-NH4 (Figure 4B–D). The optimal fitting relationships between soil AP and Simpson dominance index are represented by a compound equation (R2 = 0.136; p = 0.045); increasing soil AP decreased the Simpson index (Figure 4E). These were all consistent with the results of RDA ordination (Figure 3A).
In the invaded plots, NCP presented a significant logarithmic regression relationship with the Pielou evenness index (R2 = 0.158; p = 0.029); the Pielou index increased with an increase in NCP (Figure 4F). Increasing CP significantly decreased the Pielou evenness index, which is shown by a linear equation (R2 = 0.152; p = 0.033) (Figure 4G). These were all consistent with the results of RDA ordination (Figure 3B). However, in the control plots, the impact of CWC on species diversity was not consistent between RDA ordination and regression analysis, and this inconsistent impact also applies to the soil pH in the invaded plots.
2.4. CCA Ordination of Soil Properties and Species Distribution
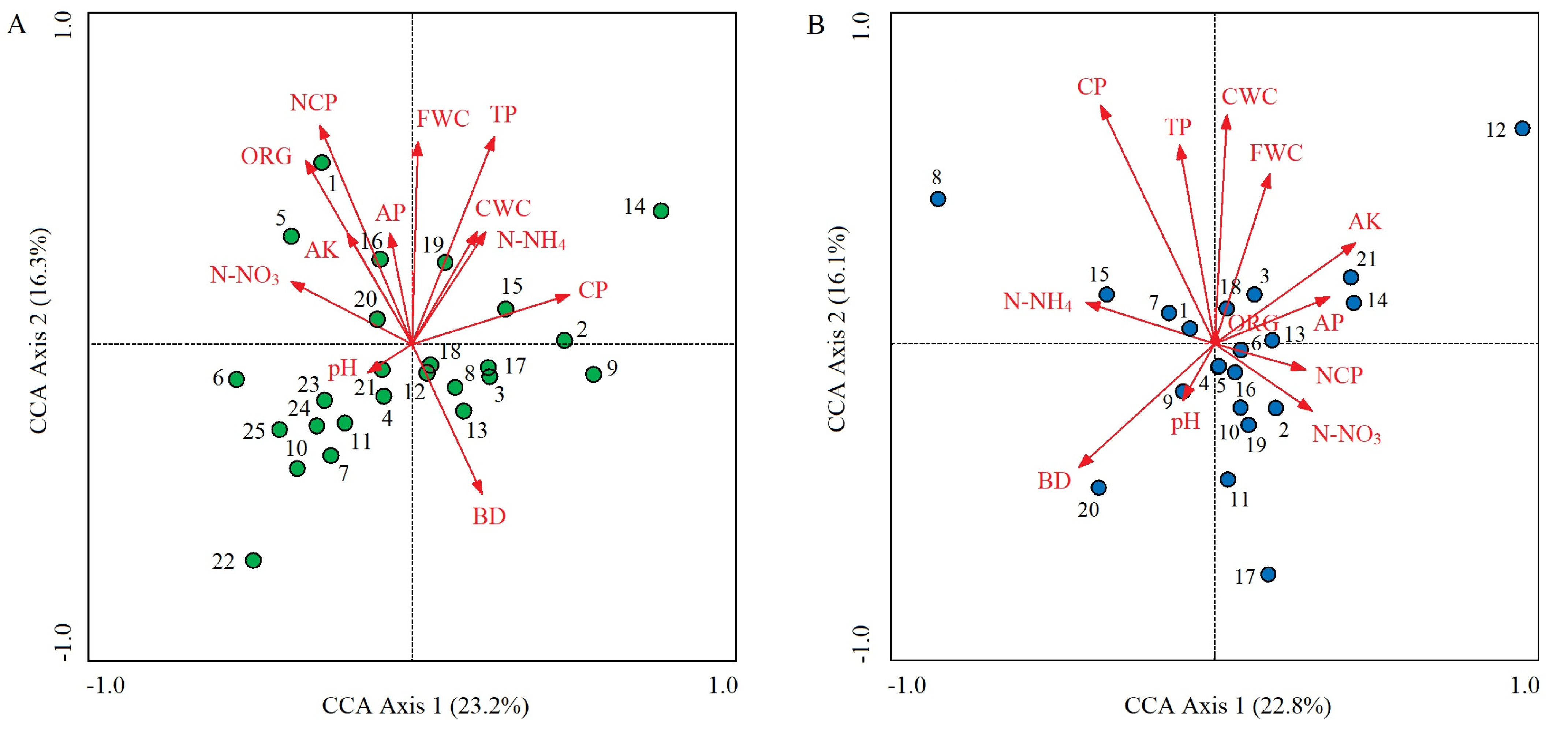
In the control plots, the cumulative percentage of variance explaining the soil properties–species distribution relationships of the first two CCA axes was 39.5% (23.2% for axis 1 and 16.3% for axis 2) (Figure 5A). The dominant soil factors that determined axis 1 were CP (r = 0.601; p < 0.01), N-NO3 (r = −0.463; p < 0.01), and ORG (r = −0.406’ p < 0.05); from left to right along axis 1, CP gradually increased, while soil N-NO3 and ORG gradually decreased (Table 3, Figure 5A). The determinants of axis 2 were NCP (r = 0.835; p < 0.01), TP (r = 0.791; p < 0.01), FWC (r = 0.772; p < 0.01), and ORG (r = 0.701; p < 0.01). These four soil factors all gradually increased from bottom to top along axis 2 (Table 3; Figure 5A). O. articulata (1) tended to be distributed in areas with high NCP and ORG; C. dactylon (2), Digitaria sanguinalis (9), and Imperata cylindrica (15) were distributed in areas with high CP but low N-NO3; E. indica (3) was distributed in areas with low ORG and N-NO3; Duchesnea indica (5) was distributed in areas with high N-NO3; and many species, including G. odoratum (4), tended to be distributed in areas with low FWC and TP (Figure 5A).
In the invaded plots, the first two CCA axes had an accumulated percentage explanation of 38.9% (22.8% for axis 1 and 16.1% for axis 2) (Figure 5B). The soil factors that determined axis 1 were soil AK (r = 0.435; p < 0.05), BD (r = −0.419; p < 0.05), and N-NH4 (r = −0.398; p < 0.05); from left to right along axis 1, AK gradually increased, while BD and N-NH4 gradually decreased (Table 3; Figure 5B). The determinants of axis 2 were CP (r = 0.737; p < 0.01), CWC (r = 0.706; p < 0.01), TP (r = 0.614; p < 0.01), and FWC (r = 0.525; p < 0.01). These four soil factors gradually increased from bottom to top along axis 2 (Table 3 and Figure 5B). Beckmannia syzigachne (20) tended to be distributed in areas with high BD but low AK and FWC; Cyperus rotundus (15) was distributed in areas with high N-NH4; Artemisia argyi (11) and Dichondra micrantha (17) were distributed in areas with low CWC, CP, and TP; and A. philoxeroides (1), O. articulata (4), E. indica (5), and Rumex acetosa (6) were all located at the center of the CCA ordination diagram, indicating that they all had wide ecological adaptability and superior growth advantages based on the same resource combination (Figure 5B). Paspalum paspaloides (8) and Hydrocotyle sibthorpioides (12) were far from the soil vectors, indicating that their distribution was weakly influenced by soil factors (Figure 5B). Compared with the control plots, the clustering degree of species distribution in the invaded plots increased (Figure 5).
3. Discussion
3.1. Mechanism of Impact of Soil on Community Structure and Plant Invasions
In natural communities, each species has the inherent ecological niche to achieve coexistence [35]. In our study, we found that O. articulata had the highest IV in control communities because, as an alien noninvasive species, it has strong stressful resistance and a strong allelopathic effect that inhibits the growth of native plants, which is beneficial to expanding the living space and achieving rapid growth [36,37]. The IV of A. philoxeroides was much higher than that of other accompanying species in the invaded communities, possibly due to its strong clonal reproductive ability and phenotypic plasticity, allowing this invader to plunder resources on a large scale, enhance competitiveness, and thus suppress native plants [22,24,38]. The IVs of some dominant species, such as O. articulata, C. dactylon, E. indica, and G. odoratum, decreased in the invaded communities compared with those in the control communities. Moreover, several native species with weaker tolerance, such as Axonopus compressus, and Clinopodium chinense, were squeezed out of the dominant layer, which indicates that A. philoxeroides invasion intensified interspecific competitions and accelerated the process of community homogenization [39].
The biotic resistance hypothesis proposes that native communities with high species richness are less susceptible to plant invasions because available niches are already filled, and invasive plants would experience higher competitive pressures [40]. Some studies also show that, due to the differentiation of species along different resource gradients, communities with high species diversity have higher structural complexity, which increases the resistance of native communities to plant invasions [35,41]. Similar to these findings, increasing species richness decreased A. philoxeroides coverage in this study. This is because habitats with abundant species usually have stronger interspecific interactions, and intensified competition would limit the establishment and spread of invasive plants [4,42,43]. Furthermore, high species richness can effectively resist plant invasion by increasing native plant biomass and increasing the difference in functional traits between invasive and native species [44,45].
Due to the heterogeneous responses of growth rate, allelopathic effect, and competitiveness between native and invasive plants to soil environments, soil properties also significantly affect the plant invasion progress [13,16,30]. Among them, soil potassium can promote protein synthesis and improve the water use efficiency of plants, and it can increase plant species diversity, which effectively resists plant invasions [46,47]. In our study, we also found a negative relationship between soil AK and A. philoxeroides coverage, possibly because AK may promote the growth of many accompanying plant species in invaded communities, and the competition of plants for potassium weakens the positive ‘invasive plant–soil’ feedback [48,49], thus leading to a decrease in A. philoxeroides invasion.
3.2. Mechanism of Impact of Soil on Plant Diversity
The Patrick richness index of the control plots had a maximum value at moderate CWC because CWC is closely related to the root distribution, moisture absorption, physiological metabolism, and electrolyte balance of plants; soil drought easily causes plant death due to water deficiency, while excessive soil moisture can weaken plant respiration and thus decrease plant richness [11,47]. In RDA, the effect of CWC on plant diversity was weak, perhaps because the interactions between multiple soil factors masked the determinant of CWC. Regression analysis and RDA all showed that the increasing N-NH4 significantly decreased the Patrick richness index, the Simpson dominance index, and the Shannon–Wiener diversity index because high N-NH4 caused toxicity in seed germination, seedling growth, and tillering and thus inhibited plant survival [43]. Excessive N-NH4 can also disrupt the balance of alkaline cations of plants, causing indirect toxicity by accelerating the replacement of soil Ca+ and Mg+ ions [50]. In addition, high N-NH4 enhances the nitrogen-scavenging activity of soil microorganisms and decreases the biomass and secretions of plant roots, which might decrease plant diversity [51,52]. The Simpson dominance index had a strong negative relationship with soil AP in the control plots because high AP decreases the activity of soil microorganisms and acid phosphatase, which decelerate nutrient cycling, weaken plant nutrient utilization efficiency, and cause a decrease in plant diversity [52,53]. A similar result was found in the plant communities of the Ebinur Lake watershed [54].
None of the soil factors in the invaded plots determined the RDA axis 1 due to plant invasions disturbing the inherent community structure and the feedback between native plants and soil, which weakens the ecological effect of the soil [55]. The water-holding and retention functions of soil are dependent on the distribution and connectivity of soil pores, among which, non-capillary pores provide channels for the infiltration and exchange of air, water, and nutrients close to plant roots, while capillary pores provide soil with water storage functions [15,47,56]. In the invaded plots in our study, the Pielou evenness index had a positive relationship with NCP and a negative relationship with CP. This is because the increasing NCP provided more underground niches for most plant roots, which alleviated the interspecific competition of A. philoxeroides-invaded communities and increased the Pielou evenness index [57]. Although increasing CP can decrease soil saturated hydraulic conductivity, invasive plants, as ‘opportunity species’, usually have strong phenotypic plasticity and ecological adaptability, which increases the invaders’ tolerance to changes in soil microhabitats compared with native accompanying plants and achieves greater competitiveness [16,48,58]. This intensifies the exclusion of A. philoxeroides against native plants and thus decreases species evenness. Moreover, compared with the control plots, the determinants of plant diversity in the invaded plots were mostly soil physical indicators, possibly because A. philoxeroides, as the constructive species, had strong adaptability to soil nutrient fluctuations, which weakened the impact of soil chemical indicators on the species diversity of the invaded communities [22,59].
Notably, our study area is located on the Qinling–Huaihe line, which is the geographical boundary between North and South China and also a climatic transition zone from a subtropical climate to a warm temperate climate. This zone has complicated variations in climate, terrain, and rainfall; these factors can significantly affect soil physicochemical properties [60], and all of them constitute a high degree of habitat heterogeneity that provides favorable conditions for the systematic development, niche differentiation, and species coexistence of plants in the study area, resulting in a high native plant diversity for resisting plant invasions [61,62]. However, the fluctuating temperature and precipitation in the climatic transition zone have caused lots of alien plants to respond more positively than native plants, which can enhance their ecological plasticity and interspecific interactions, thus accelerating the spread of many invasive plants, including A. philoxeroides, in the study area [63,64]. Therefore, under rapid global climate change, we should intensify plant diversity protection and invasion assessment in the North–South climatic transition zone of China.
3.3. Mechanism of Impact of Soil on Plant Species Distributions
Soil pore structure affects the distribution, adsorption, and release of nutrients within or between soil aggregates, while soil moisture conditions can regulate the absorption efficiency and leaching process of soil nutrients by plants [56,65]. Most plant species in the control plots were distributed in areas with low TP and FWC, which was related to their own properties and soil conditions. Consistent with our results, some studies have found that many native weeds prefer to distribute in low-porosity habitats, and some have extremely strong drought resistance, which leads to their shallower roots and lower water demand, making them accumulate in soil with lower water capacity [14,66,67]. Moreover, other related studies have found that a moderate increase in soil compaction can stimulate herbaceous plants to produce high-density fine roots in order to form channels, thus increasing soil ventilation and permeability and enabling plants to better adapt to low-FWC habitats [68,69]. I. cylindrical and C. dactylon, which belong to the Poaceae family, develop root systems that can increase the sand content of soil and indirectly enhance soil cohesion; however, their root growth consumes a large amount of water, so they tend to distribute in high-soil CP habitats with strong water storage capacities [14,70]. D. indica has a high nitrogen absorption rate, while N-NO3, as the main nitrogen source, can promote the lateral root growth of D. indica and participate in metabolic regulation [71,72]; thus, D. indica positively correlates with N-NO3. O. articulata has thick roots and mainly relies on bulb propagation, which prompts it to distribute in loose and porous soil environments with high ORG [36,37].
A. philoxeroides was located at the center of the CCA diagram, indicating that this invader has high adaptability to various soil factors, which is not only due to its strong phenotypic plasticity, reproductive ability, and stress resistance [24,38] but also its regulation by the ‘soil–plant’ feedback effect, which further promotes A. philoxeroides expansion [28,29]. C. rotundus positively correlates with N-NH4 due to the mycorrhizal fungi in its root systems; these mycorrhizal fungi can increase the absorption of soil N-NH4 and transfer it to the host plant [73,74]. P. paspaloides and H. sibthorpioides were weakly affected by soil factors due to their wet and shade-tolerant properties, as well as their vegetative and sexual reproduction, which decreased the limiting effects of soil environments on these two plant species [75]. For P. paspaloides especially, as the most dominant accompanying species of aquatic ecotype A. philoxeroides, its demand for water environments is higher than for soil environments [22]. O. articulata was distributed in a location very close to A. philoxeroides, indicating that the ecological niche of these two plant species is similar, and they thus might be intensely competitive. Our previous study also found that O. articulata has a strong biotic substitution effect on A. philoxeroides invasion, particularly in soils with high fertility; the physiological plasticity and photosynthetic efficiency of O. articulata were higher than those of A. philoxeroides [76].
However, plant invasions can also affect biotic community structures by altering soil physicochemical properties [59]. In this study, some plant species, such as A. argyi, D. micrantha, and B. syzigachne, tended to be distributed in habitats with low soil porosity and water capacity, possibly because A. philoxeroides invasion decreases the retention of soil nutrition and moisture and causes a decrease in soil permeability [77]. Furthermore, a small input from A. philoxeroides can increase soil nitrogen cycling in invaded habitats [12,48]. All of these phenomena enhanced the small-scale soil resource heterogeneity and finally exacerbated the patchy plant distribution in the invaded communities.
4. Materials and Methods
4.1. Field Survey
Our study was conducted in Xinyang City, Southern Henan Province, China. Xinyang City is located in the upper reaches of the Huai River on the Qinling Mountains–Huaihe River Line (Qinling–Huaihe Line), which is the geographical boundary between North and South China. Xinyang City is in a transitional zone from a subtropical climate to a warm temperate climate, and it has high plant species diversity within this region. Our study area has an average annual sunshine time of 2000 h, an average annual temperature of 15.2 °C, an average frost-free period of 225 d, and an average annual precipitation of 1105 mm, and the main soils are paddy soil and yellow-brown soil [78]. Due to global changes, the number of invasive plants in Xinyang City has significantly increased, among which, A. philoxeroides has widely invaded multiple ecosystems [64].
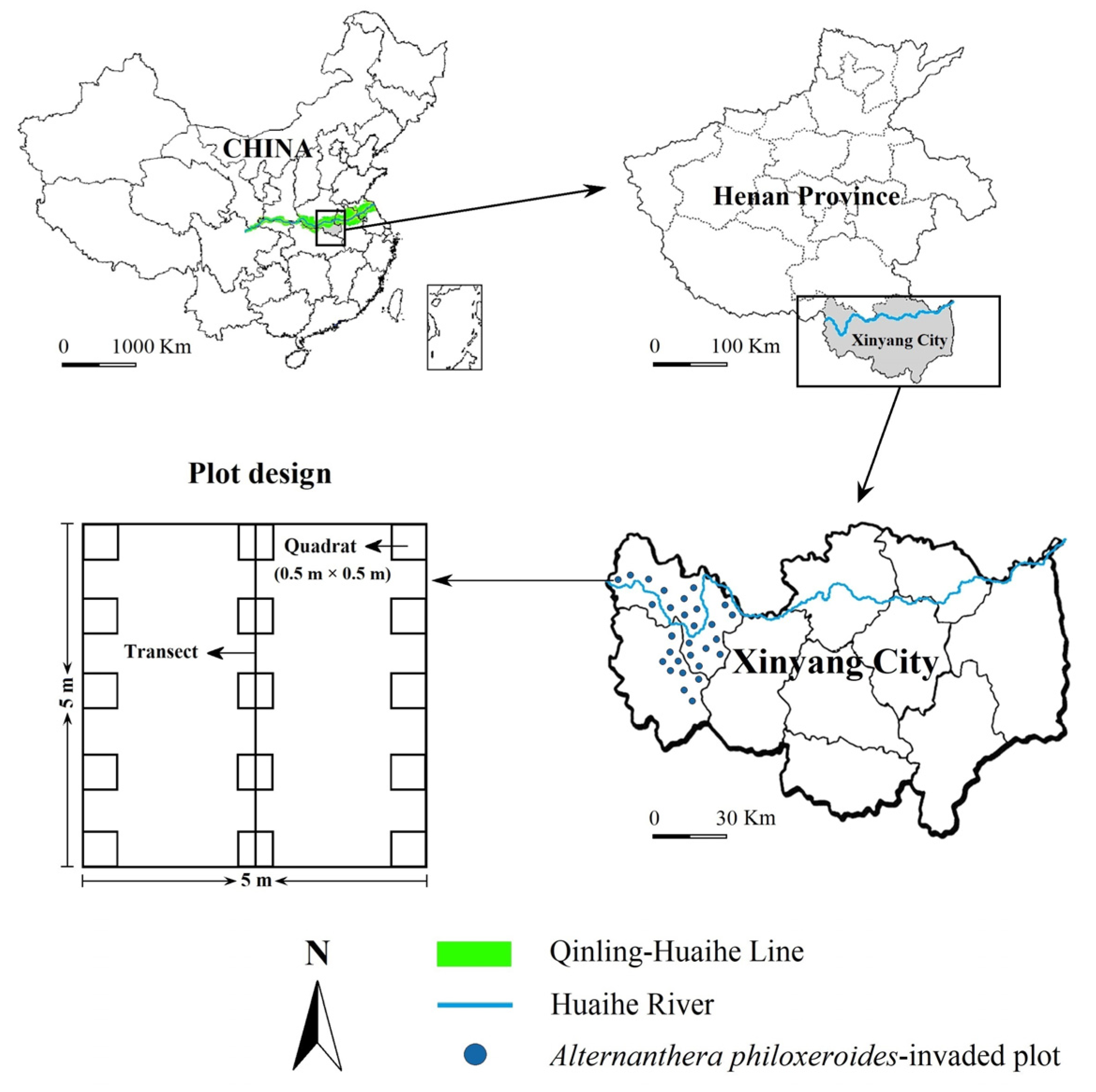
During the vigorous period of plant growth from June to July of 2022, we selected habitats with a continuous terrestrial A. philoxeroides invasion area of over 50 m2 to set invaded sampling plots in Xinyang City. In total, we set 30 A. philoxeroides-invaded plots, and every 2 plots was more than 5 km apart (Figure 6). We also set a total of 30 control plots (native plant communities without A. philoxeroides invasion) in areas with similar habitats near each invaded plot. Each plot had an area of 5 m × 5 m. We evenly set three 5 m transects in each plot and then evenly set five quadrats with an area of 0.5 m × 0.5 m along each transect for plant community investigation (Figure 6). We recorded the species name, abundance, individual height, and coverage of all plant species in each sampling plot. We recorded the number of asexual branches (for clonal plants), tillers (for Poaceae plants), or individuals (for non-clonal plants) as the abundance measurement for each plant species. We defined plant height as the vertical distance from the soil surface to the tip of the highest branch or longest leaf and measured the average height of 10 randomly selected individuals by using steel tape (if there were <10 individuals in a species, we measured all of its individuals). We used a 0.5 m × 0.5 m metal frame with 100 cells to calculate the coverage of plants [22].
After completing the plant community investigation, we evenly set three soil sampling sites along an arbitrary diagonal in each plot using ring knives, each with a volume of 100 cm3, to collect in situ soil from the 0–20 cm soil layer. We then brought these ring knives carrying in situ soil back to the laboratory to measure 6 soil physical indicators, namely, bulk density (BD), full water capacity (FWC), capillary water capacity (CWC), non-capillary porosity (NCP), capillary porosity (CP), and total porosity (TP). We also used the five-point sampling method to collect soil samples from a 0–20 cm soil layer, evenly mixed soil samples from the same plot into one part, and then put them into plastic bags separately. After drying and sieving, we measured 6 soil chemical indicators, namely, ammonium nitrogen (N-NH4), nitrate nitrogen (N-NO3), available phosphorus (AP), available potassium (AK), organic matter (ORG), and pH value.
4.2. Indicator Measurement
4.2.1. Measurement of Soil Physical Properties
We put each ring knife carrying in situ soil (without the top and bottom covers) in a flat-bottomed container, added water to the container until it was flush with the upper edge of the ring knife, and let the ring knife soak in water for several hours until the soil was saturated with water. We then removed the ring knife from the container, wiped off any external water, and covered the top and bottom covers; the total weight of the ring knife and soils in this first state was recorded as W1. We then opened the top and bottom covers (retaining the mesh) and placed the ring knife on the bracket for 12 h to facilitate the drainage of gravity water from the in situ soil and then covered the top and bottom covers again; the total weight in this second state was recorded as W2. We then opened the top and bottom covers and placed the in situ soil in a ring knife that carried it into a drying oven (Leirun 101-4A, China) set to 105 °C, and it remained there until it reached a constant weight; the total weight in this third state was recorded as W3. We also recorded the weight of an empty ring knife as W0 and the volume of a ring knife as V. Based on the above data, we could further calculate the 6 soil physical indicators [79].
4.2.2. Measurement of Soil Chemical Properties
The inorganic nitrogen of the soil samples was extracted at 2 mol/L KCl. We then measured soil N-NH4 and N-NO3 by using an auto discrete analyzer (Easychem Plus, Systea S.p.A., Rome, Italy). We measured soil AP and AK by using a continuous flow analyzer (Futura, AMS Alliance, Paris, France). We measured the total soil organic carbon by using a vario TOC cube analyzer (Elementar Analysensysteme GmbH, Hanau, Germany) and multiplied it by 1.724 to obtain the value of soil ORG. We mixed the soil samples with deionized water in a 1:5 ratio (weight–volume) and measured the soil pH value by using an intelligent acidity meter (Mettler-Toledo International Inc., Zurich, Switzerland) [22].
4.3. Data Calculation
4.3.1. Soil Physical Indicators
Based on the data measured with the ring knife and water immersion methods, as described in Section 4.2.1, the 6 soil physical indicators were calculated by using the following formulas [80]:
BD = (W3 − W0)/V;
FWC = (W1 − W3)/(W3 − W0) × 100%;
CWC = (W2 − W3)/(W3 − W0) × 100%;
NCP = (W1 − W2)/V × 100%;
CP = (W2 − W3)/V × 100%;
TP = (W1 − W3)/V × 100%.
4.3.2. Importance Value
The importance value (IV) is a comprehensive indicator that measures the status and role of a species in communities; the larger the IV, the higher the dominance of this species. The relative IV was calculated by using the following formula [38]:
where relative height, relative coverage, and relative abundance were the percentages of a species’ average height, coverage, and abundance over the sum of all species’ average height, coverage, and abundance within a plot, respectively.
Relative IV = (relative height + relative coverage + relative abundance)/3
The total IV of a species was the sum of its relative IV in all sampling plots.
4.3.3. Species Diversity
The following 4 α-species diversity indices were calculated to assess the species diversity of A. philoxeroides-invaded and native plant communities [22]:
where S is the total number of plant species in a plot, and Pi is the relative IV of species i.
Patrick richness index: R = S;
Simpson dominance index: λ = 1 − ΣPi2;
Shannon–Wiener diversity index: H = −ΣPi × lnPi;
Pielou evenness index: E = H/lnS.
4.4. Statistical Analyses
We conducted a regression analysis to examine the pairwise coupling relationships between soil physicochemical indicators, species diversity index, and A. philoxeroides invasion coverage by using the SPSS 16.0 software (SPSS Inc., Chicago, IL, USA). This software provided 11 curve-fitting models, and we selected the significant regression model with the highest fitting coefficient for analysis. We established an environmental matrix containing 12 soil physicochemical indicators (30 × 12) and a species diversity matrix containing 4 diversity indices (30 × 4). We then conducted redundancy analysis (RDA) to examine the comprehensive impacts of soil factors on plant diversity by using the Canoco 4.5 software (Microcomputer Power, New York, NY, USA). We also established a relative IV matrix containing 25 dominant plant species in native communities (30 × 25) and 21 dominant plant species in invaded communities (30 × 21), and we conducted canonical correspondence analysis (CCA) to examine the plant distribution along the soil environmental gradient with the Canoco 4.5 software. A Monte Carlo permutation test based on 499 random permutations was performed to test the significance of the correlation coefficient between soil indicators and RDA axes and CCA axes. We drew the RDA and CCA ordination diagrams by using the CanoDraw function of the Canoco 4.5 software [22].
5. Conclusions
We found that A. philoxeroides invasion widened the gap between the importance values of dominant plant species. Some native plants with weak tolerance were pushed out of the dominant layer by A. philoxeroides, and the coverage of A. philoxeroides decreased with increasing soil AK and species richness. Soil chemical properties determined the species diversity of control plant communities but had a weak impact on that of A. philoxeroides-invaded plant communities, while the species evenness of the invaded plant communities was mainly affected by soil porosity. The determinants of species distribution in the control communities were soil porosity, N-NO3, ORG, and FWC, while the determinants in invaded communities were soil AK, BD, N-NH4, porosity, and water capacity. These results indicate that A. philoxeroides invasion intensifies the interspecific competition between plant species in invaded habitats, and the plant species diversity and distribution in control and invaded communities have different responses to soil physicochemical properties. Thus, we should intensify the monitoring of changes in soil properties in A. philoxeroides-invaded habitats and implement biotic replacement strategies based on the heterogeneous responses of plants to soil resource fluctuations. Our findings could provide important implications for elucidating the species coexistence mechanisms of invaded ecosystems and predicting plant invasions under global environmental changes.
Author Contributions
Conceptualization, H.W. and B.R.; methodology, H.W.; formal analysis, H.W. and Y.L.; investigation, Y.L., T.Z. and M.X.; data curation, H.W. and Y.L.; writing—original draft preparation, H.W. and Y.L.; writing—review and editing, H.W., Y.L. and B.R.; funding acquisition, H.W. and B.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (31800460), the Xinyang Academy of Ecological Research Open Foundation (2023XYMS16), the Science and Technology Research Project of Henan Province (232102110062), the Key Scientific Research Projects of Higher Education Institutions of Henan Province (24A180028), the Nanhu Scholars Program for Young Scholars of Xinyang Normal University (XYNU) (2023A017), and the Dabie Mountain Laboratory Pre-Research Project.
Data Availability Statement
Data will be made available upon request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fernandez, R.D.; Castro-Diez, P.; Aragon, R.; Perez-Harguindeguy, N. Changes in community functional structure and ecosystem properties along an invasion gradient of Ligustrum lucidum. J. Veg. Sci. 2021, 32, e13098. [Google Scholar] [CrossRef]
- Fahey, C.; Angelini, C.; Flory, S.L. Grass invasion and drought interact to alter the diversity and structure of native plant communities. Ecology 2018, 99, 2692–2702. [Google Scholar] [CrossRef]
- Livingstone, S.W.; Isaac, M.E.; Cadotte, M.W. Invasive dominance and resident diversity: Unpacking the impact of plant invasion on biodiversity and ecosystem function. Ecol. Monogr. 2020, 90, e01425. [Google Scholar] [CrossRef]
- Carboni, M.; Livingstone, S.W.; Isaac, M.E.; Cadotte, M.W. Invasion drives plant diversity loss through competition and ecosystem modification. J. Ecol. 2021, 109, 3587–3601. [Google Scholar] [CrossRef]
- Wang, C.Y.; Wu, B.D.; Jiang, K.; Zhou, J.W.; Du, D.L. Canada goldenrod invasion affect taxonomic and functional diversity of plant communities in heterogeneous landscapes in urban ecosystems in East China. Urban For. Urban Green. 2019, 38, 145–156. [Google Scholar] [CrossRef]
- Balah, M.A.; Hassany, W.M. The relationship between Invasive Alien Solanum elaeagnifolium Cav. characters and impacts in different habitats. Biologia 2023, 78, 1253–1268. [Google Scholar] [CrossRef]
- Uddin, M.D.N.; Robinson, R.W. Responses of plant species diversity and soil physical-chemical-microbial properties to Phragmites australis invasion along a density gradient. Sci. Rep. 2017, 7, 11007. [Google Scholar] [CrossRef]
- Hansen, W.; Wollny, J.; Otte, A.; Eckstein, R.L.; Ludewig, K. Invasive legume affects species and functional composition of mountain meadow plant communities. Biol. Invasions 2021, 23, 281–296. [Google Scholar] [CrossRef]
- Ab Razak, N.; Gange, A.C.; Sutton, B.C.; Mansor, A. The invasive plant Impatiens glandulifera manipulates microbial associates of competing native species. Plants 2023, 12, 1552. [Google Scholar] [CrossRef]
- Süle, G.; Molnár, C.; Kovács-Hostyánszki, A.; Fenesi, A.; Bauer, N.; Szigeti, V. Escape from the garden: Spreading, effects and traits of a new risky invasive ornamental plant (Gaillardia aristata Pursh). NeoBiota 2023, 83, 43–69. [Google Scholar] [CrossRef]
- Yang, X.D.; Long, Y.X.; Sarkar, B.; Li, Y.; Lü, G.H.; Ali, A.; Yang, J.J.; Cao, Y.E. Influence of soil microorganisms and physicochemical properties on plant diversity in an arid desert of Western China. J. For. Res. 2021, 32, 2645–2659. [Google Scholar] [CrossRef]
- Liang, J.F.; Yuan, W.Y.; Gao, J.Q.; Roiloa, S.R.; Song, M.H.; Zhang, X.Y.; Yu, F.H. Soil resource heterogeneity competitively favors an invasive clonal plant over a native one. Oecologia 2020, 193, 155–165. [Google Scholar] [CrossRef]
- Gao, F.L.; He, Q.S.; Zhang, Y.D.; Yu, F.H.; Hou, J.H. Effects of soil nutrient heterogeneity on the growth and invasion success of alien plants: A multi-species study. Front. Ecol. Evol. 2021, 8, 619861. [Google Scholar] [CrossRef]
- Pan, J.; Bai, Z.K.; Cao, Y.G.; Zhou, W.; Wang, J.M. Influence of soil physical properties and vegetation coverage at different slope aspects in a reclaimed dump. Environ. Sci. Pollut. Res. 2017, 24, 23953–23965. [Google Scholar] [CrossRef]
- Tang, H.; Chen, M.; Wu, P.; Faheem, M.; Feng, Q.W.; Lee, X.Q.; Wang, S.S.; Wang, B. Engineered biochar effects on soil physicochemical properties and biota communities: A critical review. Chemosphere 2023, 311, 137025. [Google Scholar] [CrossRef]
- Li, W.T.; Bi, X.T.; Zheng, Y.L. Soil legacy effects on biomass allocation depend on native plant diversity in the invaded community. Sci. Prog. 2023, 106, 1–12. [Google Scholar] [CrossRef]
- Haury, J.; Druel, A.; Cabral, T.; Paulet, Y.; Bozec, M.; Coudreuse, J. Which adaptations of some invasive Ludwigia spp. (Rosidae, Onagraceae) populations occur in contrasting hydrological conditions in Western France? Hydrobiologia 2014, 737, 45–56. [Google Scholar] [CrossRef]
- Wang, C.Y.; Jiang, K.; Zhou, J.W.; Wu, B.D. Solidago canadensis invasion affects soil N-fixing bacterial communities in heterogeneous landscapes in urban ecosystems in East China. Sci. Total. Environ. 2018, 631–632, 702–713. [Google Scholar] [CrossRef]
- Yu, H.X.; Le Roux, J.J.; Jiang, Z.Y.; Sun, F.; Peng, C.L.; Li, W.H. Soil nitrogen dynamics and competition during plant invasion: Insights from Mikania micrantha invasions in China. New Phytol. 2021, 229, 3440–3452. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.X.; Lu, X.F.; Zhao, H.X.; Yang, Y.F.; Hale, L.; Gao, Q.; Liu, W.X.; Guo, J.Y.; Li, Q.; Zhou, J.Z.; et al. Ageratina adenophora invasions are associated with microbially mediated differences in biogeochemical cycles. Sci. Total. Environ. 2019, 677, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Hu, J.; Miao, L.; Yu, D.; Liu, C. The invasive stoloniferous clonal plant Alternanthera philoxeroides outperforms its co-occurring non-invasive functional counterparts in heterogeneous soil environments-invasion implications. Sci. Rep. 2016, 6, 38036. [Google Scholar] [CrossRef]
- Wu, H.; Carrillo, J.; Ding, J.Q. Species diversity and environmental determinants of aquatic and terrestrial communities invaded by Alternanthera philoxeroides. Sci. Total Environ. 2017, 581, 666–675. [Google Scholar] [CrossRef]
- Peng, X.M.; Yang, Y.F.; Yan, X.M.; Li, H.Y. The effects of water control on the survival and growth of Alternanthera philoxeroides in the vegetative reproduction and seedling stages. Sci. Rep. 2021, 11, 13556. [Google Scholar] [CrossRef]
- Wu, H.; Ismail, M.; Ding, J.Q. Global warming increases the interspecific competitiveness of the invasive plant alligator weed, Alternanthera philoxeroides. Sci. Total Environ. 2017, 575, 1415–1422. [Google Scholar] [CrossRef]
- You, W.H.; Li, N.N.; Zhang, J.; Song, A.; Du, D.L. The plant invader Alternanthera philoxeroides benefits from clonal integration more than its native co-genus in response to patch contrast. Plants 2023, 12, 2371. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, C.; Xiong, Y.T.; Wang, Y.; Li, Q.J. Combination effects of heavy metal and inter-specific competition on the invasiveness of Alternanthera philoxeroides. Environ. Exp. Bot. 2021, 189, 104532. [Google Scholar] [CrossRef]
- Ge, Y.Y.; Wang, Q.L.; Wang, L.; Liu, W.X.; Liu, X.Y.; Huang, Y.J.; Christie, P. Response of soil enzymes and microbial communities to root extracts of the alien Alternanthera philoxeroides. Arch. Agron. Soil Sci. 2018, 64, 708–717. [Google Scholar] [CrossRef]
- Cao, Q.; Zhang, H.; Ma, W.; Wang, R.; Liu, J. Composition characteristics of organic matter and bacterial communities under the Alternanthera philoxeroide invasion in wetlands. Appl. Sci. 2020, 10, 5571. [Google Scholar] [CrossRef]
- Harms, N.E.; Knight, I.A.; DeRossette, A.B.; Williams, D.A. Intraspecific trait plasticity to N and P of the wetland invader, Alternanthera philoxeroides under flooded conditions. Ecol. Evol. 2023, 13, e9966. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.Y.; Gao, H.; Li, J.; Wang, Y.H.; Gao, A.G.; Wen, J.H.; Balah, M.A.; Wu, A.P. The latitudinal and longitudinal allelopathic patterns of an invasive alligator weed (Alternanthera philoxeroides) in China. PLoS ONE 2023, 18, e0280866. [Google Scholar] [CrossRef] [PubMed]
- He, M.Y.; Hua, Z.X.; Chen, H.Y.; Liu, Y.; Li, Y.; Zhang, Z. Effects of simulated acid rain on rhizosphere microorganisms of invasive Alternanthera philoxeroides and native Alternanthera sessilis. Front. Microbiol. 2022, 13, 993147. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Chang, R.Y.; Guo, X.; Liang, X.Q.; Wang, R.Q.; Liu, J. Shifts in growth and competitive dominance of the invasive plant Alternanthera philoxeroides under different nitrogen and phosphorus supply. Environ. Exp. Bot. 2017, 135, 118–125. [Google Scholar] [CrossRef]
- Yang, R.R.; Li, K.; Fang, J.H.; Quan, Q.; Zhang, C.; Liu, J. The invasion of Alternanthera philoxeroides increased soil organic carbon in a river and a constructed wetland with different mechanisms. Front. Ecol. Evol. 2020, 8, 574528. [Google Scholar] [CrossRef]
- Yan, Z.W.; Lv, T.; Liu, Y.; Xing, B.; Chao, C.X.; Li, Y.; Wu, L.; Wang, L.G.; Liu, C.H.; Yu, D. Responses of soil phosphorus cycling and bioavailability to plant invasion in river-lake ecotones. Ecol. Appl. 2023, 33, e2843. [Google Scholar] [CrossRef] [PubMed]
- McIntire, E.J.B.; Fajardo, A. Facilitation as a ubiquitous driver of biodiversity. New Phytol. 2014, 201, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, S.; Watanabe, I.; Kuno, K.; Fujii, Y. Evaluation of the allelopathic activity of five Oxalidaceae cover plants and the demonstration of potent weed suppression by Oxalis species. Weed Biol. Manag. 2005, 5, 128–136. [Google Scholar] [CrossRef]
- Lorenzo, P.; González, L.; Ferrero, V. Effect of plant origin and phenological stage on the allelopathic activity of the invasive species Oxalis pescaprae. Am. J. Bot. 2021, 108, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Carrillo, J.; Ding, J.Q. Invasion by alligator weed, Alternanthera philoxeroides, is associated with decreased species diversity across the latitudinal gradient in China. J. Plant Ecol. 2016, 9, 311–319. [Google Scholar] [CrossRef]
- Chen, H.; Qian, H.; Spyreas, G.; Crossland, M. Native-exotic species richness relationships across spatial scales and biotic homogenization in wetland plant communities of Illinois, USA. Divers. Distrib. 2010, 16, 737–743. [Google Scholar] [CrossRef]
- Mungi, N.A.; Qureshi, Q.; Jhala, Y.V. Role of species richness and human impacts in resisting invasive species in tropical forests. J. Ecol. 2021, 109, 3308–3321. [Google Scholar] [CrossRef]
- Yan, Y.Z.; Jarvie, S.; Zhang, Q.; Han, P.; Liu, Q.F.; Zhang, S.S.; Liu, P.T. Habitat heterogeneity determines species richness on small habitat islands in a fragmented landscape. J. Biogeogr. 2023, 50, 976–986. [Google Scholar] [CrossRef]
- Chen, D.; Kleunen, M.V. Invasional meltdown mediated by plant-soil feedbacks may depend on community diversity. New Phytol. 2022, 235, 1589–1598. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, Q.; Yu, Q.G.; Ye, J.; Gao, J.W.; Liu, H.T.; Yong, J.W.H.; Yu, Y.J.; Liu, X.X.; Kong, H.M.; et al. Is the NH4+-induced growth inhibition caused by the NH4+ form of the nitrogen source or by soil acidification? Front. Plant Sci. 2022, 13, 968707. [Google Scholar] [CrossRef] [PubMed]
- Li, S.P.; Jia, P.; Fan, S.Y.; Wu, Y.T.; Liu, X.; Meng, Y.N.; Li, Y.; Shu, W.S.; Li, J.T.; Jiang, L. Functional traits explain the consistent resistance of biodiversity to plant invasion under nitrogen enrichment. Ecol. Lett. 2022, 25, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Qian, H.; Zhang, J. Does regional species diversity resist biotic invasions? Plant Divers. 2023, 45, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Tang, Y.; Tian, J.; Wu, X. Correlation between plant diversity and the physicochemical properties of soil microbes. Appl. Ecol. Environ. Res. 2019, 17, 10371–10388. [Google Scholar] [CrossRef]
- Di, W.U.; Andales, A.A.; Yang, H.; Sun, Q.; Chen, S.; Guo, X.; Li, D.H.; Du, T.S. Linking crop water productivity to soil physical, chemical and microbial properties. Front. Agric. Sci. Eng. 2021, 8, 545–558. [Google Scholar]
- Crawford, K.M.; Knight, T.M. Competition overwhelms the positive plant-soil feedback generated by an invasive plant. Oecologia 2017, 183, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Khatri, K.; Negi, B.; Bargali, K.; Bargali, S.S. Trait plasticity: A key attribute in the invasion success of Ageratina adenophora in different forest types of Kumaun Himalaya, India. Environ. Dev. Sustain. 2023. on line. [Google Scholar] [CrossRef]
- Xiao, C.B.; Fang, Y.; Wang, S.M.; He, K. The alleviation of ammonium toxicity in plants. J. Integr. Plant Biol. 2023, 65, 1362–1368. [Google Scholar] [CrossRef]
- Meier, I.C.; Finzi, A.C.; Phillips, R.P. Root exudates increase N availability by stimulating microbial turnover of fast-cycling N pools. Soil Biol. Biochem. 2017, 106, 119–128. [Google Scholar] [CrossRef]
- Luo, R.Y.; Fan, J.L.; Wang, W.J.; Luo, J.F.; Kuzyakov, Y.; He, J.S.; Chu, H.Y.; Ding, W.X. Nitrogen and phosphorus enrichment accelerates soil organic carbon loss in alpine grassland on the Qinghai-Tibetan Plateau. Sci. Total Environ. 2019, 650, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Qiu, T.Y.; Yu, J.L.; He, L.Y.; Liu, J.; Cui, Q.L.; Cui, Y.X.; Duan, C.J.; Zhao, S.L.; Wang, Y.Q.; Fang, L.C. Slope position mediates the co-utilization of phosphorus by plants and microbes through rhizosphere processes in a phosphorus-limited forest. Catena 2023, 222, 106808. [Google Scholar] [CrossRef]
- Ji, S.W.; Jiang, L.M.; Hu, D.; Lv, G.H. Impacts of plant and soil stoichiometry on species diversity in a desert ecosystem. AoB Plants 2022, 14, plac034. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.P.; Li, W.T.; Li, J.; Feng, Y.L. Temporal dynamics of plant-soil feedback and related mechanisms depend on environmental context during invasion processes of a subtropical invader. Plant Soil 2024, 496, 539–554. [Google Scholar] [CrossRef]
- Feng, X.; Xia, X.; Chen, S.T.; Lin, Q.M.; Zhang, X.H.; Cheng, K.; Liu, X.Y.; Bian, R.J.; Zheng, J.F.; Li, L.Q.; et al. Amendment of crop residue in different forms shifted micro-pore system structure and potential functionality of macroaggregates while changed their mass proportion and carbon storage of paddy topsoil. Geoderma 2022, 409, 115643. [Google Scholar] [CrossRef]
- De Long, J.R.; Heinen, R.; Heinze, J.; Morrien, E.; Png, G.K.; Sapsford, S.J.; Teste, F.P.; Fry, E.L. Plant-soil feedback: Incorporating untested influential drivers and reconciling terminology. Plant Soil. 2023, 485, 7–43. [Google Scholar] [CrossRef]
- Wu, J.; Yu, F.H. Belowground bud bank of invasive plants contributes to their successful invasion in coastal wetlands. Restor. Ecol. 2023, 31, e13821. [Google Scholar] [CrossRef]
- Lone, P.A.; Dar, J.A.; Subashree, K.; Raha, D.; Pandey, P.K.; Ray, T.; Khare, P.K.; Khan, M.L. Impact of plant invasion on physical, chemical and biological aspects of ecosystems: A review. Trop. Plant Res. 2019, 6, 528–544. [Google Scholar] [CrossRef]
- Kou, Z.; Yao, Y.; Hu, Y.; Zhang, B. Discussion on position of China’s north-south transitional zone by comparative analysis of mountain altitudinal belts. J. Mt. Sci. 2020, 17, 1901–1915. [Google Scholar] [CrossRef]
- Maron, J.; Marler, M. Native plant diversity resists invasion at both low and high resource levels. Ecology 2007, 88, 2651–2661. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Jia, X.; Han, L.; Tian, G.; Kang, S.; Zhao, Y. Spatial characteristics of the dominant fungi and their driving factors in forest soils in the Qinling Mountains, China. Catena 2021, 206, 105504. [Google Scholar] [CrossRef]
- Wallingford, P.D.; Morelli, T.L.; Allen, J.M.; Beaury, E.M.; Blumenthal, D.M.; Bradley, B.A.; Dukes, J.S.; Early, R.; Fusco, E.J.; Goldberg, D.E.; et al. Adjusting the lens of invasion biology to focus on the impacts of climate-driven range shifts. Nat. Clim. Change 2020, 10, 398–405. [Google Scholar] [CrossRef]
- Yan, H.Y.; Feng, L.; Zhao, Y.F.; Feng, L.; Wu, D.; Zhu, C.P. Prediction of the spatial distribution of Alternanthera philoxeroides in China based on ArcGIS and MaxEnt. Glob. Ecol. Conserv. 2020, 21, e00856. [Google Scholar] [CrossRef]
- Gerlach, R.; Buhk, C.; Schirmel, J.; Jungkunst, H.F. Exploration of the response of soil properties and plant species composition to centuries-old traditional meadow irrigation. J. Plant Ecol. 2023, 16, 72. [Google Scholar] [CrossRef]
- Yu, B.Q.; Xie, C.K.; Cai, S.Z.; Chen, Y.; Lv, Y.P.; Mo, Z.L.; Liu, T.L.; Yang, Z.W. Effects of tree root density on soil total porosity and non-capillary porosity using a ground-penetrating tree radar unit in Shanghai, China. Sustainability 2018, 10, 4640. [Google Scholar] [CrossRef]
- Wu, W.J.; Chen, G.J.; Meng, T.F.; Li, C.; Feng, H.; Si, B.C.; Siddique, K.H.M. Effect of different vegetation restoration on soil properties in the semi-arid Loess Plateau of China. Catena 2023, 220, 106630. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Niu, J.Z.; Yu, X.X.; Zhu, W.L.; Du, X.Q. Effects of fine root length density and root biomass on soil preferential flow in forest ecosystems. For. Ecosyst. 2015, 24, e012. [Google Scholar] [CrossRef]
- Lyu, Q.; Liu, J.L.; Liu, J.J.; Luo, Y.; Chen, L.M.; Chen, G.; Zhao, K.J.; Chen, Y.Q.; Fan, C.; Li, X.W. Response of plant diversity and soil physicochemical properties to different gap sizes in a Pinus massoniana plantation. PeerJ 2021, 9, e12222. [Google Scholar] [CrossRef]
- Ma, Q.; Li, Z.; Xiao, H.L.; Hu, Z.; Pung, L. Mechanical properties of clay reinforced with Bermuda grass root under drying-wetting cycles. Environ. Earth Sci. 2021, 80, 31. [Google Scholar] [CrossRef]
- Blagodatskaya, E.; Littschwager, J.; Lauerer, M.; Kuzyakov, Y. Plant traits regulating N capture define microbial competition in the rhizosphere. Eur. J. Soil Biol. 2014, 61, 41–48. [Google Scholar] [CrossRef]
- Shannon-Firestone, S.; Reynolds, H.L.; Phillips, R.P.; Flory, S.L.; Yannarell, A. The role of ammonium oxidizing communities in mediating effects of an invasive plant on soil nitrification. Soil Biol. Biochem. 2015, 90, 266–274. [Google Scholar] [CrossRef]
- Aiswarya, N.K.; Muthuraj, K.; Siva, P.K.T.; Krishnan, E.; Nagarajan, N. Studies on the arbuscular mycorrhizal fungal diversity of selected medicinal plant species from kodikuthimala, malappuram, kerala. Kongunadu Res. J. 2017, 4, 51–58. [Google Scholar]
- Daryanto, S.; Wang, L.; Gilhooly, W.P.; Jacinthe, P.A. Nitrogen preference across generations under changing ammonium nitrate ratios. J. Plant Ecol. 2019, 12, 235–244. [Google Scholar] [CrossRef]
- Yang, J.; Li, E.H.; Zhou, R.; Xia, Y.; Yang, C.; Zhang, Y.Y. The effects of edaphic factors on riparian plants in the middle and lower reaches of the Hanjiang River, China. Plants 2022, 11, 531. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Dong, S.J.; Wang, L.; Zhu, Y.Y.; Jia, S.Q.; Rao, B.Q. Nitrogen enrichment alters the resistance of a noninvasive alien plant species to Alternanthera philoxeroides invasion. Front. Ecol. Evol. 2023, 11, 1215191. [Google Scholar] [CrossRef]
- Xie, H.T.; Tang, Y.; Yu, M.K.; Wang, G.G. The effects of afforestation tree species mixing on soil organic carbon stock, nutrients accumulation, and understory vegetation diversity on reclaimed coastal lands in Eastern China. Glob. Ecol. Conserv. 2021, 26, e01478. [Google Scholar] [CrossRef]
- Lei, Y.; Liao, R.W.; Su, Y.M.; Zhang, X.; Liu, D.Y.; Zhang, L. Variation characteristics of temperature and rainfall and their relationship with geographical factors in the Qinling Mountains. Atmosphere 2023, 14, 696. [Google Scholar] [CrossRef]
- Shi, S.L.; Zhao, F.Y.; Ren, X.M.; Meng, Z.J.; Dang, X.H.; Wu, X.L. Soil Infiltration properties are affected by typical plant communities in a semi-arid desert grassland in China. Water 2022, 14, 3301. [Google Scholar] [CrossRef]
- Bi, Y.L.; Zou, H.; Zhu, C.W. Dynamic monitoring of soil bulk density and infiltration rate during coal mining in sandy land with different vegetation. Int. J. Coal Sci. Technol. 2014, 1, 198–206. [Google Scholar] [CrossRef]
Figure 1.
Terrestrial A. philoxeroides-invaded plant communities (A) and the studied species, A. philoxeroides (B), in Xinyang City, China (picture: Hao Wu).
Figure 1.
Terrestrial A. philoxeroides-invaded plant communities (A) and the studied species, A. philoxeroides (B), in Xinyang City, China (picture: Hao Wu).

Figure 2.
Regression analysis between A. philoxeroides invasion coverage and soil factors (A) and species diversity index (B).
Figure 2.
Regression analysis between A. philoxeroides invasion coverage and soil factors (A) and species diversity index (B).
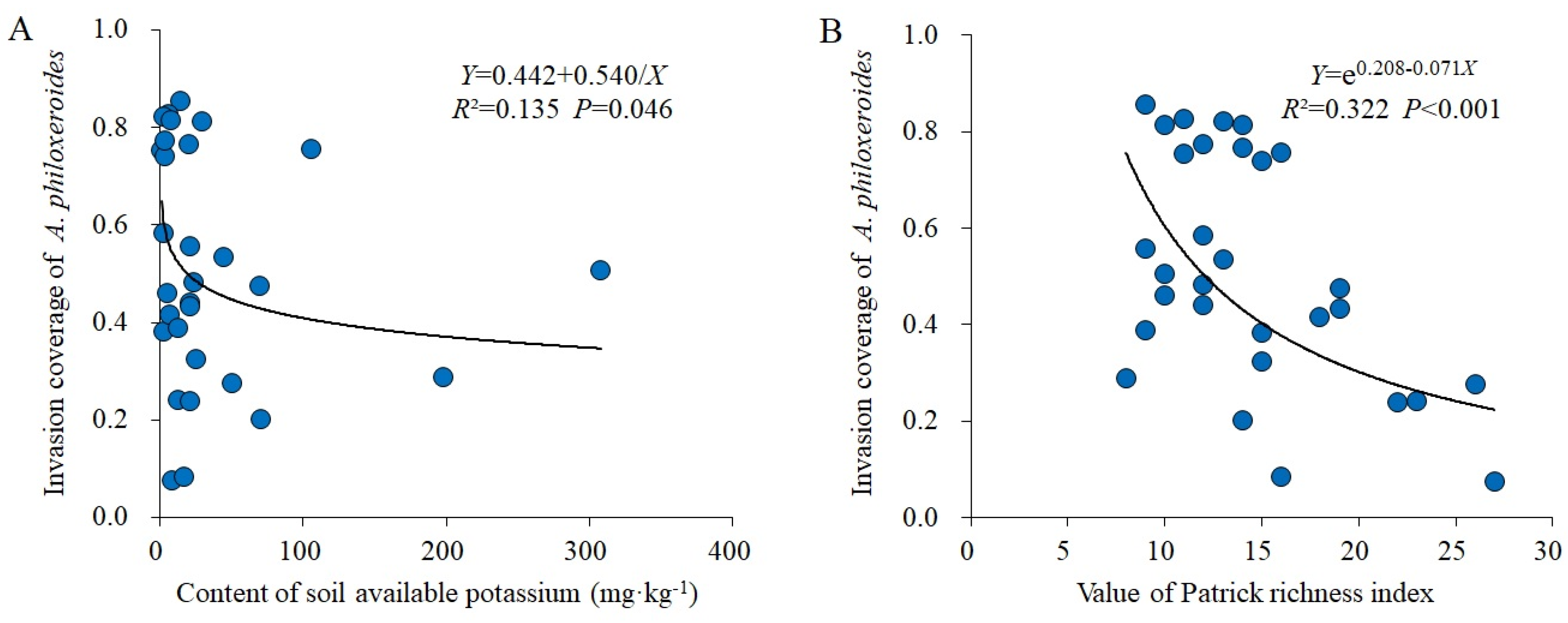
Figure 3.
RDA ordination diagram of soil factors and species diversity index in control (A) and A. philoxeroides-invaded plant communities (B). Patrick, Simpson, Shannon, and Pielou represent the Patrick richness index, Simpson dominance index, Shannon–Wiener diversity index, and Pielou evenness index. Solid red vectors represent the 12 soil factors; hollow vectors represent 4 α-species diversity indices. Arrows indicate the direction of increase in variables from the RDA ordination center. The angle between each pair of variables represents their correlations; the smaller the angle, the greater the correlation.
Figure 3.
RDA ordination diagram of soil factors and species diversity index in control (A) and A. philoxeroides-invaded plant communities (B). Patrick, Simpson, Shannon, and Pielou represent the Patrick richness index, Simpson dominance index, Shannon–Wiener diversity index, and Pielou evenness index. Solid red vectors represent the 12 soil factors; hollow vectors represent 4 α-species diversity indices. Arrows indicate the direction of increase in variables from the RDA ordination center. The angle between each pair of variables represents their correlations; the smaller the angle, the greater the correlation.

Figure 4.
Regression analysis between soil factors and species diversity index in control (A–E) and A. philoxeroides-invaded plant communities (F,G).
Figure 4.
Regression analysis between soil factors and species diversity index in control (A–E) and A. philoxeroides-invaded plant communities (F,G).

Figure 5.
CCA ordination diagrams of dominant plant species in control (A) and A. philoxeroides-invaded plant communities (B) along soil property gradient. Red vectors represent the 12 soil factors. Solid dots represent the dominant plant species, and the locations represent their optimal distribution positions under multiple soil factor combinations. Arabic numbers are codes of dominant plant species, and their Latin names are shown in Table 1. Vertical distance between dots and vectors represents the influence degree of a certain soil factor on a certain species distribution; the shorter the vertical distance, the greater the influence degree.
Figure 5.
CCA ordination diagrams of dominant plant species in control (A) and A. philoxeroides-invaded plant communities (B) along soil property gradient. Red vectors represent the 12 soil factors. Solid dots represent the dominant plant species, and the locations represent their optimal distribution positions under multiple soil factor combinations. Arabic numbers are codes of dominant plant species, and their Latin names are shown in Table 1. Vertical distance between dots and vectors represents the influence degree of a certain soil factor on a certain species distribution; the shorter the vertical distance, the greater the influence degree.
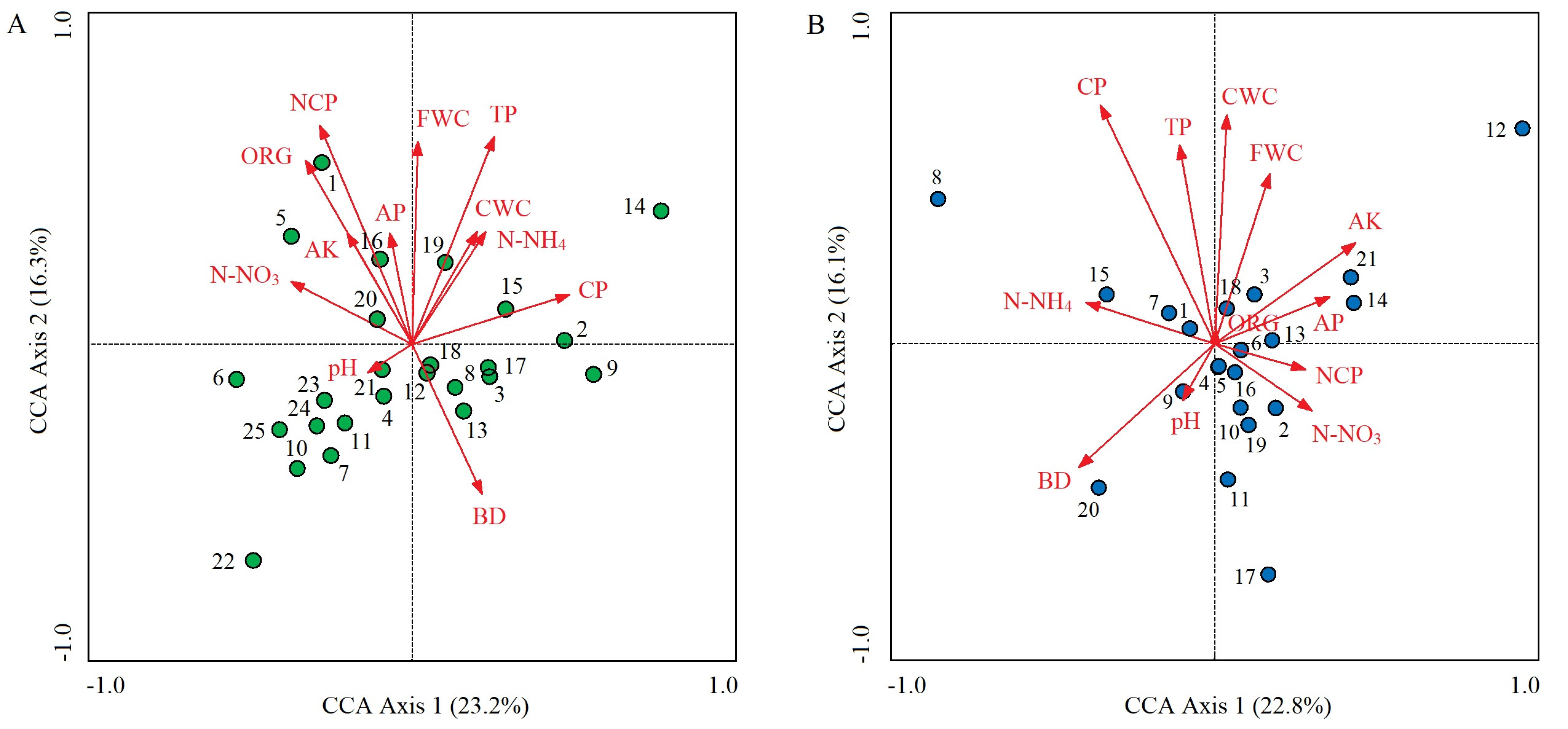
Figure 6.
Sampling plots of terrestrial A. philoxeroides-invaded plant communities along the geographical boundary between North and South China. Plot size: 5 × 5 m; transect size: 5 m; quadrat size: 0.5 × 0.5 m.
Figure 6.
Sampling plots of terrestrial A. philoxeroides-invaded plant communities along the geographical boundary between North and South China. Plot size: 5 × 5 m; transect size: 5 m; quadrat size: 0.5 × 0.5 m.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Total importance value (IV) of dominant species in control and A. philoxeroides-invaded plant communities.
Table 1.
Total importance value (IV) of dominant species in control and A. philoxeroides-invaded plant communities.
| Code | Plant Species | Total IV | Code | Plant Species | Total IV |
|---|---|---|---|---|---|
| Control communities | Invaded communities | ||||
| 1 | Oxalis articulata | 4.030 | 1 | Alternanthera philoxeroides | 15.309 |
| 2 | Cynodon dactylon | 3.817 | 2 | Cynodon dactylon | 2.266 |
| 3 | Eleusine indica | 3.533 | 3 | Galium odoratum | 1.917 |
| 4 | Galium odoratum | 2.862 | 4 | Oxalis articulata | 1.915 |
| 5 | Duchesnea indica | 1.645 | 5 | Eleusine indica | 1.758 |
| 6 | Dichondra micrantha | 1.614 | 6 | Rumex acetosa | 1.243 |
| 7 | Axonopus compressus | 1.376 | 7 | Medicago sativa | 1.116 |
| 8 | Lolium perenne | 1.318 | 8 | Paspalum paspaloides | 1.078 |
| 9 | Digitaria sanguinalis | 1.234 | 9 | Erigeron annuus | 0.993 |
| 10 | Medicago sativa | 1.119 | 10 | Duchesnea indica | 0.978 |
| 11 | Poa annua | 1.057 | 11 | Artemisia argyi | 0.956 |
| 12 | Erigeron annuus | 1.029 | 12 | Hydrocotyle sibthorpioides | 0.824 |
| 13 | Vicia hirsuta | 0.823 | 13 | Poa annua | 0.822 |
| 14 | Kyllinga brevifolia | 0.765 | 14 | Ophiopogon bodinieri | 0.822 |
| 15 | Imperata cylindrica | 0.760 | 15 | Cyperus rotundus | 0.710 |
| 16 | Ophiopogon bodinieri | 0.732 | 16 | Lolium perenne | 0.655 |
| 17 | Cayratia japonica | 0.721 | 17 | Dichondra micrantha | 0.646 |
| 18 | Mazus japonicus | 0.711 | 18 | Cayratia japonica | 0.622 |
| 19 | Stellaria media | 0.700 | 19 | Imperata cylindrica | 0.543 |
| 20 | Veronica polita | 0.648 | 20 | Beckmannia syzigachne | 0.532 |
| 21 | Sedum sarmentosum | 0.616 | 21 | Mazus japonicus | 0.512 |
| 22 | Kummerowia striata | 0.529 | |||
| 23 | Rumex acetosa | 0.526 | |||
| 24 | Plantago asiatica | 0.518 | |||
| 25 | Clinopodium chinense | 0.512 | |||
Table 2.
Correlations between the soil physicochemical factors and the first two RDA axes.
| Soil Factors | Control Communities | Invaded Communities | ||
|---|---|---|---|---|
| Axis 1 | Axis 2 | Axis 1 | Axis 2 | |
| BD | 0.081 | −0.064 | −0.107 | −0.047 |
| FWC | −0.290 | 0.041 | 0.009 | 0.011 |
| CWC | −0.121 | −0.273 | −0.054 | −0.333 |
| NCP | −0.321 | 0.404 * | −0.150 | 0.629 ** |
| CP | −0.064 | −0.402 * | −0.097 | −0.594 ** |
| TP | −0.196 | 0.007 | −0.162 | −0.280 |
| N-NH4 | −0.379 * | 0.388 * | 0.162 | −0.140 |
| N-NO3 | −0.241 | −0.025 | 0.001 | 0.047 |
| AP | −0.307 | 0.634 ** | −0.329 | 0.235 |
| AK | −0.418 * | 0.426 * | −0.271 | 0.380 * |
| ORG | −0.400 * | 0.289 | −0.309 | 0.325 |
| pH | −0.433 * | 0.135 | −0.273 | 0.677 ** |
BD, FWC, CWC, NCP, CP, TP, N-NH4, N-NO3, AP, AK, ORG, and pH represent bulk density, full water capacity, capillary water capacity, non-capillary porosity, capillary porosity, total porosity, ammonium nitrogen, nitrate nitrogen, available phosphorus, available potassium, organic matter, and pH value, respectively. * p < 0.05 level; ** p < 0.01 level. The same applies below.
Table 3.
Correlations between the soil physicochemical factors and the first two CCA axes.
| Soil Factors | Control Communities | Invaded Communities | ||
|---|---|---|---|---|
| Axis 1 | Axis 2 | Axis 1 | Axis 2 | |
| BD | 0.267 | −0.573 ** | −0.419 * | −0.383 * |
| FWC | 0.022 | 0.772 ** | 0.171 | 0.525 ** |
| CWC | 0.248 | 0.429 * | 0.038 | 0.706 ** |
| NCP | −0.352 | 0.835 ** | 0.280 | −0.080 |
| CP | 0.601 ** | 0.189 | −0.352 | 0.737 ** |
| TP | 0.314 | 0.791 ** | −0.109 | 0.614 ** |
| N-NH4 | 0.281 | 0.428 * | −0.398 * | 0.127 |
| N-NO3 | −0.463 ** | 0.238 | 0.300 | −0.207 |
| AP | −0.084 | 0.422 * | 0.355 | 0.145 |
| AK | −0.247 | 0.423 * | 0.435 * | 0.311 |
| ORG | −0.406 * | 0.701 ** | 0.004 | 0.046 |
| pH | −0.168 | −0.108 | −0.098 | −0.172 |
* p < 0.05 level; ** p < 0.01 level.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wu, H.; Liu, Y.; Zhang, T.; Xu, M.; Rao, B. Impacts of Soil Properties on Species Diversity and Structure in Alternanthera philoxeroides-Invaded and Native Plant Communities. Plants 2024, 13, 1196. https://doi.org/10.3390/plants13091196
AMA Style
Wu H, Liu Y, Zhang T, Xu M, Rao B. Impacts of Soil Properties on Species Diversity and Structure in Alternanthera philoxeroides-Invaded and Native Plant Communities. Plants. 2024; 13(9):1196. https://doi.org/10.3390/plants13091196
Chicago/Turabian StyleWu, Hao, Yuxin Liu, Tiantian Zhang, Mingxia Xu, and Benqiang Rao. 2024. "Impacts of Soil Properties on Species Diversity and Structure in Alternanthera philoxeroides-Invaded and Native Plant Communities" Plants 13, no. 9: 1196. https://doi.org/10.3390/plants13091196
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.